

Ботанический журнал, 2021, T. 106, № 2, стр. 107-125
РЯДЫ ВОССТАНОВЛЕНИЯ ЕЛЬНИКОВ ЧЕРНИЧНЫХ ВОСТОЧНОЙ ФЕННОСКАНДИИ
А. М. Крышень 1, *, Н. В. Геникова 1, Ю. В. Преснухин 1
1 Институт леса Карельского научного центра РАН
185910 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: kryshen@krc.karelia.ru
Поступила в редакцию 23.04.2020
После доработки 11.10.2020
Принята к публикации 14.10.2020
Аннотация
Представлена эколого-динамическая модель восстановления ельников черничных, отражающая современное состояние лесов. В пределах возрастной стадии ассоциации выстраиваются по условиям увлажнения и именуются по индикаторным видам. На стадии вырубки в условиях ельников черничных (Picea abies–Vaccinium myrtillus) выделено 4 ассоциации (P.a.–V.m.: Vaccinium vitis-idaea, P.a.–V.m.: Avenella flexuosa, P.a.–V.m.: Calamagrostis arundinacea, P.a.–V.m.: Carex canescens), на стадии молодняков (10–40 (60) лет) – 3 ассоциации (P.a.–V.m.: Picea abies + Betula spp. + Pinus sylvestris–Vaccinium vitis-idaea, P.a.–V.m.: Betula spp. + Populus tremula – Avenella flexuisa, P.a.–V.m.: Betula spp. + Populus tremula–Calamagrostis arundinacea + Geranium sylvaticum), возрастом от 60 до 100 (120) лет – 3 (P.a.–V.m.: Picea abies + Betula spp.+ Pinus sylvestris–Vaccinium myrtillus–Pleurozium schreberi, P.a.–V.m.: Picea abies + Populus tremula + Betula spp.–Vaccinium myrtillus + Ca-lamagrostis arundinacea + Hylocomium splendens, P.a.–V.m.: Betula spp. + Picea abies–Calamagrostis arundinaceae + Gymnocarpium dryopteris). В возрасте 100–120 лет происходит снижение участия лиственных пород в структуре древостоя, доминирование ели выравнивает условия и структуру напочвенного покрова – динамические ряды сходятся. С этого момента до климакса выделяется только одна ассоциация P.a.–V.m.: Picea abies–Vaccinium myrtillus–Pleurozium schreberi. Временные состояния или локальные проявления, отличающиеся структурой и не отражающие естественную динамику, а связанные с хозяйственной деятельностью или разрушением в результате природных явлений, относятся к рангу субассоциаций. Типология учитывает современное состояние лесов, естественную и антропогенную динамику и может служить основой для планирования лесохозяйственных мероприятий.
Статья продолжает серию публикаций, посвященных закономерностям формирования лесных растительных сообществ на автоморфных почвах и представляющих эколого-динамический подход к типологии лесов.
Среди причин катастрофического разрушения лесных сообществ в европейской тайге в настоящее время абсолютно преобладает рубка древостоя, масштабы вмешательства человека хорошо заметны из космоса (Lukina et al., 2015) – происходит активное изменение породной и возрастной структуры таежных лесов. Леса на автоморфных почвах, являющиеся основой лесоэксплуатационного фонда, в настоящее время представляют мозаику от вырубки до коренных сообществ. Их разнообразие определяется экотопом, сукцессионной стадией и разнообразным влиянием природных и антропогенных факторов. Систематизация этого ценотического разнообразия должна опираться на фундаментальные знания формирования структуры лесных растительных сообществ. Все эти моменты мы ранее подробно разбирали в публикациях в Ботаническом журнале: принципы выделения типов лесорастительных условий и возрастных стадий (Kryshen, 2010), эколого-динамическую типологию сосняков (Kryshen et al., 2018); анализ ценофлоры сосняков (Genikova et al., 2012) и ельников черничных (Genikova et al., 2019). Очень коротко повторим основные моменты, необходимые для понимания динамики ельников черничных и принципов классифицирования сообществ. Мы исходим из того, что каждому типу лесорастительных условий соответствует только один тип климаксового сообщества, характеристики которого достаточно четко определяются геоморфологическими особенностями и характером увлажнения (уровнем залегания грунтовых вод), а также условиями трофности. Всего для покрытых лесом местообитаний Восточной Фенноскандии нами выделено 5 типов лесорастительных условий с автоморфными почвами (подробно см. Kryshen, 2010): песчаные сухие олиготофные почвы с климаксом лишайниковые сосняки – Pinus sylvestris–[Cladonia] (P.s.–Cl.); песчаные сухие мезо-олиготрофные почвы – Pinus sylvestris–Vaccinium vitis-idaea (P.s.–V.v.-i.); песчаные свежие мезо-олиготрофные почвы – Pinus sylvestris–Vaccinium myrtillus (P.s.–V.m.); супесчаные свежие мезотрофные почвы – Picea abies–Vaccinium myrtillus (P.a.–V.m.); супесчаные и суглинистые свежие мезо-эвтрофные почвы – Picea abies–Oxalis acetosella (P.a.–O.a.).
Условия ельника черничного являются, пожалуй, самыми распространенными в европейской части таежной зоны России. Климаксным сообществом здесь является ельник чернично-зеленомошный (P.a.–V.m.: Picea abies–Vaccinium myrtillus–Pleurozium schreberi). В пределы указанных лесорастительных условий укладываются многие типы леса других исследователей (см., например, Kazimirov, 1971; L’vov, Ipatov, 1976; Vasilevich, 1983; 2004). Наиболее точно условия P.a–V.m. соответствуют группе типов ельников зеленомошных В.Н. Сукачева (Sukachev, 1972), группе ассоциаций черничных ельников В.И. Василевича (Vasilevich, 2004), а также Hylocomium-Myrtillus тип и Myrtillus тип А.К. Каяндера (Cajander, 1926).
Исследования проводились в основном на территории Восточной Фенноскандии, где главными лесообразующими породами являются сосна и ель. Здесь в отличие от остальной территории Северо-Запада преобладают сосняки, что в свою очередь объясняется широким распространением песчаных почв. Механический состав почвы является определяющим для доминирования сосны или ели не только из-за различий в плодородии (Kazimirov, 1971), но главным образом из-за коагуляционной способности частиц разной размерности (Kachinskiy, 1965; Morozova, 1991). Именно этот момент является решающим в силу того, что сосна обыкновенная (Pinus sylvestris L.) и ель европейская (Picea abies s.l. (L.) H. Karst.) значительно отличаются строением корневых систем – ель с поверхностными корнями не устойчива на песках. Следует при этом отметить, что в виде исключения ельники черничные могут произрастать на песчаных почвах. Все случаи находят конкретные физические объяснения (рельеф, защищающий от преобладающих ветров, каркас из крупных деревьев сосны, сильнозавалуненные почвы и др.) устойчивости ели от вывалов. Поэтому не случайно мы проводим формальную границу между сосняками и ельниками черничными по границе песчаные–супесчаные почвы, понимая, что она во многом условна. В монографии Н.Г. Федорец с соавторами (Fedorets et al., 2000) отмечается, что ельники черничные характеризуются разнообразием почвенных сочетаний, но в автоморфных условиях увлажнения абсолютное большинство ельников приурочены к подзолам иллювиально-гумусово-железистым и подзолам иллювиально-железистым, а также почвам пятнисто-подзолистым супесчаным и суглинистым, буроземам типичным. По данным Р.М. Морозовой (Morozova, 1991) ельники черничные произрастают на подзолах иллювиально-гумусово-железистых, подбурах и буроземах и что особо подчеркивается – с повышенным содержанием илистых частиц. Ранее (Genikova et al., 2019) мы показали, что в отношении ельников черничных разделение на супесчаные и суглинистые почвы нецелесообразно – небольшое число описаний ельников черничных на суглинках располагалось на ординационной диаграмме, построенной по характеристикам напочвенного покрова, “внутри основного облака”. Этот вывод, однако, требует дальнейшей проверки, так как в регионе исследований кисличные и неморально-травные ельники представлены редкими сообществами в условиях богатых почв вблизи крупных озер Ладожского и Онежского, и необходимо дополнить ряд описаний коренных ельников и производных от них сообществ на суглинках.
Исходя из того, что в потенциале любой фитоценоз, сформировавшийся в определенных лесорастительных условиях, в ходе своего развития станет определенным климаксовым сообществом, мы в пределах одного типа лесорастительных условий выделили сообщества разных возрастных стадий: вырубка (до формирования древесного яруса, что в условиях P.a.–V.m. составляет примерно 5–10 лет), молодняк (в целом соответствует характеристикам в лесоводстве – до 40 лет в средней тайге, до 60 лет в северной; в это время, как правило, происходит самоизреживание древостоя и изменяется структура напочвенного покрова), далее период, продолжающийся до 100–120 лет (соответствует средневозрастным и частично спелым сообществам по параметрам древостоя, верхняя граница определяется продолжительностью жизни березы и осины, а в чистых ельниках очередным периодом активного самоизреживания, что также влечет за собой изменения в структуре напочвенного покрова); далее – до 200–250 лет (в это время в ельниках, развивающихся спонтанно, формируется условно разновозрастный древостой); далее субклимакс и климакс (мы в данном случае их не разделяем, т.к. климаксовые ельники – это в большей степени объекты теоретической фитоценологии при существующей периодичности естественных пожаров и других разрушающих древостой факторов). Аналогичные стадии были выделены С.Н. Дыренковым (Dyrenkov, 1984), но только по признаку возрастной структуры древесного яруса. Нами же этапы развития сообщества определялись по признакам древостоя, подлеска и напочвенного покрова, и мы считаем это принципиальным. Кроме этого, этапы развития ельников могут серьезно отличаться по интенсивности обменных процессов, накоплению и структуре биомассы сообщества, разнообразию не только растительной составляющей, но и других групп организмов (Bobkova et al., 2006; Pristova, 2008; Genikova et al., 2012; Dolgin et al., 2012; Kryshen et al., 2018; Likhanova et al., 2018; Osipov et al., 2019 и др.). Ассоциации выделяются уже в пределах ранее определенных этапов развития. В результате мы в равной степени учитываем условия местообитания (тип лесорастительных условий), видовой состав и физиономию сообщества. Видовой состав определяется одновременно лесорастительными условиями, влажностью почвы (уровень залегания грунтовых вод) и возрастом древостоя. Физиономия (внешний вид) фактически определяется доминантами всех ярусов и возрастом древостоя. Таким образом, наш подход в выделении ассоциации в целом соответствует принятому в 1910 году на Ботаническом конгрессе в Брюсселе определению ассоциации (по Aleksandrova, 1969) и следует отметить, что указанные признаки (видовой состав, лесорастительные условия, возраст) только совместно могут четко обозначить положение сообщества в синтаксономической системе, это особенно справедливо для ранних стадий восстановления.
Такие факторы как экспозиция склона, мезорельеф, временное заболачивание после рубки, изреживание древесного яруса в результате выборочных рубок и т.п. оказывают сильное влияние на состав и структуру растительного сообщества. В итоге даже в пределах одного типа лесорастительных условий повышается ценотическое разнообразие, отражающее временные и локальные состояния. Это разнообразие может быть выражено в широком наборе субассоциаций, которые в данной работе не обсуждаются.
МАТЕРИАЛ И МЕТОДЫ
Описания лесных сообществ для создания эколого-динамической типологии растительных сообществ в условиях ельников черничных (Picea abies – Vaccinium myrtillus (P.a.–V.m.)) выполнялись с 2002 г. на территории Республики Карелия, незначительное количество описаний выполнено также в Архангельской области. Ставилась задача максимально отразить ценотическое разнообразие на всех стадиях развития от рубки до климакса в исследуемых условиях. Такой подход не предполагал большой повторности описаний сходных сообществ в аналогичных и близких по географическому положению условиях. Важно было выявить различные факторы, определяющие варианты развития сообществ после катастрофических нарушений (главным образом сплошных рубок). Участки для описаний выбирались в удалении от берегов озер и рек, исключалось непосредственное примыкание к дорогам, постройкам и другим антропогенным местообитаниям, влияние которых могло помешать четкому определению основных причин формирования структуры сообщества. Всего проанализировано 196 описаний, которые распределены неравномерно по возрастным стадиям, что обусловлено историей освоения лесов – сплошные рубки получили широкое распространение на территории Карелии с 50–60-х годов прошлого столетия, и поэтому в настоящее время преобладают молодняки, в значительно меньшей степени – средневозрастные и еще меньше спелых насаждений.
В случае небольшого по размерам выдела геоботаническое описание сообщества проводилось в его границах, при относительно больших размерах однородного выдела определялась наиболее характерная его часть и в зависимости от возраста, полноты и состава древостоя описывался участок, достаточный для отражения структуры древостоя и напочвенного покрова (не менее 400 кв. м). При описании древесного яруса учитывались состав, возраст, высота, диаметр, полнота древостоя, состав и обилие подлеска и подроста. На каждом участке выполнялись почвенный разрез и описание напочвенного покрова. Видовой состав исследовался на территории выдела. Описания выполнялись общепринятыми методами. На каждом участке определялась его “история”– антропогенные и естественные нарушения (масштабы и сроки) по комплексу признаков (почвенный разрез, наличие и состояние пней, состав и структура древесного яруса, наличие и характер повреждений деревьев, состав и структура напочвенного покрова). Для анализа использовались характеристики напочвенного покрова, для уточнения положения сообществ в классификационной схеме использовались также данные по почвам и продуктивности древостоя.
Для определения факторов, влияющих на формирование сообществ в разных типах лесорастительных условий и разного возраста, и проверки гипотезы об усилении влияния древостоя с возрастом растительного сообщества, проведена ординация геоботанических описаний растительных сообществ на основе бестрендового анализа соответствия – DCA (Hill, 1979) с помощью программы PC-ORD. В анализе учитывалось проективное покрытие видов мохово-лишайникового и травяно-кустарничкового ярусов. Нагрузки на оси рассчитаны с использованием коэффициента Съеренсена.
Наименование ассоциаций мы приводим по предложенному В.В. Алехиным (Alekhin, 1951) принципу – латинские наименования индицирующих видов (как правило, доминантов) древесного, травяно-кустарничкового и мохово-лишайникового ярусов. Ярусы разделяются “–”, а знаком “+" объединяются виды, в пределах яруса одновременно индицирующие ассоциацию. Добавление возраста и отнесение синтаксона к конкретным лесорастительным условиям (указывается перед собственно именованием синтаксона), делает наименование максимально информативным, указывая доминантов всех ярусов, тип местообитания и стадию развития.
РЕЗУЛЬТАТЫ
Всего в условиях ельников черничных нами учтены 30 видов напочвенных мхов и лишайников, 240 видов сосудистых растений. Анализ ценофлоры ельников черничных нами обсуждался ранее (Genikova et al., 2019). Здесь мы лишь коротко повторим некоторые моменты, важные для понимания динамики сообществ и их классифицирования.
Повсеместно в условиях ельников черничных преобладают зеленые мхи Pleurozium schreberi (Brid.) Mitt., Dicranum spp., Hylocomium splendens (Hedw.) Bruch et al., мозаика которых определяется влиянием древесного яруса. В окнах и на повышениях мезорельефа могут временно доминировать кустистые лишайники, а в понижениях – мхи р. Sphagnum. Видовое разнообразие сосудистых растений в условиях ельников черничных более чем в 2 раза выше, чем в сосняках (Genikova et al., 2012) при том, что площадь последних в Карелии в 1.5 раза больше (Yurkovskaya, Elina, 2009).
В условиях ельников черничных наибольшее видовое разнообразие наблюдается на ранних стадиях восстановления растительности, наименьшее – в старовозрастных (старше 100–120 лет) лесах (табл. 1). Общими для ценофлор растительных сообществ всех возрастных стадий являются 53 вида сосудистых растений. Только на вырубках обнаружено 79 видов растений (33% от всей ценофлоры ельников черничных), в молодняках – 10 видов, в 60–120-летних – 15. В лесах с древостоем старше 100–120 лет специфичные виды отсутствуют. По результатам анализа адвентивной и аборигенной фракций ценофлоры ельников разных возрастных стадий был сделан вывод о том, что возрастные стадии восстановления ельников черничных обладают характерными свойствами и принципиально отличаются по составу видов, эколого-ценотической и таксономической структурам. Этот вывод еще раз свидетельствует о том, что ассоциации должны выделяться в пределах стадий сукцессии.
Таблица 1.
Видовой состав ассоциаций растительных сообществ на различных стадиях восстановления Table 1. Species composition of associations of plant communities at different stages of restoration
| Вырубки Clear cuttings |
Молодняки (10–40 лет) Young stands (10–40 years) |
41(61)–100(120) лет 41(61)–100(120) years |
121–200 лет 121–200 years |
>200 лет >200 years |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| № ассоциации/№ of association | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| Количество описаний Number of relevés |
10 | 10 | 22 | 8 | 15 | 6 | 20 | 33 | 9 | 15 | 24 | 7 |
| Всего видов сосудистых растений* Number of vascular plant species |
38 | 46 | 133 | 63 | 62 | 40 | 95 | 78 | 70 | 102 | 54 | 30 |
| Всего видов в мохово-лишайниковом ярусе* Number of species in moss-lichen layer |
12 | 9 | 4 | 11 | 25 | 7 | 11 | 24 | 9 | 23 | 25 | 12 |
| Всего видов в травяно-кустарничковом ярусе* Number of species in herb-dwarf shrub layer |
30 | 37 | 120 | 53 | 53 | 30 | 80 | 64 | 58 | 88 | 43 | 24 |
| Число видов по этапам развития* Number of species by successional stages |
200 | 119 | 129 | 71 | ||||||||
| Древесный ярус и подрост/Tree stand and regrowth | ||||||||||||
| Acer platanoides L. | – | – | I | – | – | – | – | – | – | – | – | – |
| Alnus incana (L.) Moench | – | – | II | I | I | II | I | II | I | – | ||
| Betula pendula Roth | V | IV | V | V | V | V | V | V | V | V | V | V |
| Picea abies (L.) H. Karst. | V | IV | II | V | V | V | V | V | V | V | V | V |
| Pinus sylvestris L. | II | III | II | III | IV | V | IV | IV | V | III | V | III |
| Populus tremula L. | I | II | II | I | II | III | IV | IV | III | II | III | – |
| Подлесок/Undergrowth | ||||||||||||
| Daphne mezereum L. | – | – | – | – | – | I | I | I | I | – | – | – |
| Frangula alnus Mill. | – | – | I | – | – | – | – | I | III | I | – | – |
| Juniperus communis L. | II | I | – | – | I | II | I | II | III | II | II | III |
| Padus avium Mill. | – | – | – | – | – | – | I | – | II | I | – | – |
| Ribes spicatum E. Robson | – | – | – | – | – | – | I | – | – | I | – | – |
| Rosa acicularis Lindl. | II | I | – | II | – | – | I | I | I | I | I | III |
| Rosa majalis Herrm. | – | – | I | – | – | – | – | – | – | – | – | – |
| Rubus idaeus L. | – | I | II | I | I | I | II | I | II | I | I | – |
| Salix aurita L. | – | – | I | I | – | – | – | I | – | – | – | – |
| Salix caprea L. | – | – | II | – | III | III | IV | II | – | I | I | – |
| Salix phylicifolia L. | I | II | II | – | – | II | I | – | – | – | I | – |
| Sorbus aucuparia L. | V | IV | V | III | V | V | IV | V | V | V | V | IV |
| Мохово-лишайниковый ярус/Moss-lichen layer | ||||||||||||
| Cetraria islandica (L.) Ach. | – | – | – | – | I+ | – | – | I+ | – | – | – | – |
| Cladonia spp. | I+ | I+ | – | – | III1–2 | III1–2 | – | II+ | I+ | I+ | I+ | II+ |
| Peltigera aphthosa (L.) Willd. | – | – | – | – | I+ | – | I+ | I+ | – | I+ | I+ | III+ |
| Aulacomnium palustre (Hedw.) Schwägr. | I+ | – | – | I+ | I+ | – | – | I+ | – | I+ | I+ | – |
| Climacium dendroides (Hedw.) F. Weber & D. Mohr | – | – | – | – | – | – | – | – | – | I+ | – | – |
| Dicranum spp. | III1 | II1 | – | IV1 | IV1–2 | II1 | I+ | V1–2 | III1 | III1–2 | V1–2 | V1 |
| Hylocomium splendens (Hedw.) Bruch et al. | III1–2 | II1 | – | III1 | IV1–2 | I1 | II1 | V1–2 | II1–2 | III1–2 | V2–3 | V2–3 |
| Mnium sp. | – | – | – | – | I+ | – | – | I+ | I+ | I+ | – | – |
| Plagiothecium laetum Bruch et al. | – | – | – | – | – | – | – | I1 | I+ | I+ | – | |
| Pleurozium schreberi (Brid.) Mitt. | IV1–2 | II1–2 | I1 | IV1–2 | V2–3 | III2–3 | II1–2 | V1–3 | IV1–2 | IV1–3 | V2–3 | V2 |
| Polytrichum commune Hedw. | IV1–2 | IV2 | I2 | IV1 | II2 | III2 | I1–2 | III1–2 | II+ | I1 | III1 | V1–2 |
| Polytrichum juniperinum Hedw. | I1 | – | I+ | I+ | III+ | II1 | II+ | II+ | – | I+ | I+ | – |
| Ptilium crista-castrensis (Hedw.) De Not. | II+ | I+ | – | II+ | II+ | – | – | II+ | I+ | I+ | I+ | IV+ |
| Rhytidiadelphus triquetrus (Hedw.) Warnst. | – | – | – | I+ | I+ | – | I+ | II+ | I+ | II+ | II+ | I+ |
| Rhodobryum roseum (Hedw.) Limpr. | – | – | – | – | I+ | – | – | II+ | – | I+ | I+ | – |
| Sphagnum spp. | II1 | III1 | I2 | IV2 | III+ | – | I1 | III1–2 | – | II+ | III1–2 | V2 |
| Травяно-кустарничковый ярус/Herb-dwarf shrub layer | ||||||||||||
| Achillea millefolium L. | – | I+ | – | – | I+ | – | – | – | I+ | I+ | – | – |
| Aegopodium podagraria L. | – | – | – | – | – | – | I1 | I+ | I+ | – | – | – |
| Agrostis capillaris L. | – | I1 | II2 | I1 | I+ | II+ | I2 | I+ | II1 | II1 | – | – |
| Alchemilla acutiloba Opiz | – | – | I+ | – | – | – | I+ | – | – | I+ | – | – |
| Alopecurus aequalis Sobol. | – | I+ | I+ | – | – | – | – | – | – | – | – | – |
| Angelica sylvestris L. | – | – | II+ | – | I1 | – | III1 | I+ | II+ | II+ | I+ | – |
| Anthoxanthum odoratum L. | – | – | I+ | – | I+ | – | – | I+ | – | – | – | – |
| Anthriscus sylvestris (L.) Hoffm. | – | – | II+ | – | – | – | II1–2 | – | II+ | I+ | – | – |
| Athyrium filix-femina (L.) Roth | – | – | I+ | – | – | – | I+ | I+ | – | I+ | – | – |
| Avenella flexuosa (L.) Drej. | V2–3 | V3–4 | IV1–2 | V1–2 | V1 | V3 | III1–2 | IV+–2 | V2 | III+–2 | V+–1 | V+–1 |
| Calamagrostis arundinacea (L.) Roth | I2 | II1 | V2–3 | I3 | III1 | III2 | V2–3 | IV1 | V2 | IV1–2 | II1 | – |
| Calamagrostis epigeios (L.) Roth | – | – | I+ | – | I+ | I2 | I2 | I+ | I+ | I2 | I+ | – |
| Calamagrostis phragmitoides C. Hartm. | I+ | – | I2 | – | – | I2 | – | I+ | – | I+ | – | – |
| Callitriche palustris L. | – | – | I+ | – | – | – | – | – | – | – | – | – |
| Calluna vulgaris (L.) Hull | I1 | I1 | I+ | – | III+–1 | I2 | I1 | I+ | I+ | – | I+ | – |
| Campanula patula L. | – | – | I+ | – | – | – | I1 | – | – | – | – | – |
| Campanula persicifolia L. | – | – | I+ | – | – | – | I1 | – | – | – | – | – |
| Campanula rotundifolia L. | – | – | – | – | – | – | – | – | II+ | I+ | – | – |
| Carex acuta L. | – | – | I+ | – | – | – | – | – | – | – | – | – |
| Carex brunnescens (Pers.) Poir | I1 | II+ | I2 | II2–3 | – | – | – | – | – | – | I2 | – |
| Carex canescens L. | – | – | I1 | I3 | – | – | I+ | – | I+ | I+ | – | – |
| Carex globularis L. | I1 | I2 | I1 | IV2 | II+ | – | I+ | I+ | – | I+ | II+ | V+ |
| Carex leporina L. | – | I+ | II+ | II+ | – | – | – | – | – | – | – | – |
| Carex nigra (L.) Reichard | – | – | I+ | II1 | – | – | – | – | – | – | – | – |
| Carex pallescens L. | – | I+ | I+ | – | – | – | – | – | – | I+ | – | – |
| Carex vaginata Tausch | – | – | – | – | I+ | – | – | I+ | – | I1 | – | – |
| Chamaenerion angustifolium (L.) Scop. | V1–2 | V1–2 | IV1–2 | V1–2 | III+ | V1–2 | V1–2 | I+ | I+ | II+ | I+ | I+ |
| Chamaepericlymenum suecicum (L.) Asch. & Graebn. | I2 | – | – | II+ | I+ | – | – | I+ | – | – | I+ | – |
| Cirsium heterophyllum (L.) Hill | – | – | I+ | – | – | – | I+ | I+ | – | I+ | – | – |
| Cirsium palustre (L.) Scop. | – | – | I+ | – | – | – | – | – | – | – | – | – |
| Convallaria majalis L. | I1 | – | II+ | – | I+ | I2 | III1–2 | II+–1 | IV1–2 | III+–1 | I+ | – |
| Deschampsia cespitosa (L.) Beauv. | – | I+ | I+ | – | – | I1 | I1 | I+ | – | I1 | – | – |
| Dryopteris carthusiana (Vill.) H.P. Fuchs | I+ | – | II+ | II1 | II+ | I+ | II+–1 | III+ | III+ | IV+ | I+ | – |
| Dryopteris expansa (C. Presl) Fraser-Jenk. & Jermy | – | I+ | I1 | – | – | – | – | I+ | – | I1 | I+ | – |
| Empetrum nigrum L. s.l. | I1 | – | – | II1 | II1 | I+ | – | I+ | – | – | III+ | III1 |
| Epilobium palustre L. | – | – | II+ | I+ | – | – | – | – | – | – | – | – |
| Equisetum sylvaticum L. | II+ | III+ | II1 | IV1 | I1 | – | II+ | II+ | I2 | III+–1 | III+ | V+ |
| Eriophorum vaginatum L. | – | I+ | – | I+ | – | – | I+ | – | – | – | – | – |
| Filipendula ulmaria (L.) Maxim. | – | – | I+ | I1 | – | – | I+ | – | – | I1 | I+ | – |
| Fragaria vesca L. | – | – | II+–1 | – | I+ | I2 | II1 | I+ | III+–1 | II+ | – | – |
| Galium album Mill. | – | – | I+–1 | – | – | I1 | II+–1 | – | – | I+ | – | – |
| Geranium sylvaticum L. | I+ | – | III+–1 | II+ | I+ | I1 | III1–2 | I+ | II+ | II+–1 | I+ | I+ |
| Goodyera repens (L.) R. Br. | – | – | – | – | – | – | – | I+ | II+ | – | I+ | – |
| Gymnocarpium dryopteris (L.) Newm. | I+ | I+ | II+–1 | II+–1 | I1 | III1 | I1 | III+–1 | – | II+–1 | I+–1 | I+ |
| Hepatica nobilis Mill. | – | – | – | – | – | – | – | I+ | II1 | – | – | – |
| Hieracium umbellatum L. | – | – | I+ | – | I+ | I+ | – | I+ | II+ | I+ | – | – |
| Hieracium vulgatum Fries | – | – | I+ | – | II+ | – | – | I+ | II+ | I+ | I+ | – |
| Hypericum maculatum Crantz | – | – | I+ | – | – | – | II+ | – | I+ | I+ | – | – |
| Juncus effusus L. | – | – | II1–2 | – | – | – | – | – | – | – | – | – |
| Juncus filiformis L. | – | I+ | I+ | I+ | – | – | I+ | – | – | – | – | – |
| Lamium album L. | – | – | I+ | – | – | – | – | – | – | – | – | – |
| Lathyrus vernus (L.) Bernh. | I+ | – | I+ | – | – | – | I1 | I+ | II+ | I+ | I+ | – |
| Ledum palustre L. | – | – | – | II2 | I+ | – | – | I+ | – | – | I+ | – |
| Linnaea borealis L. | III+ | I+ | I+ | III+ | II+ | I+ | I+ | III+ | II+ | II+ | V+ | V+ |
| Listera cordata (L.) R. Br. | – | – | – | I+ | – | – | – | I+ | – | – | I+ | – |
| Luzula pilosa (L.) Willd. | IV+ | IV+–1 | III+–1 | II+ | III+ | V+–1 | II+ | IV+ | IV+ | II+ | V+ | IV+ |
| Lycopodium annotinum L. | II+ | – | I+ | II+ | III+ | I+ | I+ | II+ | II+ | II+ | II+ | III+ |
| Maianthemum bifolium (L.) F.W. Schmidt | III1 | II+ | II+ | III1 | III+ | IV+ | III+–1 | IV+–1 | V1 | IV+–1 | III+–1 | III+ |
| Melampyrum pratense L. | III+ | I+ | II+–1 | IV+ | IV+–1 | IV2 | IV1–2 | IV+–1 | V+–1 | III+–1 | III+ | IV+ |
| Melampyrum sylvaticum L. | III+ | I+ | – | I+ | II+ | I+ | I+ | III+–1 | IV+–1 | III+–1 | II+ | III+ |
| Melandrium dioicum (L.) Coss. & Germ. | – | – | – | – | – | – | – | – | – | I+ | – | – |
| Melica nutans L. | – | – | I+ | – | – | – | II+–1 | I+ | II+ | I+ | I+ | – |
| Milium effusum L. | – | – | I+ | – | – | – | – | – | – | II+ | – | – |
| Moehringia trinervia (L.) Clairv. | – | – | – | – | – | – | I+ | – | – | I+ | – | – |
| Omalotheca sylvatica (L.) Sch. Bip. & F. Schultz | – | I+ | I+ | – | – | – | – | – | – | – | – | – |
| Orthilia secunda (L.) House | – | I+ | I+ | I+ | II+ | – | I+ | III+ | II+ | II+ | II+ | III+ |
| Oxalis acetosella L. | I+ | – | II+ | II+ | I+ | – | II1 | III1–2 | III1–2 | IV1–2 | I1 | I+ |
| Paris quadrifolia L. | – | – | I+ | – | – | – | II+ | I+ | II+ | II+ | – | – |
| Phegopteris connectilis (Michx.) Watt | – | – | I+ | – | – | – | – | – | I+ | I+ | I+ | – |
| Platanthera bifolia (L.) Rich. | – | – | – | – | I+ | – | I+ | I+ | II+ | I+ | I+ | – |
| Potentilla erecta (L.) Raeusch. | – | I+ | II+ | I+ | I+ | – | I+ | I+ | I+ | I+ | – | – |
| Pteridium latiusculum (Desv.) Hieron. ex Fries | – | – | – | – | – | – | I+ | I+ | II+ | – | – | – |
| Pyrola minor L. | – | – | I+ | – | I+ | – | – | I+ | – | I+ | – | – |
| Pyrola rotundifolia L. | – | – | I+ | – | – | – | – | I+ | – | II1 | – | I+ |
| Ranunculus acris L. | – | – | I+ | I+ | I+ | – | I+ | – | – | I+ | – | – |
| Ranunculus repens L. | – | – | I+ | – | – | – | I+ | – | – | I+ | – | – |
| Rubus arcticus L. | I+ | I+ | I+ | II1 | – | – | – | – | – | I+ | – | II+ |
| Rubus chamaemorus L. | – | I+ | – | II+ | – | – | – | – | – | – | I+ | III+ |
| Rubus saxatilis L. | – | I+ | IV1 | I+ | II1 | – | IV1–2 | III+–1 | V1–2 | III1 | II+–1 | – |
| Rumex acetosella L. | – | I+ | I+ | I1 | – | – | – | – | – | – | – | – |
| Solidago virgaurea L. | III1 | II1 | IV+–1 | I+ | IV+–1 | V1 | IV1 | IV1 | IV1 | III1 | III+ | II+ |
| Stellaria graminea L. | – | – | I+ | I+ | – | – | I1 | – | I+ | I+ | – | – |
| Taraxacum officinale L. | – | – | I+ | – | – | – | I+ | – | – | – | – | – |
| Trientalis europaea L. | III+ | I+ | I+ | IV+ | III+ | I+ | III+ | III+ | V+ | IV+ | III+–1 | IV+ |
| Trifolium pratense L. | – | – | I+ | – | – | – | I1 | – | – | – | – | – |
| Trifolium repens L. | – | – | I+ | – | – | – | I1 | I+ | – | – | – | – |
| Trollius europaeus L. | – | – | I+ | – | – | – | I+ | – | – | I1 | – | – |
| Tussilago farfara L. | – | – | I+ | – | – | – | I+ | – | – | – | – | – |
| Urtica dioica L. | – | – | I+ | – | – | – | I+ | – | – | – | – | – |
| Vaccinium myrtillus L. | V1–2 | IV+–1 | V+–1 | V1 | V2 | V1–2 | IV1 | V2–3 | V2 | IV1–2 | V3–4 | V3–4 |
| Vaccinium uliginosum L. | II+ | I+ | – | I1 | I1 | I+ | – | I1 | – | – | I1–2 | II1 |
| Vaccinium vitis-idaea L. | V2 | IV1 | III+–1 | V2 | V1–2 | V1–2 | III1–2 | V1–2 | V1–2 | III+–1 | V2 | V2 |
| Veronica chamaedrys L. | – | – | I+ | – | I+ | – | II+ | – | II+ | II+ | – | – |
| Veronica officinalis L. | – | I+ | I+ | – | I+ | – | I+ | I+ | II+ | I+ | – | – |
| Vicia sepium L. | – | – | – | – | – | – | I+ | – | II+ | I+ | – | – |
| Vicia sylvatica L. | – | – | – | – | – | – | I1 | – | – | – | – | – |
| Viola epipsila Ledeb. | – | – | – | – | – | – | – | I+ | – | I+ | – | – |
| Viola mirabilis L. | – | – | – | – | – | – | I+ | I+ | II+ | – | – | – |
| Viola nemoralis Kütz. | – | – | I+ | – | I+ | – | II1 | I+ | II+ | I+ | – | – |
| Viola riviniana Reichenb. | – | – | I+ | – | – | – | – | – | – | – | – | – |
| Viola tricolor L. | – | – | I+ | – | – | – | – | – | – | – | – | – |
Примечание. * – приводится информация о всех учтенных видах, а не только тех, которые представлены в таблице – в таблице не указаны виды, встреченные единично в 1–2 описаниях.
Классы постоянства (%): I – менее 20; II – 21–40; III – 41–60; IV – 61–80; V – 81–100. В верхнем индексе указано обилие вида по шкале Браун–Бланке: “+” – вид редкий и имеет малое проективное покрытие до 1%; 1 – проективное покрытие вида 1 – 5%; 2 – 5–25%; 3 – 25–50%; 4 – 50–75%; 5 – выше 75%.
Note. * – the number of species is that of all recorded species, not only of those presented in the table; the table does not specify the species encountered singly in 1–2 relevés.
Constancy classes (%): I – less than 20; II – 21–40; III – 41–60; IV – 61–80; V – 81–100. The upper index indicates the species abundance on the Braun–Blanquet scale: + – the species is rare and has a projective cover less than 1%; 1 – the projective cover of the species is 1–5%; 2 – 5–25%; 3 – 25–50%; 4 – 50–75%; 5 – above 75%.
Названия ассоциаций/Names of plant communities associations:
1 – P.a.–V.m.: Vaccinium vitis-idaea;
2 – P.a.–V.m.: Avenella flexuosa;
3 – P.a.–V.m.: Calamagrostis arundinacea;
4 – P.a.–V.m.: Carex canescens;
5 – P.a.–V.m.: Betula spp.+Pinus sylvestris–Vaccinium myrtillus–Pleurozium schreberi;
6 – P.a.–V.m.: Betula spp.+Populus tremula+Picea abies–Avenella flexuosa;
7 – P.a.–V.m.: Betula spp.+Populus tremula+Picea abies–Calamagrostis arundinacea+Geranium sylvaticum;
8 – P.a.–V.m.: Picea abies+Betula spp.+Pinus sylvestris–Vaccinium myrtillus–Pleurozium schreberi;
9 – P.a.–V.m.: Picea abies+Betula spp.– Vaccinium myrtillus+Calamagrostis arundinacea–Hylocomium splendens;
10 – P.a.–V.m.: Picea abies+Betula spp.–Calamagrostis arundinacea+Gymnocarpium dryopteris;
11 – P.a.–V.m.: Picea abies–Vaccinium myrtillus–Pleurozium schreberi;
12 – P.a.–V.m.: Picea abies–Vaccinium myrtillus–Pleurozium schreberi.
Число видов на ранних стадиях развития сообществ после катастрофических разрушений во многом состоит из непостоянной составляющей (Kryshen, 2003, 2006). Состав таких сообществ в большей степени определяется окружением и удаленностью сообщества от населенных пунктов и сельхозземель и число видов здесь будет ожидаемо увеличиваться за счет синантропной составляющей флоры (см. Kryshen et al., 2016).
В основе эколого-динамической модели лежит тезис о том, что развитие лесного сообщества идет по пути восстановления коренного (климаксного), характеризующего лесорастительные условия. Структура и динамика (главным образом древесного яруса) коренных ельников черничных описана во многих работах (Dyrenkov, 1984; Volkov, 2003; Shorohova et al., 2009; Kuuluvainen, Aacala, 2011; Kuuluvainen et al., 2014; Kreutz et al., 2015 и др.). Береза и осина, присутствующие в древостое – следствие оконной динамики, а единичные сосны, часто, старшего, чем ель возраста – свидетельства прошлого пожара. Подрост в ельниках черничных в зависимости от условий может быть и редким, и обильным, как правило, еловый, в окнах – с березой и осиной. В подлеске обычны Sorbus aucuparia L., Salix caprea L. и Juniperus communis L. В средней тайге встречается шиповник (Rosa acicularis Lindl). Кроме кустарничков в коренных ельниках черничных в травяно-кустарничковом ярусе присутствуют и могут доминировать в окнах Avenella flexuosa (L.) Drej., Convallaria majalis L., Calamagrostis arundinacea (L.) Roth, Geranium sylvaticum L. Обычны также Linnaea borealis L., Maianthemum bifolium (L.) F.W. Schmidt, Melampyrum pratense L., Oxalis acetosella L., Rubus saxatilis L., Solidago virgaurea L., Trientalis europaea L.
Ельники черничные являются одной из основных составляющих лесоэксплуатационного фонда на Северо-Западе России, поэтому в этих лесорастительных условиях большие площади вырубок. Для сообществ в условиях Picea abies – Vaccinium myrtillus (P.a.–V.m.) на стадии вырубки нами выделено 4 ассоциации (перечислены в порядке увеличения влажности почвы): 1.11 брусничная (P.a.–V.m.: Vaccinium vitis-idaea), 2. луговиковая (P.a.–V.m.: Avenella flexuosa), 3. вейниковая (P.a.–V.m.: Calamagrostis arundinacea) и 4. осоковая (P.a.–V.m.: Carex canescens) (табл. 1).
В условиях ельников черничных на вырубках нами не выделяется (как в сосняках) “центральная” ассоциация, которая образуется в случае незначительных изменений влажности верхних горизонтов почвы после рубки. Дело в том, что здесь наблюдается разделение на среднюю и северную тайгу – в первом случае вырубки зарастают вейником лесным, а луговик вытесняется на наиболее сухие, или, наоборот, на переувлажненные участки. По всей видимости луговик имеет более широкую экологическую амплитуду, чем вейник лесной, но не способен конкурировать с последним в условиях его фитоценотического оптимума. Таким образом, мы имеем две ассоциации сообществ, образующихся на вырубках ельников черничных при незначительных изменениях увлажнения: 3. P.a.–V.m.: Calamagrostis arundinacea (средняя тайга) и 2. P.a.–V.m.: Avenella flexuosa (северная тайга, средняя тайга – при снижении влажности почвы). В сообществах обеих ассоциаций указанным индикаторным видам практически всегда содоминирует иван-чай, обильны таежные кустарнички, мелкотравье, золотарник и др. Естественное возобновление древесных пород на вырубках данных ассоциаций состоит преимущественно из Betula spp.
Рубка ельников черничных в верхних частях пологих склонов южной и юго-западной экспозиции приводит к иссушению верхних слоев почвы. Следствием является формирование сообществ с доминированием таежных кустарничков, в первую очередь, брусники (ассоциация 1. P.a.–V.m.: Vaccinium vitis-idaea). Невысокое обилие злаков позволяет сохранить свое присутствие таежному мелкотравью, зеленым мхам и лишайникам. Такие вырубки возобновляются в равной степени Betula spp. и Picea abies.
В нижних частях склонов, наоборот, уничтожение древесного яруса приводит к увеличению увлажнения. Характерными видами в таких условиях являются осоки, сфагновые и политриховые мхи (ассоциация 4. P.a.–V.m.: Carex canescens). Кроме этого, отсутствие конкуренции со стороны злаков позволяет сохраняться с достаточно высоким обилием таежным кустарничкам и разнотравью. Подрост древесных пород состоит из Betula spp. и Picea abies.
На следующем этапе, в целом соответствующем молоднякам в лесоводстве, развитие напочвенного покрова идет уже под влиянием формирующегося древесного яруса. Естественное восстановление древостоя в условиях ельников черничных практически всегда идет через смену пород. Восстанавливающийся древостой с одной стороны упрощает структуру напочвенного покрова, выравнивая среду под пологом. С другой – сам древесный ярус неоднороден, и на ранних стадиях здесь могут содоминировать ель, сосна, береза и осина. Структура древесного яруса определяется в первую очередь составом вырубленного древостоя, а также способом рубки, проведенными мероприятиями по восстановлению леса, уходами. В то же время исследования показали, что все сообщества достаточно четко “укладываются” в три ассоциации: 5. P.a.–V.m.: Betula spp.+Pinus sylvestris–Vaccinium myrtillus–Pleurozium schreberi, 6. P.a.–V.m.: Betula spp.+Populus tremula+Picea abies–Avenella flexuosa, 7. P.a.–V.m.: Betula spp.+Populus tremula+Picea abies–Calamagrostis arundinacea+Geranium sylvaticum (табл. 1, рис. 1).
Рис. 1.
Ординационная диаграмма геоботанических описаний молодняков (ассоциации №№ 5–7) в лесорастительных условиях ельников черничных (A). Б, В, Г, Д, Е – та же ординационная диаграмма, где размером значка, обозначающего геоботаническое описание сообщества, показано участие определенного вида растений в сложении напочвенного покрова (чем больше значок, тем выше проективное покрытие вида): Б – Vaccinium vitis-idaea, В – V. myrtillus, Г – Pleurozium schreberi, Д – Avenella flexuosa, Е – Calamagrostis arundinacea.
Fig. 1. Ordination diagram of geobotanical relevés of young forest (associations 5–7) in bilberry spruce conditions (A). Б, В, Г, Д, Е – the same ordination diagram, where the circle size cooresponds to the participation of a certain plant species in the ground cover in geobotanical relevés of the community (the larger circle the higher coverage of the species): Б – Vaccinium vitis-idaea, В – V. myrtillus, Г – Pleurozium schreberi, Д – Avenella flexuosa, Е – Calamagrostis arundinacea. Ось – axis.
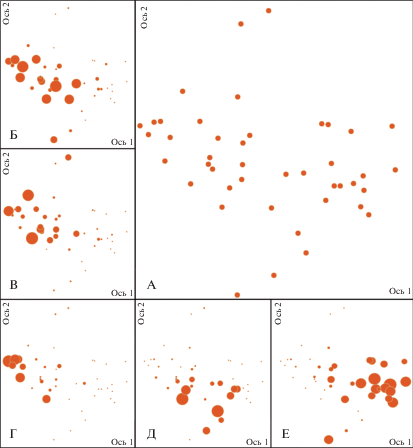
Сообщества ассоциации 5. P.a.–V.m.: Betula spp.+Pinus sylvestris–Vaccinium myrtillus–Pleurozium schreberi (березняки черничные) формируются на вырубках, отнесенных нами к ассоциации 1. P.a.–V.m.: Vaccinium vitis-idaea. В этих условиях при естественном зарастании древесный ярус складывается, главным образом, березой и сосной. Развитие напочвенного покрова идет по пути смены светолюбивой брусники теневыносливой черникой. Ее среднее покрытие уже даже несколько выше, чем у брусники, и ее экологический ареал более четко разделяется с луговиком, индицирующим другую ассоциацию (рис. 1). Кроме таежных кустарничков достаточно обильны Melampyrum pratense (может содоминировать), Solidago virgaurea, Trientalis europaea, Maianthemum bifolium, Chamaenerion angustifolium, а также злаки Avenella flexuosa и Calamagrostis arundinacea, особенно обильные в окнах. Состав древостоя – смешанный с участием Picea abies, Betula spp. и Pinus sylvestris.
Ассоциация 6. P.a.–V.m.: Betula spp.+Populus tremula+Picea abies–Avenella flexuosa образуется на луговиковых вырубках. В напочвенном покрове помимо луговика обильны Calamagrostis arundinacea и Chamaenerion angustifolium. C формированием древесного яруса возрастает обилие лесных видов Vaccinium myrtillus, Gymnocarpium dryopteris, Solidago virgaurea, Trientalis europaea, Maianthemum bifolium, Luzula pilosa, присутствующих практически во всех описаниях, но пока еще с невысоким проективным покрытием. В древостое преобладает Betula spp.
Ассоциация 7. P.a.–V.m.: Betula spp.+Populus tremula+Picea abies–Calamagrostis arundinacea+Geranium sylvaticum занимает крайнее положение в ряду увлажнения и продуктивности местообитания. Сообщества этой ассоциации формируются на вейниковой и осоковой вырубках. Древесный ярус с каждым годом усиливает свое влияние, становясь ведущим фактором, он выравнивает условия увлажнения, что в свою очередь приводит к замещению осок злаками, таежными кустарничками, разнотравьем, а сфагновых и политриховых мхов – зелеными мхами. Ассоциацию индицируют помимо вейника лесного Geranium sylvaticum, Angelica sylvestris и/или Anthriscus sylvestris, Convallaria majalis, Rubus saxatilis. В древостое преобладают Betula spp. и Populus tremula.
Следующий этап развития совпадает по возрасту древостоя со средневозрастными и частично спелыми лесами, окончание его мы определяем предельным возрастом жизни березы и осины в условиях ельников черничных. Это, как правило, 100–120 лет. В чистых ельниках в этот период часто наблюдается естественное самоизреживание древостоя. Здесь нами выделены также 3 ассоциации: 8. P.a.–V.m.: Picea abies+Betula spp.+Pinus sylvestris–Vaccinium myrtillus–Pleurozium schreberi, 9. P.a.–V.m.: Picea abies+Betula spp.– Vaccinium myrtillus+Calamagrostis arundinacea–Hylocomium splendens, 10. P.a.–V.m.: Picea abies+Betula spp.–Calamagrostis arundinacea+Gymnocarpium dryopteris.
Большинство описаний сообществ от 60 до 100–120 лет в условиях ельников черничных отнесены нами к ассоциации 8. P.a.–V.m.: Picea abies+Betula spp.+Pinus sylvestris–Vaccinium myrtillus–Pleurozium schreberi (табл. 1). Сформировавшийся древесный ярус уже фактически контролирует среду и обеспечивает доминирование типичных таежных видов и зеленых мхов, среди последних наиболее обилен Pleurozium schreberi. Характерным для этой ассоциации является высокая встречаемость при низком проективном покрытии лесных злаков – сообщество постепенно выстраивает структуру напочвенного покрова, типичную для коренных ельников – луговик и вейник в надземной части представлены мелкими вегетирующими особями. Таежное мелкотравье, также как и злаки, имеет высокую встречаемость, но низкое проективное покрытие. Рядом с деревьями березы и осины могут быть обильны ландыш и костяника. В древостое преобладает Picea abies, реже Pinus sylvestris и Betula spp., единично присутствует Populus tremula.
Следующая ассоциация 9. P.a.–V.m.: Picea abies+Betula spp.–Vaccinium myrtillus+Calamagrostis arundinacea–Hylocomium splendens отличается от предыдущей более активным участием березы и, как следствие, увеличенным покрытием Hylocomium splendens и злаков. Из разнотравья к индицирующим видам можно также отнести Convalla-ria majalis и Rubus saxatilis (табл. 1). В древостое преобладают Betula spp. и Populus tremula.
Еще одна ассоциация 10. P.a.–V.m.: Betula spp.+Picea abies–Calamagrostis arundinacea+Gymnocarpium dryopteris характеризуется еще более высоким обилием злаков. Индицирует ассоциацию низкое проективное покрытие таежных кустарничков. В напочвенном покрове множество видов разнотравья, но, как правило, с невысоким обилием. Моховой покров мозаичен – на фоне Pleurozium schreberi пятнами произрастают примерно в равных долях Hylocomium splendens, Dicranum spp., Polytrichum spp., Sphagnum spp. Мозаика определяется структурой древесного яруса и микрорельефом. В древостое преобладают Picea abies, Betula spp., реже Pinus sylvestris.
В сообществах, достигших возраста спелости, в период после естественного отмирания лиственных пород и активного самоизреживания ценопопуляции ели (Alekseev, Molchanov, 1954; Dyrenkov, 1984) формируется напочвенный покров, близкий к климаксовому сообществу. Фактически с этого момента мы выделяем одноименные ассоциации P.a.–V.m.: Picea abies–Vaccinium myrtillus–Pleurozium schreberi, обозначенные в таблице 1 под номерами 11 и 12 в соответствии с этапом развития древостоя, где и происходят основные (качественные) изменения. С достижением елью предельного возраста динамика сообщества определяется локальными разрушениями (оконная динамика). В окнах в зависимости от их размеров могут вырастать и выходить в первый ярус лиственные породы, которые позволяют существовать неопределенно долгое время лесным злакам, разнотравью. Периодические пожары определяют участие сосны в древостое. В древесном ярусе преобладает Picea abies, редко Pinus sylvestris; Betula spp. и Populus tremula представлены незначительно (до 2 единиц по запасу). В целом древостой ели, несмотря на локальные разрушения, имеет относительно стабильную возрастную, породную и размерную структуру (см. также Dyrenkov, 1984; Manov, 2017; 2019) за счет непрерывного возобновления и взаимодействий деревьев.
ОБСУЖДЕНИЕ
Таким образом, в условиях ельников черничных демонстрируется четкая закономерность уменьшения ценотического разнообразия по мере развития древостоя. На рисунке 2 представлена ординация описаний в условиях ельников черничных – конфигурация “облака” имеет форму треугольника, в вершине которого – сообщества коренных лесов. Следует отметить, что интерпретация осей затруднена, т.к. обилие видов напочвенного покрова определяется одновременно увлажнением, плодородием и составом древесного яруса. Кроме этого, все основные древесные породы имеют широкие и значительно накладывающиеся друг на друга экологические ареалы. Здесь же на рисунке 2 продемонстрировано, как виды индицируют ассоциации. Основная масса описаний сообществ старше 120 лет характеризуется доминированием в травяно-кустарничковом ярусе черники, а в мохово-лишайниковом – Pleurozium schreberi. Здесь также обильна брусника, но ее “облако” смещено по обеим осям, и что удивительно, в том же направлении, что и описания с высоким обилием сфагнума. Эти виды индицируют противоположные условия увлажнения, как на вырубках, так и в напочвенном покрове коренных лесов. Объясняться этот момент может тем, что оба вида в исследуемых лесорастительных условиях вытесняются доминантами и на диаграмме они сближены (но не накладываются) по признаку низкого обилия черники, и плеуроциума, или в случае вырубок – вейника и луговика. Говоря о двух поздних этапах развития сообществ, следует отметить еще два момента. Черника доминирует как в старо-, так и в средневозрастных сообществах. Пара старовозрастных сообществ с высоким обилием кислицы (рис. 2) оказалась смещена в зону распространения средневозрастных ельников, причиной изменений в напочвенном покрове явились выборочные рубки, проведенные 40–50 лет назад, в результате которых в первом ярусе по количеству стволов преобладали деревья ели 60 лет, поднявшиеся из подроста. Еще два сообщества, отнесенные нами по возрасту главной породы к старовозрастным, по сути, находились на границе между двух стадий – в период отмирания березы и осины. В образовавшихся окнах естественным образом доминировали лесные злаки, и это объясняет “сдвиг” этих сообществ на диаграмме (рис. 2) в сторону молодняков. Как отмечалось выше, они могут быть отражены в наборе субассоциаций как временные состояния.
Рис. 2.
Ординационная диаграмма геоботанических описаний растительных сообществ в типе лесорастительных условий ельника черничного.
Нагрузки на оси: 1-й фактор – 45%, 2-й – 15.1%. Условные обозначения: 1 – растительные сообщества вырубок, 2 – сообщества до 60 лет, 3 – сообщества с возрастом древостоя от 60 до 100–120 лет, 4 – сообщества с возрастом древостоя больше 120 лет. Вокруг основной диаграммы (А) представлены иллюстрации участия видов в структуре напочвенного покрова (чем больше диаметр кружка, тем выше проективное покрытие вида).
Fig. 2. Ordination diagram of geobotanical relevés in conditions of bilberry spruce forest (A).
Factor contribution: 1st factor (axes) – 45%, 2nd –15.1%. Colored symbols: 1 – plant communities of clear cuttings, 2 – young forest communities under 60 years old, 3 – communities with the age of the stand 60 to 100–120 years, 4 – communities with the age of the stand over 120 years. Around the main diagram, there are pictures of the same ordination diagram where the circle size cooresponds to participation of a certain plant species in the ground cover in the geobotanical relevés of the community (the larger circle the higher coverage of the species).
Б – Vaccinium myrtillus, В – Pleurozium shreberi, Г – Sphagnum spp., Д – Vaccinium vitis-idaea, Е – Rubus saxatilis, Ж – Carex globularis, З – Oxalis acetosella, И – Chamaenerion angustifolium, К – Geranium sylvaticum, Л – Calamagrostis arundinacea, М – Solidago virgaurea, Н – Avenella flexuosa. Ось – axis.
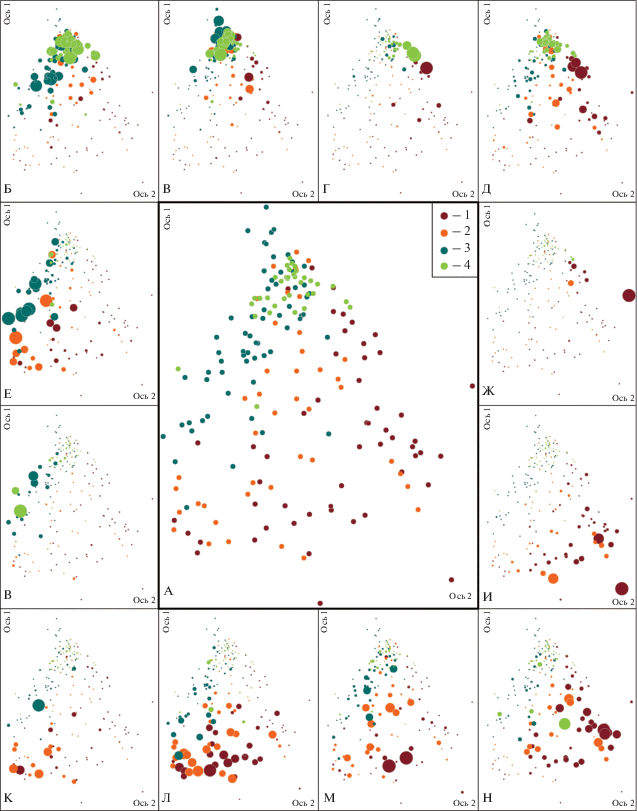
Описания сообществ 40–120 лет расположены на диаграмме (рис. 2) также достаточно компактно, но вытянутым облаком, пересекающим пространство старовозрастных лесов. Напочвенный покров здесь во многом сходен со старовозрастными лесами, но при значительном участии в древесном ярусе лиственных пород или при изреженном древостое в числе доминантов могут быть виды лесного разнотравья: Rubus saxatilis, Geranium sylvaticum, Solidago virgaurea, Gymnocarpium dryopteris и др.
Центральная часть “облака” занята описаниями молодняков, в напочвенном покрове которых содоминируют лесные апофиты, лесное разнотравье и кустарнички, индицируя различные ассоциации, описанные выше.
“Треугольное” облако с двух сторон ограничивают (очерчивают?) описания вырубок с достаточно четкими границами между выделенными ассоциациями. Временно заболачивающиеся участки с доминированием Carex globularis – это достаточно редкий случай, продемонстрированный на рис. 2, – выделены нами в отдельную ассоциацию т.к. их существование стабильно на протяжение всей стадии вырубки и только при формировании сомкнутого древесного яруса на стадии молодняка происходит нормализация увлажнения и перестройка структуры напочвенного покрова. Следует особо прокомментировать позицию распространенного на вырубках Chamaenerion angustifolium, на диаграмме он выделяется очень высоким проективным покрытием в отдельных описаниях участков, пройденных пожаром.
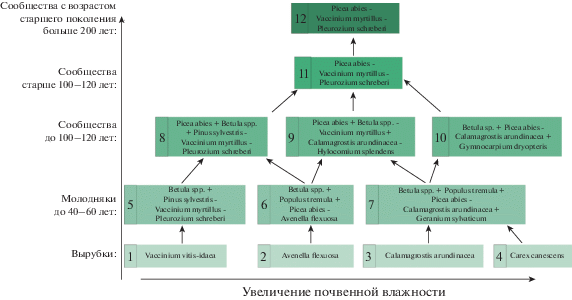
Динамические ряды в условиях ельников черничных отличаются от таковых в условиях сосняков (Kryshen et al., 2018) не только тем, что отсутствует “центральная” ассоциация на вырубках, но и тем, что на определенных этапах может происходить раздвоение направления развития (рис. 3). Обусловлено это не только более широким экологическим ареалом ельников черничных, но и развитием древесного яруса, главным образом, густотой древостоя и породным составом, что в свою очередь может объясняться естественными или антропогенными причинами. В монографии С.Н. Дыренкова (Dyrenkov, 1984) представлена схема, названная им “Основные направления современной динамики южно- и среднетаежных ельников и сопряженности возрастной структуры древостоев с отдельными фазами сукцессии”. Представленные в ней динамические ряды, построенные только по признакам древесного яруса, в целом согласуются с предложенными нами.
Рис. 3.
Восстановительные ряды растительных ассоциаций в условиях ельников черничных (P.a.–V.m.).
Fig. 3. Restoration series of plant associations in bilberry spruce forests (P. a.–V. m.). Увеличение почвенной влажности – soil moisture increase. Вырубки – clear cuttings. Молодняки до 40– 60 лет – young forests 40–60 years old. Сообщества до 100–120 лет – communities aged up to 100–120 years. Сообщества с возрастом старшего поколения больше 200 лет – communities with the age of oldest trees of stand over 200 years.
Типологии многих авторов предлагают различное число типов леса в пределах условий ельников черничных. Так Ф.С. Яковлев и В.С. Воронова (Yakovlev, Voronova, 1959) выделили для Карелии северотаежные воронично-черничный и воронично-брусничный ельники, отличающиеся друг от друга большим или меньшим участием черники или брусники. Н.И. Казимиров (Kazimirov, 1971) выделил также ельник черничный свежий и ельник брусничный на супесчаных почвах. У Т.К. Юрковской (Yurkovskaya, 1993) определены ассоциации ельник воронично-черничный и воронично-брусничный или просто брусничный. Действительно в ельниках черничных содоминантом в травяно-кустарничковом ярусе выступает брусника, ее обилие зависит от рельефа и микрорельефа, от стадии развития сообщества, проведенных лесоводственных мероприятий и т.п. В северной тайге часто встречаются сообщества с приблизительно равным обилием Vaccinium vitis-idaea, V. myrtillus, V. uliginosum, Empetrum nigrum s.l., и преобладание на отдельных участках или в отдельные периоды развития сообщества того или иного вида, на наш взгляд, может быть выражением временных или локальных состояний, не отражающих общих закономерностей формирования сообщества. Кроме того, здесь мы сталкиваемся с общеизвестным принципом – географические (климатические, зональные) черты сообществ могут компенсироваться плодородием почвы. В северной тайге на наиболее плодородных участках встречаются типичные по продуктивности и составу всех ярусов среднетаежные сообщества, справедлива и обратная закономерность. Ряд исследователей представляли еще более дробное деление на типы в пределах исследуемых лесорастительных условий (Chertovskoj, 1978; Rysin, Savel’eva, 2002 и др.), что объясняется не только стремлением отразить все разнообразие сообществ, но и тем, что шире был географический охват и не учитывалась динамика (типы не выстраивались в динамические ряды, а представлялись одним комплексом). С практической точки зрения большое число синтаксонов делает невозможным не только прикладной аспект, но и маскирует основные факторы динамики растительности множеством второстепенных, затрудняя определение закономерностей развития сообщества. Поэтому мы посчитали логичным при выделении ассоциаций в пределах типа лесорастительных условий и этапов восстановления ограничиться только реакцией видов древесного, травяно-кустарничкового и мохово-лишайникового ярусов на изменение условий увлажнения. В результате же все три признака (тип лесорастительных условий, возраст, влажность–продуктивность) индицируются обилием определенных видов, набор которых не велик, и поэтому делает возможным развитие прикладного аспекта типологии.
Список литературы
[Alekhin] Алехин В.В. 1951. Растительность СССР в основных зонах. М. 512 с.
[Aleksandrova] Александрова В.Д. 1969. Классификация растительности. Обзор принципов классификации и классификационных систем в разных геоботанических школах. Л. 275 с.
[Alekseev, Molchanov] Алексеев С.В., Молчанов А.А. 1954. Выборочные рубки в лесах Севера. М. 146 с.
[Bobkova et al.] Бобкова К.С., Галенко Э.П., Загирова С.В., Сенькина С.Н., Тужилкина В.В., Машика А.В., Патов А.И., Никонов В.В., Лукина Н.В., Исаева Л.Г. 2006. Коренные еловые леса севера: биоразнообразие, структура, функции. СПб. 337 с.
Cajander A.K. 1926. The theory of forest types. – Acta Forestalia Fennica. 29: 1–108.
[Chertovskoj] Чертовской В.Г. 1978. Еловые леса европейской части СССР. М. 176 с.
[Dolgin et al.] Долгин М.М., Колесникова А.А., Конакова Т.Н. 2012. Динамика численности почвенной мезофауны в среднетаежных лесах республики Коми. – Вестник САФУ. Серия: Естественные науки. 4: 61–68.
[Dyrenkov] Дыренков С.Н. 1984. Структура и динамика таежных ельников. Л. 174 с.
[Fedorets] Федорец Н.Г., Морозова Р.М., Синькевич С.М., Загуральская Л.М. 2000. Оценка продуктивности лесных почв в Карелии. Петрозаводск. 195 с.
[Genikova et al.] Геникова Н.В., Гнатюк Е.П., Крышень А.М. 2012. Анализ ценофлоры лесов на автоморфных песчаных почвах в Карелии. – Бот. журн. 97 (11): 1424–1435.
[Genikova et al.] Геникова Н.В., Гнатюк Е.П., Крышень А.М. 2019. Ценофлора ельников черничных Восточной Фенноскандии. – Бот. журн. 104 (5): 699–716. https://doi.org/10.1134/S0006813619050041
Hill M.O. 1979. DECORANA – a FORTRAN programm for detrended correspondence analysis and reciprocal averaging. N.Y. 31 p.
[Kachinskiy] Качинский Н.А. 1965. Физика почв. М. 324 с.
[Kazimirov] Казимиров Н.И. 1971. Ельники Карелии. Л. 140 с.
Kreutz A., Aakala T., Grenfell R., Kuuluvainen T. 2015. Spatial tree community structure in three stands across a forest succession gradient in northern boreal Fenno-scandia. Silva Fennica. 49 (2). https://doi.org/10.14214/sf.1279
[Kryshen] Крышень А.М. 2003. Структура и динамика растительного сообщества вейниковой вырубки. Ч. 1. Видовой состав. – Бот. журн. 88 (4): 48–62.
[Kryshen] Крышень А.М. 2006. Растительные сообщества вырубок Карелии. М. 262 с.
[Kryshen] Крышень А.М. 2010. Типы лесорастительных условий на автоморфных почвах в Карелии. – Бот. журн. 95 (3): 281–297.
[Kryshen et al.] Крышень А.М., Геникова Н.В., Гнатюк Е.П., Преснухин Ю.В., Ткаченко Ю.Н. 2018. Ряды восстановления сосняков восточной Фенноскандии на песчаных автоморфных почвах. – Бот. журн. 103 (1): 5–35. https://doi.org/10.1134/S0006813618010015
[Kryshen et al.] Крышень А.М., Гнатюк Е.П., Геникова Н.В., Рыжкова Н.И. 2016. Сравнительный анализ эколого-ценотических групп в структуре парциальных флор антропогенно фрагментированной территории. – Бот. журн. 101 (5): 489–516.
Kuuluvainen, T., Aakala, T. 2011. Natural forest dynamics in boreal Fennoscandia: a review and classification. – Silva Fennica. 45 (5): 823–841.
Kuuluvainen T., Wallenius T.H., Kauhanen H., Aakala T., Mikkola K., Demidova N., et Ogibin, B. 2014. Episodic, patchy disturbances characterize an old-growth Picea abies dominated forest landscape in northeastern Europe. – For. Ecol. Manag. 320: 96–103. https://doi.org/10.1016/j.foreco.2014.02.024
[L’vov, Ipatov] Львов П.Н., Ипатов Л.Ф. 1976. Лесная типология на географической основе. Архангельск. 195 с.
[Likhanova et al.] Лиханова И.А., Генрих Э.А., Перминова Е.М. 2018. Динамика растительности после сплошнолесосечных вырубок среднетаежных ельников черничных Северо-Востока европейской части России. – В сб.: Материалы ХVI Всерос. конф. “Биодиагностика состояния природных и природно-техногенных систем”. Киров. С. 77–81.
[Lukina et al.] Лукина Н.В., Исаев А.С., Крышень А.М., Онучин А.А., Сирин А.А., Гагарин Ю.Н., Барталев С.А. 2015. Приоритетные направления развития лесной науки как основы устойчивого управления лесами. – Лесоведение. 4: 243–254.
[Manov] Манов А.В. 2017. Динамика горизонтальной структуры древостоя коренного разнотравно-черничного ельника подзоны средней тайги республики Коми. – Известия РАН. Серия биологическая. 5: 573–580.
[Manov] Манов А.В. 2019. Горизонтальная структура древостоя и подроста ельника разнотравно-черничного средней тайги республики Коми. – Лесоведение. 4: 286–293. https://doi.org/10.1134/S024114819030069
[Morozova] Морозова Р.М. 1991. Лесные почвы Карелии. Л. 184 с.
[Osipov et al.] Осипов А.Ф., Тужилкина В.В., Дымов А.А., Бобкова К.С. 2019. Запасы фитомассы и органического углерода среднетаежных ельников при восстановлении после сплошнолесосечной рубки. – Известия РАН. Серия биологическая. 2: 215–224.
[Pristova] Пристова Т.А. 2008. Биологический круговорот веществ во вторичном лиственно-хвойном насаждении средней тайги. – Экология. 3: 189–195.
[Rysin, Savel’eva] Рысин Л.П., Савельева Л.И. 2002. Еловые леса России. М. 335 с.
Shorohova E., Kuuluvainen T., Kangur A., Jogiste K. 2009. Natural stand structures, disturbance regimes and successional dynamics in the Eurasian boreal forests: a review with special reference to Russian studies. – Ann. For. Sci. 66 (2): 1–20. htps://doi.org/https://doi.org/10.1051/forest/2008083
[Sukachev] Сукачев В.Н. 1972. Избранные труды. Т. 1. Л. 418 с.
[Vasilevich] Василевич В.И. 1983. О растительных ассоциациях ельников Северо-Запада. – Бот. журн. 68 (12): 1604–1613.
[Vasilevich] Василевич В.И. 2004. Ельники черничные Европейской России. – Бот. журн. 89 (11): 1728–1739.
[Volkov] Волков А.Д. 2003. Биоэкологические основы эксплуатации ельников северо-запада таежной зоны России. Петрозаводск. 246 с.
[Yakovlev, Voronova] Яковлев Ф.С., Воронова В.С. 1959. Типы лесов Карелии и их природное районирование. Петрозаводск. 190 с.
[Yurkovskaya] Юрковская Т.К. 1993. Растительный покров Карелии – В кн.: Растительный мир Карелии и проблемы его охраны. Петрозаводск. С. 8–36.
[Yurkovskaya, Elina] Юрковская Т.К., Елина Г.А. 2009. Восстановленная растительность Карелии на геоботанической и палеокартах. Петрозаводск. 136 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал