

Ботанический журнал, 2021, T. 106, № 3, стр. 272-291
МОРФОЛОГИЯ ПЫЛЬЦЫ ТРИБЫ ANDROSACEAE (PRIMULACEAE)
Д. А. Брицкий 1, *, В. В. Григорьева 1, **
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
* E-mail: dmibri@mail.ru
** E-mail: mikhailov_val@mail.ru
Поступила в редакцию 25.08.2020
После доработки 22.09.2020
Принята к публикации 29.09.2020
Аннотация
С помощью светового (СМ) и сканирующего электронного (СЭМ) микроскопов изучена морфология пыльцевых зерен 26 видов рода Androsace, 7 видов Douglasia, 1 вида Vitaliana, 1 вида Pomatosace и 6 видов Primula. Пыльцевые зерна всех изученных видов 3-бороздно-оровые, эллипсоидальные или вытянутые. Выявлено 3 типа эндоапертур: округлые, эллиптические и экваториально вытянутые, образующие экваториальный поясок. Скульптура перфорированная, микросетчатая, складчато-перфорированная, мелкобугорчатая. Границы между секциями рода Androsace по морфологическим признакам пыльцы в большинстве случаев установить не удается. Исключение составляет секция Megista, пыльцевые зерна видов из этой секции обладают уникальной для пыльцы Androsace мелкобугорчатой скульптурой. Форма и очертания пыльцевых зерен, особенности строения эндоапертур, скульптура экзины позволяют различать отдельные виды внутри секций.
Триба Androsaceae Rchb. ex Pax. (сем. Primulaceae Vent.) объединяет роды Androsace L., Douglasia Lindl., Pomatosace Maxim. и Vitaliana Sesl. Наиболее многочисленным в трибе является род Androsace (проломник), в пределах которого насчитывается около 120 видов, распространенных преимущественно в горных областях Евразии, с наибольшим обилием в горах Малой Азии и Китая (Wendelbo, 1961; Hu, 1994; Hu, Kelso, 1996; Mabberley, 2009). Виды Androsace – однолетние, двулетние или многолетние травы с восходящими или стелющимися побегами, образующими рыхлые или плотные подушкообразные дерновины. По числу входящих в него видов этот род занимает третье место в семействе Primulaceae. Род Androsace разделен на несколько секций, критериями для выделения которых стали такие признаки, как жизненная форма, строение цветка, способ раскрывания коробочек и хромосомные числа (Wendelbo,1961; Hu, Yang, 1986). Во Флоре СССР описано 33 вида Androsace из 5 секций (Chamaejasme, Pseudodouglasia, Samuelia, Andraspis, Megista) (Shishkin, Bobrov, 1952).
Род Douglasia объединяет 9–11 видов, которые встречаются в Северной Америке, на Аляске и Дальнем Востоке России, чаще всего на прибрежных скалах и в высокогорьях. Основное отличие представителей рода от Androsace: большее число хромосом и неравномерно утолщенные клеточные стенки эндосперма (Kelso, 1992; Anderberg and Kelso, 1996).
Монотипный род Vitaliana встречается в горах Италии, Франции, Швейцарии, Австрии, с конца XIX века широко культивируется при создании альпийских садов. V. primuliflora Bertol. отличается от всех других представителей трибы Androsaceae наличием гетеростилии, что и стало основанием для выделения этого вида в самостоятельный род (Ferguson, 1972; Smith, Lowe, 1997). Некоторые авторы включают V. primuliflora в род Androsace в ранге секции (Wendelbo, 1961; Kress, 1965, 1967). Современные молекулярно-генетические исследования также свидетельствуют о том, что этот вид должен располагаться среди Androsace (Trift et al., 2002; Martins et al., 2003; Dixon et al., 2016).
Pomatosace filicula Maxim. – единственный представитель рода Pomatosace, встречается только в северо-восточной части Цинхай-Тибетского плато, в китайских провинциях Сычуань, Сицзан (Тибет) и Цинхай, растет в различных местах обитания, включая альпийские луга и песчаные равнины вдоль рек, на высотах 2800–4500 м. Отличается от представителей Androsace морфологическими особенностями семян (Hu, Kelso, 1996).
Проломники обычно тесно связывают с такими родами, как Douglasia, Vitaliana, Pomatosace, статус которых до сих пор остается предметом дискуссии (Martins et al., 2003; de Vos et al., 2014 и др.).
Молекулярно-филогенетические данные показали, что роды Androsace и Primula L. рассматриваются как две основные клады семейства Primulaceae, от которых, вероятно, произошли еще несколько небольших родственных родов (Martins et al., 2003; de Vos et al., 2014 и др.).
Род Primula – самый большой и полиморфный род в семействе Primulaceae, объединяет около 500 видов травянистых растений, произрастающих в холодном, умеренном и субтропическом поясах Северного полушария. Центрами видового разнообразия считаются Гималаи, горы Юго-Западного Китая, Кавказ и Альпы (Mast et al., 2001). Еще в первой монографии, посвященной роду Primula (Pax, 1889) впервые было высказано предположение о близком родстве между родами Androsace и Primula и отсутствии между ними четких границ. Основанием для такого предположения послужил тот факт, что ряд видов Primula (P. forbesii Franch., P. malacoides Franch., P. malvaceae Franch. и др.), произрастающих в первичных центрах разнообразия этих родов (Гималаях и горах Юго-Западного Китая) обладают признаками, свойственными как Primula, так и Androsace и, по мнению F. Pax (1889), могут быть отнесены как к тому, так и к другому роду.
P. Wendelbo (1961), используя световую микроскопию (СМ), описал пыльцевые зерна 48 видов Androsace из 7 секций. На основании полученных данных этот автор сделал вывод об уникальности пыльцы родов Androsace, Douglasia, Vitaliana и Pomatosace и выделил в семействе Primulaceae отдельный палиноморфологический “Androsace-тип”. Кроме того, Wendelbo предположил, что размер пыльцы может быть использован для разграничения секций в пределах рода Androsace. Y.J. Nasir (1986), изучив морфологию пыльцы 22 видов Androsace, произрастающих в Пакистане, отметил, что ширину пыльцевого зерна и длину борозд можно считать таксономически значимыми признаками секционного уровня в этом роде. Масштабное палинологическое исследование 78 видов из рода Androsace, произрастающих главным образом в Китае, а также связанных с ним родов Douglasia, Vitaliana и Pomatosace значительно расширило представления о палиноморфологии этих родов (Xu et al., 2016) и подтвердило вывод о монофилетичности группы Androsace, сделанный рядом авторов в ходе молекулярно-филогенетических исследованиий (Mast et al., 2001; Trift et al., 2002; Martins et al., 2003; de Vos et al., 2014).
Сведения о пыльце отдельных видов имеются также в работах, посвященных систематике рода Androsace (Kim, Kim, 1988) и палиноморфологии семейства Primulaceae (Spanowsky, 1962; Punt et al., 1974; Kupriyanova, Aleshina, 1978 и др.).
Несмотря на довольно большой объем информации о морфологии пыльцы Androsace, остаются неясными некоторые вопросы их палиноморфологии. В частности нет сведений о строении эндоапертур, при том что этот признак часто играет важную роль для уточнения вопросов систематики и филогении. Кроме того, до сих пор остается открытым вопрос о границах между родами Androsace и Primula, в связи с чем дополнительно была изучена пыльца некоторых видов рода Primula, обладающих значительным морфологическим (и прежде всего палиноморфологическим) сходством с представителями Androsace.
Цель данной работы – оценка таксономической значимости особенностей морфологии пыльцы родов Androsace, Douglasia, Vitaliana, Pomatosace и видов Primula с 3-бороздно-оровым типом пыльцы.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для исследования послужили зрелые пыльцевые зерна видов Androsace, Douglasia, Vitaliana, Pomatosace, Primula (Primulaceae), взятые с гербарных образцов, хранящихся в Гербарии Ботанического института им. В.Л. Комарова (LE).
Для светооптического исследования применяли классический ацетолизный метод Эрдтмана (Erdtman, 1952). Световые микрофотографии получены на микроскопе Carl Zeiss Jenaval, с помощью цифровой фотокамеры Canon EOS 20D.
Детали строения поверхности пыльцевых зерен уточняли на сканирующем электронном микроскопе JEOL JSM-6390 в центре коллективного пользования Ботанического института им. В.Л. Комарова. Для исследования необработанную пыльцу извлекали из пыльников и закрепляли на специальном столике с помощью двусторонней липкой ленты, затем напыляли сплав золота и палладия в вакуумной установке. Из-за мелких размеров пыльцы исследованных видов при изучении поверхности пыльцевых зерен с помощью СЭМ использовали большее увеличение по сравнению со стандартным, общепринятым в палинологии.
При изучении морфологии пыльцевых зерен учитывали: тип и число апертур, форму и очертания пыльцевых зерен, размеры полярной оси и экваториального диаметра, особенности строения борозд и межапертурных участков, толщину экзины и характер скульптуры (Kupriyanova, Aleshina, 1967, 1972). Измерения проводили с помощью окулярмикрометра, в каждом образце измеряли не менее 15 пыльцевых зерен.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Основные морфологические характеристики пыльцы изученных видов представлены в табл. 1 “Основные характеристики пыльцы исследованных видов Androsace, Douglasia, Vitaliana, Pomatosace, Primula”.
Таблица 1.
Основные характеристики пыльцы исследованных видов Androsace, Douglasia, Vitaliana, Pomatosace, Primula Table 1. Main characteristics of pollen of the studied species of Androsace, Douglasia, Vitaliana, Pomatosace, Primula
| Исследован-
ный образец Studied sample |
Очертание
с экватора Equatorial view |
Очертание
с полюса Polar view |
Длина полярной
оси (п.о.) (мкм) Polar axis length (P) (μm) |
Длина эквато-
риального диаметра (э.д.) (мкм) Equatorial axis length (E) (μm) |
Соотно-
шение P/E P/E ratio |
Эндоапертура Endoaperture |
Толщина
экзины (мкм) Exine thick- ness (μm) |
Скульптура Exine orna- mentation |
|---|---|---|---|---|---|---|---|---|
| Genus Androsace L./Sect. Samuelia Schlechtd. | ||||||||
| A. gmelini Guert. 08.08.1970. В. Грубов/V. Grubov | Эллиптическое или почти округлое/Elliptic to almost circular | 3-лопастное 3-lobate |
12.4–15.6 | 12.0–13.0 | 1.0–1.3 | Нечеткая, вытянута по п.о. Fuzzy outline, extended along the P axis 2.2–2.4 × 2.4–2.6 μm | 0.7–0.8 | Перфорированная Perforate |
| A. henryi Oliver China. 22.08.1904. № 4033. E.H. Wilson | Округло-
прямоугольное Slightly rectangular |
3-лопастное 3-lobate |
12.4–13.2 | 9.2–9.6 | 1.3–1.4 | Нечеткая, округлая или вытянута по п.о. Fuzzy outline, circular or extended along the P axis 1.6–1.9 × 2.0–2.3 μm | 0.8–0.9 | Перфорированная Perforate |
| A. rotundifolia Sm. Китай, Сычуань/China, Sichuan. 26.03.1893. G.N. Potanin | Вытянутое, слегка сжатое с боков Lalongate |
3-лопастное 3-lobate |
11.7–12.4 | 7.2–7.6 | 1.6 | Округлые или вытянуты по п.о./Fuzzy outline, circular or extended along the P axis 2.0–2.2 × 1.9–2.2 μm | 0.4–0.6 | Перфорированная Perforate |
| A. umbellata Merril Rynkyn. Pref. Okinawa. 06.03.1975. M. Togashi | Эллиптическое Elliptic |
3-лопастное 3-lobate |
11.3–12.1 | 7.9–8.5 | 1.4 | Неясная в очертании Fuzzy outline |
0.8–0.9 | Перфорированная Perforate |
| Sect. Aizoidium Hand.-Mazz. | ||||||||
| A. aizoon Duby Yunnan. 1887. Delavay | Округло-
прямоугольное Rectangular |
3-лопастное 3-lobate |
17.3–18.6 | 9.9–10.7 | 1.7 | Экваториальный пояс, 2.0–2.2 мкм шириной Endocingula, 2.0–2.2 μm wide | 1.4–1.5 | Перфорированная Perforate |
| A. coccinea Franch. Yunnan. 1886. Delavay | Округло-прямоугольное слегка сжатое
по экватору Rectangular |
Округло-
треугольное Triangular |
19.5–20.7 | 10.5–11.8 | 1.8–1.9 | Экваториальный пояс Endocingula |
0.8–0.9 | Микро-
сетчатая Micro- reticulate |
| Sect. Chamaejasme C. Koch | ||||||||
| A. acbaitalensis Derg. Памир. Чечекты/Pamir. Chechekty. 05.08.1960. Э.О. Белянская, С.С. Иконников/E.O. Belyanskaya, S.S. Ikonnikov | Эллиптическое Elliptic |
3-лопастное 3-lobate |
12.7–13.5 | 9.2–10.4 | 1.3–1.4 | Эллиптические/Elliptic 2.2–2.6 × 3.7–4.5 μm | 1.1–1.2 | Микро-
сетчатая Micro- reticulate |
| A. alaschanica Maxim. Ю. Монголия. Алашан/S. Mongolia. Alashan. 27.04.1908. С.С. Четырин/S.S. Chetyrin | Вытянутое Lalongate |
Округло-треугольное Triangular |
17.5–18.6 | 9.9–11.1 | 1.7–1.8 | Экваториально вытянуты, иногда образуют пояс Lalongate 1.8–2.1 × 3.0–3.7 μm |
0.8–0.9 | Микро-
сетчатая Micro- reticulate |
| A. alascana Coville et Standley ex Hulten Alaska, 4.07.1951. J.A. Calder. № 5626 | Округло-
прямоугольное Rectangular |
3-лопастное 3-lobate |
20.7–21.9 | 11.8–12.6 | 1.7–1.75 | Эллиптические, вытянуты по э.д./Lalongate 2.3–2.8 × 4.2–4.7 μm | 1.1–1.2 | Микро-
сетчатая Micro- reticulate |
| A. bungeana Schischk. et Bobrov Анадырский р-н/Anadyr Distr. 11.06.1951. Б. Юрцев\B. Yurtsev |
Округло-
прямоугольное Rectangular |
Треугольное Triangular |
19.8–21.1 | 14.1–14.9 | 1.4 | Экваториальный пояс
с нечеткими краями,
1.7–2. 2 мкм шириной Endocingula with fuzzy margins, 1.7–2.2 μm wide |
0.7–0.8 | Перфорированная Perforate |
| A. capitata Willd. ex Roem. et Schult. Чукотский нац. округ/ Chukotka National District. 08.07.1950. М.Н. Аврамчик/ M.N. Avramchik | Эллиптическое Elliptic |
Округло-треугольное Triangular |
13.1–14.1 | 9.1–9.9 | 1.4 | Эллиптические по э.д. Elliptic 1.5–1.8 × 2.9–3.3 μm |
0.8–0.9 | Микро-
сетчатая Microreticulate |
| A. incana Lam. МНР/Mongolian People’s Republic. 05.06.1975. Чепурнов/Chepurnov | Округло-
прямоугольное Rectangular |
3-лопастное 3-lobate |
13.0–13.9 | 8.8–9.9 | 1.4–1.5 | Эллиптические по э.д. или пояс Lalongate or endocingula | 0.8–0.9 | Микро-
сетчатая Microreticulate |
| A. chamaejasme Wulfen ex Host Чукотский п-ов, бассейн реки Адзьвы/Chukchi Peninsula. Adzva River basin. 12.07.1959. В.М. Герасимова/V.M. Gerasimova |
Округло-
прямоугольное Rectangular |
Округло-треугольное Triangular |
16.9–17.9 | 10.3–11.5 | 1.5–1.6 | Эллиптические
вытянуты по э.д.
Lalongate 2.4–2.6 × 2.4–2.8 μm |
0.7–0.8 | Складчато-
перфорированная Rugulate- perforate |
| A. spinulifera Knuth Tibet oriental. Tongolo. 07.1894. № 2929. J.-A. Soulie | Округло-прямо-
угольное, слегка
сжатое по экватору Rectangular |
Округло-треугольное Triangular |
16.6–17.8 | 9.2–10.4 | 1.7–1.8 | Экватор пояс 2.0–2.3 мкм шириной или экваториально вытянутые Lalongate or endocingula 2.0–2.3 μm wide | 0.9–1.0 | Микро-
сетчатая Micro- reticulate |
| A.strigillosa Franch. Сычуань/ Sichuan. 1938. № Ас225. McLaren | Округло-
прямоугольное Rectangular |
Округло-треугольное Triangular |
18.6–19.8 | 8.9–9.5 | 2.1 | Экваториальный пояс
2.4–2.9 мкм шириной Endocingula 2.4–2.9 μm wide |
0.6–0.7 | Микро-
сетчатая Microreticulate |
| A. triflora Adams. Таймырский п-ов/ Taimyr Peninsula. 03.08.1949. И. Серебряков, П. Кузнецов/I. Se-rebryakov, P. Kyznetsov | Округло-прямоугольное, слегка сжатое
по экватору Rectangular |
Округло-треугольное Triangular |
13.6–14.4 | 8.5–9.7 | 1.5–1.6 | Экваториальный пояс
2.2–2.6 мкм шириной Endocingula 2.2–2.6 μm wide |
0.7–0.8 | Складчато-
перфорированная Rugulate- perforate |
| A. villosa L. Кирг. ССР. Джеты-Огузовский р-н/Kirghiz SSR. Jeti-Ögüz Distr. 20.07.1935. Савич, Сабардина/Savich, Sabardina | Округло-прямоугольное, реже почти эллиптическое Rectangular or elliptic |
Округло-треугольное Triangular |
13.4–14.6 | 8.7–9.3 | 1.5–1.6 | Нечеткий экваториальный пояс 2.2–2.6 мкм шириной Endocingula 2.2–2.6 μm wide |
0.9–1.0 | Перфорированная или складчато-
перфорированная Perforat or rugate- perforate |
| Sect. Orthocaulon Hand.-Mazz. | ||||||||
| A. erecta Maxim. 14–26.07.1880 | Округло-
прямоугольное Rectangular |
слабо-3-лопастное или почти округлое Slightly 3-lobate or almost circular |
14.1–15.3 | 9.5–10.3 | 1.5 | Эллиптические по э.д. Elliptic 2.8–3.3 × 4.1–4.6 μm |
0.7–0.8 | микро-
сетчатая Micro- reticulate |
| Sect. Andraspis (Duby) C. Koch | ||||||||
| A. albana Stev. Кавказ, верховья Кубани/Caucasus, upper reaches of the Kuban. 20.05.1900. Десулави/Desoulavi | Эллиптическое Elliptic |
3-лопастное 3-lobate |
15.8–16.9 | 11.4–12.6 | 1.3–1.5 | Округлые
Circular 2.3–2.5 μm |
0.9 | Микро-
сетчатая Micro- reticulate |
| A. elongata L. Самарская губ. Новоуездный у./Samara Govern. Novouyezdnyi Uyezd/01.05.1900. В. Богдан/V. Bogdan | Эллиптическое Elliptic |
Почти округлое Almost circular |
14.9–16.2 | 10.9–12.1 | 1.3–1.4 | Округлые
Circular 4.1–4.6 μm |
0.9 | Микро-
сетчатая Micro- reticulate |
| A. fedtschenkoi Ovcz. Киргизская ССР. Кочкорский р-н. Верховья р. Кара-Кобта/Kirghiz SSR. Kochkor Distr. Upper reaches of the Kara-Kobta River. 28.06. 1936. Блументаль, Овчинников/Blumental, Ovchinnikov | Эллиптическое или почти округлое Elliptic or almost circular |
3-лопастное 3-lobate |
21.7–23.1 | 18.0–19.1 | 1.1–1.2 | Эллиптические
Elliptic 1.9–2.2 × 2.9–3.5 μm |
1.1–1.2 | Микро-
сетчатая Micro- reticulate |
| A. filiformis Retz. Приморский край. Ханкайский р-н/Primorye Territory. Khanka Distr. 05.06.1955. Г.Ф. Патреевская/G.F. Patreyevskaya | Округлое или
эллиптическое Circular or elliptic |
Округлое Circular |
10.3–11.3 | 10.1–11.3 | 1–1.2 | Эллиптические
Elliptic 1.7–2.1 × 2.6–3.1 μm |
Перфорированная Perforate |
|
| A. lactiflora Franch.ex Willd. 18.06.1971. № 98. В.И. Грубов/ V.I. Grubov | Округло-
прямоугольное Rectangular |
Округло-треугольное Triangular |
14.7–15.9 | 8.1–9.3 | 1.7–1.8 | Округлые
Circular 2.3–2.7 × 2.7–3.2 μm |
0.7–0.8 | Перфорированная Perforate |
| A. septentrionalis L. Омск/Omsk. 15.05.1920. Г.И. Городков/ G.I. Gorodkov | Эллиптическое
или округло-
прямоугольное Elliptic or rectangular |
Округлое или округло-треугольное Circular or triangular |
13.3–14.5 | 8.7–9.4 | 1.5 | Вытянуты по э.д.,
часто образуют пояс
Endocingula 1.5–2.1 × 3.1–3.4 μm |
0.6–0.7 | Морщинисто-перфорированная Rugate- perforate |
| Sect. Megista Schlechtd. | ||||||||
| A. maxima L. Центральный Саян. Бадаиха/Central Sayan. Badaikha. 24.05.1948. А. Федоров и др./ A. Fedorov et al. | Эллиптическое Elliptic |
Округло-треугольное или почти округлое Triangular or almost circular |
18.4–19.6 | 14.9–15.9 | 1.2 | Эллиптические
Elliptic 3.2–3.6 × 5.9–6.7 μm |
0.8–0.9 | На полюсе перфорированная, на экваторе мелкобугорчатая/Perfo-rate at apocolpium and microverrucate at mesocolpium |
| A. turczaninowii Freyn. Тувинская АССР/Tuva ASSR. 29.05.1972. В. Ханминчун/V. Khanminchun | Эллиптическое Elliptic |
Слабо 3-лопастное или почти округлое Slightly 3-lobate or almost circular |
14.9–16.0 | 12.1–12.8 | 1.2–1.25 | Эллиптические Elliptic 2.6–2.9×3.2–3.7 μm |
0.8–0.9 | Мелко-бугорчатая Microver-rucate |
| Genus Douglasia Lindl. | ||||||||
| D. arctica Hook. Alaska. 28.07.1964. Fran Chauvin | Эллиптическое или прямоугольное Elliptic or rectangular |
3-лопастное 3-lobate |
18.3–20.1 | 9.8–10.3 | 1.9–1.95 | Округлые или
вытянутые по э.д. Circular or lalongate 3.3–3.9×2.8–3.9 μm |
1.0 | Складчато-перфорированная Rugate- perforate |
| D. biflora A. Nelson Yellowstone National park. 29.07.1899. № 6203. Aven Nelson, Elias Nelson | Эллиптическое
или округло-
прямоугольное Elliptic or rectangular |
3-лопастное 3-lobate |
20.1–21.3 | 10.5–11.2 | 1.9 | Экваториально вытянутые Lalongate 2.8–3.1 × 4.2–4.8 μm |
0.8 | Сладчато-перфорированная Rugate- perforate |
| D. gormanii Constance Canada, Yukon. 15.07.1971. № 11123. S.L. Welsh, J.K. Rigby | Эллиптическое
или округло-
прямоугольное Elliptic or rectangular |
Округло-треугольное или 3-
лопастное Triangular or 3-lobate |
19.7–20.9 | 10.5–11.6 | 1.8–1.9 | Эллиптические по э.д. Lalongate 2.4–3.2 × 3.6–4.0 μm |
0.9–1.0 | Сладчато-перфорированная Rugate- perforate |
| D. laevigata A. Gray Washington. 31.05.1936. W.J. Egerdan | Округло-
прямоугольное Rectangular |
3-лопастное 3-lobate |
24.1–25.6 | 12.9–13.5 | 1.8–1.9 | Вытянуты по э.д. Lalongate 2.8–3.3 × 3.3–4.0 μm |
0.9–1.0 | Сладчато-перфорированная Rugate- perforate |
| D. montana A. Gray Idaho. № 13766. Dr. Lincoln Constance | Округло-
прямоугольное Rectangular |
3-лопастное 3-lobate |
20.5–21.7 | 11.4–12.2 | 1.8 | Округлые или слегка
вытянуты по э.д. Circular or lalongate 3.5–3.9 × 3.8–4.1 μm |
0.9–1.0 | Сладчато-перфорированная Rugate- perforate |
| D. nivalis Lindl. Washington. 4.07.1933. W.J. Egerdam | Округло-
прямоугольное Rectangular |
3-лопастное 3-lobate |
19.7–20.9 | 11.1–11.7 | 1.7–1.8 | Округлые или слегка
вытянутые по э.д. Circular or lalongate 3.3–3.6 × 3.7–4.1 μm |
0.9–1.0 | Сладчато-перфорированная Rugate- perforate |
| D. ochotensis (Willd. ex Roem. et Schult.) Hult. Якутская АССР/Yakut ASSR. 02.07.1974. А.А. Коробков, В.В. Культина, М.А. Михайлова/A.A. Korobkov, V.V. Kultina, M.A. Mikhailova | Эллиптическое
или округло-
прямоугольное Elliptic or rectangular |
Округло-треугольное Triangular |
18.4–19.5 | 11.3–11.8 | 1.6 | Эллиптические
Elliptic 2.3–2.6 × 3.4–3.7 μm |
0.9–1.0 | Сладчато-перфорированная Rugate- perforate |
| Genus Vitaliana Sesl. | ||||||||
| V. primuliflora Bertol. СПб. Ботанический сад БИН РАН/St. Petersburg. Botanical Garden of the Komarov Boranical Institute. 13.05.1998. Д.А. Брицкий/D.A. Britskii | Округло-
прямоугольное Rectangular |
Округло-треугольное Triangular |
26.3–27.8 | 14.4–15.5 | 1.8 | Округлые или слегка
вытянуты по э.д. Circular or lalongate 4.5–4.9 × 4.8–5.1 μm |
1.1–1.2 | Перфорированная Perforate |
| Genus Pomatosace Maxim. | ||||||||
| P. filicula Maxim. Тибет. Озеро Русское/Tibet. Russkoye Lake. 20.06.1900. В.Ф. Ладыгин/ V.F. Ladygin | Округло-
прямоугольное Rectangular |
3-лопастное 3-lobate |
17.9–18.9 | 10.7–11.1 | 1.7 | Эллиптические, по э.д. Elliptic 4.1–4.4 × 5.6–5.9 μm |
0.9–1.1 | Микро-
сетчатая Micro- reticulate |
| Genus Primula L./Sect. Aleurita Daby | ||||||||
| P. darialica Rupr., короткостолбиковая форма/short-styled form/Гербарий Срединского/ Sredinsky’s herbarium | Эллиптическое Elliptic |
Округло-треугольное Triangular |
12.8–14.2 | 12.1–12.8 | 1–1.1 | Элиптические по э.д. Elliptic 3.8–4.2 × 4.0–4.6 μm |
1.1–1.3 | Микро-
сетчата Micro- reticulate |
| P. farinifolia Rupr., короткостолбиковая форма/short-styled form./Грузинская ССР/Georgian SSR. 08.04.1966. Е.В. Мордак/E.V. Mordak | Широкоэллип-
тическое Broadly elliptic |
Округло-треугольное Triangular |
11.4–12.3 | 11.1–11.9 | 1 | Неясные Fuzzy outline |
1.1–1.2 | Микро-
сетчатая Micro- reticulate |
| P. longiscapa Ledeb., длинностолбиковая форма/long-styled form Тургайская обл./Turgay Region 08.07.1908. М. Крашенинников/ M. Krasheninnikov |
Эллиптическое Elliptic |
3-лопастное 3-lobate |
16.1–17.1 | 8.5–9.2 | 1.8–1.9 | Округлые или слегка
вытянутые в полярном направлении
Circular 1.9–2.2 × 1.5–1.8 μm |
1.1–1.2 | Микро-
сетчатая Micro- reticulate |
| P. longiscapa Ledeb., короткостолбиковая форма/short-styled form/Тургайская обл./Turgay Region 08.07.1908. М. Крашенинников/ M. Krasheninnikov | Эллиптическое Elliptic |
3-лопастное 3-lobate |
10.9–12.1 | 10.1–11.5 | 1 | Fuzzy outline | 1.1–1.2 | Микро-
сетчатая Micro- reticulate |
| P. zeylamica Charadze et Kapell. Бассейн реки Ассы/Assa River basin. H 2300 m. 24.07.1948. А. Харадзе, Д. Очиаури, Л. Хинтибидзе/A. Kharadze, D. Ochiauri, L. Khintibidze |
Эллиптическое Elliptic |
Округло-треугольное Triangular |
12.4–13.5 | 12.5–13.7 | 0.9 | Эллиптическая
Elliptic 3.6–4.2 × 4.4–4.7 μm |
1.2–1.3 | Микро-
сетчатая Micro- reticulate |
| Sect. Malacoides Balf. | ||||||||
| P. forbesii Franch., длинностолбиковая форма/long-styled form Yunnan. № 9853. A. Henry |
Эллиптическое Elliptic |
3-лопастное 3-lobate |
11.7–12.5 | 8.5–9.3 | 1.3–1.4 | Округлые или слегка
вытянуты по п.о.
Circular 2.2–2.6 × 2.1–2.4 μm |
1.1–1.3 | Микро-
сетчатая Micro- reticulate |
| P. forbesii Franch., короткостолбиковая форма/short-styled form/Yunnan. 02.1887. № 311. Delavay | Эллиптическое Elliptic |
3-лопастное 3-lobate |
13.4–14.8 | 9.1–9.7 | 1.5 | Округлые или слегка
вытянуты по п.о.
Circular 2.1–2.4 × 2.0–2.3 μm |
1.1–1.3 | Микро-
сетчатая Micro- reticulate |
| P. malacoides Franch., короткостолбиковая форма/short-styled form/Yunnan. 20.02.1891. № 6770. Delavay | Эллиптическое Elliptic |
3-лопастное 3-lobate |
28.4–30.2 | 23.5–25.5 | 1.2 | Слегка вытянуты по э.д. Elliptic 1.1–1.3 × 1.6–2.0 μm |
1.0 | Микро-
сетчатая Micro- reticulate |
Род Androsace
Пыльцевые зерна радиально-симметричные, изополярные, 3-бороздно-оровые, мелкие (табл. I ). Длина полярной оси не превышают 25 мкм. Самые мелкие зерна выявлены у A. filiformis (полярная ось 10.3–11.3 мкм, экваториальный диаметр 10.1–11.3 мкм). Следует отметить, что у пыльцы некоторых видов, при несколько больших значениях длины полярной оси, экваториальный диаметр не достигает 10 мкм. Например, у пыльцевых зерен A. rotundifolia полярная ось 11.7–12.4 мкм, тогда как экваториальный диаметр всего 7.2–7.6 мкм. Самые крупные зерна обнаружены у A. fedtschenkoi (полярная ось 21.7–23.1 мкм, экваториальный диаметр 18.0–19.1 мкм). У пыльцы большинства исследованных видов длина полярной оси колеблется от 0.7 до 1.3 мкм. Только у пыльцы A. fedtschenkoi эта разница достигает 2 мкм. Значения длины экваториального диаметра изменяются от 0.3 мкм у пыльцы A. coccinea до 1.4 мкм у пыльцы A. gmelini.
Таблица I . Морфология пыльцы представителей рода Androsace (СМ)
1–6 – секция Samuelia; 1–2 – A. gmelini; 3–4 – A. henryi; 5–6 – A. umbellata; 7–9 – секция Aizoidium; 7–8 – A. aizoon; 9 – A. coccinea; 10–21 – секция Chamaejasme; 10 – A. alascana; 11 – A. bungeana; 12–14 – A. incana; 15–18 – A. strigillosa; 19–21 – A. villosa; 22–24 – секция Orthocaulon, A. erecta; 25–30 – секция Andraspis; 25–28 – A. albana; 29–30 – A. elongata; 31–35 – секция Megista, A. turczaninowii; 36 – Douglasia ochotensis. 1, 2, 4, 6, 7, 9–11, 13, 14, 16–21, 23, 24, 28–30, 33–36 – общий вид пыльцевого зерна с экватора. 3, 5, 8, 12, 15, 22, 25, 26, 31, 32 – общий вид пыльцевого зерна с полюса.
Масштабные линейки, мкм: 1–36 – 10.
Plate I . Pollen morphology of the Androsace (LM)
1–6 – section Samuelia; 1–2 – A. gmelini; 3–4 – A. henryi; 5–6 – A. umbellata; 7–9 section Aizoidium; 7–8 – A. aizoon; 9 – A. coccinea; 10–21 – section Chamaejasme; 10 – A. alascana; 11 – A. bungeana; 12–14 – A. incana; 15–18 – A. strigillosa; 19‒21 – A. villosa; 22–24 – section Orthocaulon, A. erecta; 25–30 – section Andraspis; 25–28 – A. albana; 29–30 – A. elongata; 31–35 – section Megista, A. turczaninowii; 36 – Douglasia ochotensis. 1, 2, 4, 6, 7, 9–11, 13, 14, 16–21, 23, 24, 28–30, 33–36 – equatorial view of pollen grain. 3, 5, 8, 12, 15, 22, 25, 26, 31, 32 – polar view of pollen grain.
Scale bars, μm: 1–36 – 10.
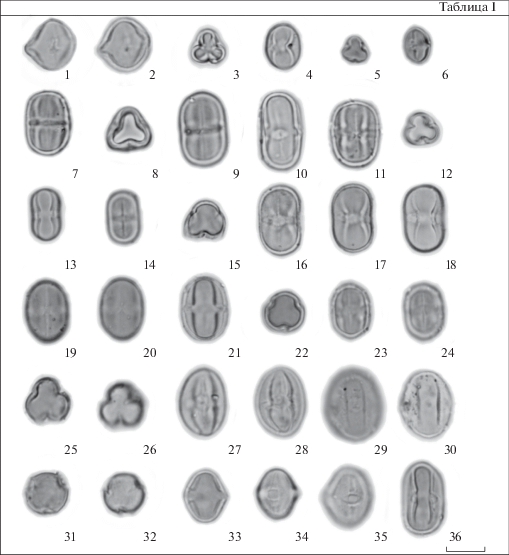
Как известно, форму пыльцевого зерна определяют по соотношению размеров полярной оси и экваториального диаметра (P/E) (Erdtman, 1952). Исходя из полученных значений P/E, пыльцевые зерна почти всех изученных видов имеют эллипсоидальную форму (P/E > 1). Исключение составляет A. filiformis, у этого вида пыльца почти сфероидальная (табл. III , 1, 2). В препаратах пыльцы A. gmelini (табл. I , 1, 2) наряду с эллипсоидальными пыльцевыми зернами встречается довольно много пыльцы почти сферической формы. В зависимости от значения P/E можно выделить несколько вариантов формы пыльцевого зерна у изученных видов Androsace.
P/E = 1, пыльца почти сфероидальная (A. filiformis).
P/E = 1.2, пыльца широкоэллипсоидальная (A. fedtschenkoi, A. maxima, A. turczaninowii).
P/E = 1.3–1.5, пыльца эллипсоидальная (A. acbaitalensis, A. albana, A. bungeana, A. capitata, A. elongata, A. erecta, A. henryi, A. incana, A. septentrionalis, A. umbellata).
P/E > 1.5, пыльца вытянуто-эллипсоидальная (A. aizoon, A. alascana, A. alaschanica, A. coccinea, A. chamaejasme, A. rotundifolia, A. spinulifera, A. stri-gillosa).
Выделенные группы не имеют четких границ, так как встречаются виды, форма пыльцы которых может считаться промежуточной между описанными выше группами. Примером может служить пыльца видов A. chamaejasme, A. triflora, A. villosa со значением P/E = 1.5–1.6. Кроме того, в препаратах A. gmelini встречаются почти сфероидальные (P/E = 1) и эллипсоидальные пыльцевые зерна (P/E = 1.3).
Важными палиноморфологическими признаками являются очертания пыльцевого зерна, прежде всего очертание пыльцы в экваториальном положении. По этому признаку можно выделить 4 группы: почти округлые (табл. I , 1, 2; табл. III , 1, 2), эллиптические (табл. I , 29, 30, 34, 35), сжато-округло-прямоугольные (табл. I , 13) и округло-прямоугольные (табл. I , 7, 11). Так же как в случае с формой, деление на группы по очертаниям – условно из-за наличия переходных форм.
В очертании с полюса пыльцевые зерна могут быть 3-лопастными (табл. I , 25, 26), от слабо-3-лопастных до почти округлых (табл. I , 31, 32) и округло-треугольными (табл. I , 8). Для зерен, имеющих эллиптические очертания с экватора, чаще характерно 3-лопастное или слабо-3-лопастное очертание с полюса. Тогда как округло-4-угольные в очертании с экватора пыльцевые зерна обычно имеют округло-треугольные очертания с полюса. Следует отметить, что в препаратах с сильно вытянутыми по полярной оси пыльцевыми зернами положение зерен с полюса встречается крайне редко.
Апертуры у пыльцы изученных видов сложные. Наружная апертура представлена длинной, очень узкой, практически щелевидной, бороздой с ровными краями и острыми концами. СЭМ позволяет рассмотреть скульптуру мембраны борозды. В большинстве случаев поверхность мембраны шероховатая. И только у пыльцевых зерен A. maxima и A. turczaninowii из секции Megista мембрана борозд крупно-зернистая. Число борозд у большинства видов стабильно, однако в образце A. spinulifera кроме типичных для рода 3-бороздно-оровых зерен встречаются одиночные 2-бороздно-оровые. Вариация числа борозд явление довольно частое, как в целом среди цветковых, так и в семействе Primulaceae. Вариабельность числа апертур подробно описана у пыльцы многих видов рода Primula (Grigoryeva et al., 2018; Britskii et al., 2019).
Эндоапертуры (оры) у пыльцы разных видов Androsace различаются размерами и очертаниями, от едва заметных до хорошо очерченных. По форме можно выделить 3 основных морфологических типа ор: округлые или слегка вытянутые по полярной оси или по экваториальному диаметру (табл. I , 4, 27, 29), эллиптические, вытянутые по экваториальному диаметру (табл. I , 10) и экваториальный поясок (endocingula), образованный за счет слияния концов длинных экваториально вытянутых ор (табл. I , 7, 9).
Светооптический микроскоп не позволяет рассмотреть скульптуру экзины. На фотографиях, полученных с помощью СМ пыльцевые зерна проломников выглядят бесцветными, прозрачными, гладкими (табл. I ). Использование же сканирующего электронного микроскопа позволяет не только рассмотреть детали рельефа поверхности, но и выделить 4 основных варианта скульптуры.
1. Перфорированная. Просветы очень мелкие, не более 0.1 мкм в диаметре, округлые или вытянутые, расстояние между ними, как правило, превышает размеры самих перфораций (табл. II , 3).
Таблица II . Морфология пыльцы представителей рода Androsace (секция Chamaejasme) (СЭМ)
1–3 – A. capitata; 4–6 – A. villosa; 7–10 – A. triflora; 11–12 – A. chamaejasme. 1, 4, 7 – общий вид пыльцевого зерна с полюса. 2, 5, 8, 10, 11 – общий вид пыльцевого зерна с экватора. 3, 6, 9, 12 – скульптура поверхности.
Масштабные линейки, мкм: 1–12 – 1.
Plate II . Pollen morphology of the Androsace section Chamaejasme (SEM)
1–3 – A. capitata; 4–6 – A. villosa; 7–10 – A. triflora; 11–12 – A. chamaejasme.
1, 4, 7 – polar view of pollen grain. 2, 5, 8, 10, 11 – equatorial view of pollen grain. 3, 6, 9, 12 – detail of ornamentation.
Scale bars, μm: 1–12 – 1.
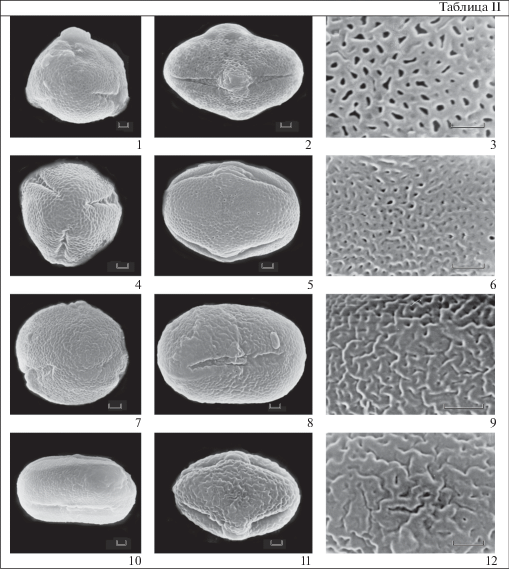
2. Складчато-перфорированная. Складки короткие извитые, между ними довольно редкие перфорации. Ширина складок у пыльцы разных видов различается. Так, широкие складки, до 0.5 мкм шириной характерны для экзины пыльцевых зерен A. chamaejasme. Тогда как у пыльцы большинства видов они более узкие и не превышают 0.2 мкм, как например, у пыльцы A. triflora (табл. II , 6, 9, 12).
3. Микросетчатая. Просветы округлые, слегка вытянутые или неправильной формы, до 0.8 мкм длиной. Стенки ячей узкие, 0.4–0.5 мкм шириной, слегка извитые, гладкие. Самая крупная сетчатость поверхности экзины характерна для пыльцы A. fedtschenkoi (табл. III , 9).
Таблица III . Морфология пыльцы представителей рода Androsace (секции Andraspis и Megista) (СЭМ)
1–3 – A. filiformis; 4–5 – A. elongata; 6–7 – A. septentrionalis; 8–9 – A. fedtschenkoi; 10–12 – A. turczaninowii; 13–15 – A. maxima. 1, 5, 10, 13 – общий вид пыльцевого зерна с полюса. 2, 4, 6, 8, 11, 14 – общий вид пыльцевого зерна с экватора. 3, 7, 9, 12, 15 – скульптура поверхности.
Масштабные линейки, мкм: 1–15 – 1.
Plate III . Pollen morphology of the Androsace sections Andraspis and Megista (SEM)
1–3 – A. filiformis; 4–5 – A. elongata; 6–7 – A. septentrionalis; 8–9 – A. fedtschenkoi; 10–12 – A. turczaninowii; 13–15 – A. maxima. 1, 5, 10, 13 – polar view of pollen grain. 4, 6, 8, 11, 14 – equatorial view of pollen grain. 3, 7, 9, 12, 15 – detail of ornamentation.
Scale bars, μm: 1–15 – 1.
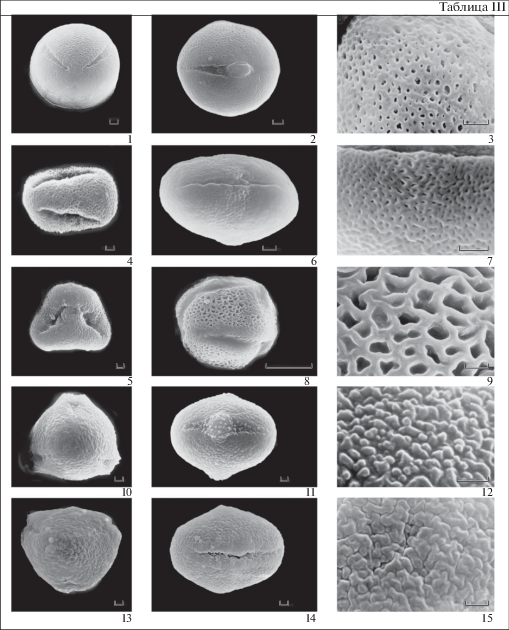
4. Мелкобугорчатая. Характерна для пыльцы A. maxima и A. turczaninowii. Бугорки мелкие, невысокие, 0.4–0.5 мкм в диаметре, округлые, вытянутые или разветвленные, довольно плотно прилегают друг к другу или расположены рыхло, на расстоянии друг от друга (табл. III , 12, 15).
Как и в случае с другими палиноморфологическими признаками выделенные группы условны, поскольку существуют переходные варианты скульптуры, которые сложно отнести к какому-то одному из перечисленных выше типов. В группе сладчато-перфорированой скульптуры экзины можно построить размерный ряд с постепенными переходами от широких складок к узким. Переходные формы существуют между мелкоскладчатой и перфорированной скульптурой, а также между перфорированной и сетчатой скульптурой.
Палиноморфологический анализ секций рода Androsace.
Секция Samuelia (A. gmelini, A. henryi, A. rotundifolia, A. umbellata) (табл. I , 1–6, табл. 1).
Палиноморфологически секция неоднородна.
Пыльца мелкая, длина полярной оси составляет 15 мкм. Очертания с экватора у разных видов либо эллиптические, либо округло-прямоугольные. P/E колеблется от 1.0 до 1.6. Оры нечетко очерченные, б. м. округлые или вытянутые по экватору. Нечеткие оры – признак, характерный для пыльцы всех изученных видов проломников из этой секции. Скульптура перфорированная или микросетчатая.
Секция Aizoidium (A. aizoon, A. coccinea) (табл. I , 7–9).
Пыльцевые зерна крупнее чем у видов из предыдущей секции. Полярная ось 17.3–20.7 мкм. В очертании с экватора округло-прямоугольные, слегка сжатые по экватору. P/E от 1.7 до 1.9. Эндоапертуры экваториально вытянутые, сливаются или почти сливаются, образуя экваториальный поясок. Скульптура складчато-перфорированная.
Пыльца изученных видов различается незначительно. У пыльцы A. aizoon по сравнению с пыльцой A. coccinea меньше значение P/E и в меньшей степени выражено сжатие в экваториальной области.
Секция Chamaejasme (A. acbaitalensis, A. alaschanica, A. alascana, A. bungeana, A. capitata, A. incana, A. chamaejasme, A. spinulifera, A. strigillosa, A. triflora, A. villosa) (табл. I , 10–21, табл. II ).
Палиноморфологически секция неоднородна. Длина полярной оси пыльцевых зерен варьирует в диапазоне от 12.7 до 21.9 мкм. Соотношение P/E колеблется от 1.3 до 2.1. В очертании с экватора эллиптические или округло-прямоугольные. Эндоапертуры большей частью эллиптические, вытянутые по экватору или сливаются концами, образуя экваториальный поясок. Скульптура от микросетчатой до складчато-перфорированной. Пыльца A. chamaejasme отличается от пыльцы других видов секции широкими складками на поверхности экзины.
Секция Orthocaulon. Из этой секции изучена пыльца только 1 вида A. erecta (табл. I , 22–24). Полярная ось 14.1–15.3 мкм, очертания пыльцевых зерен с экватора округло-прямоугольные, P/E = = 1.5. Оры эллиптические вытянуты по экватору. Скульптура микросетчатая.
Секция Andraspis (A. albana, A. elongata, A. fed-tschenkoi, A. filiformis, A. lactiflora, A. septentrionalis) (табл. I , 25–30, табл. III ), палиноморфологически полиморфная. Длина полярной оси варьирует в диапазоне от 10.3 мкм до 23.1 мкм. В очертании с экватора пыльцевые зерна почти округлые, эллиптические, реже округло-прямоугольные. Значение P/E от 1 до 1.8. Оры разнообразны: округлые или слегка вытянутые по полярной оси, слегка вытянутые по экватору, экваториально вытянутые, образующие экваториальный поясок. Скульптура микросетчатая или складчато-перфорированная. У пыльцевых зерен разных видов с микросетчатой скульптурой экзины варьируют размеры скульптурных элементов.
Секция Megista (A. maxima, A. turczaninowii) (табл. I , 31–35, табл. III , 10–15). Длина полярной оси 14.9–19.6 мкм, в очертании с экватора пыльцевые зерна эллиптические. Значения P/E от 1.2 до 1.25. Оры четко очерченные, эллиптические, вытянутые по экватору, с острыми концами. Скульптура мелко бугорчатая. Экзина пыльцы A. maxima и A. turczaninowii различается элементами скульптуры. Так у пыльцы A. turczaninowii бугорки в очертании почти округлые и располагаются на расстоянии друг от друга, тогда как у пыльцевых зерен A. maxima бугорки большей частью вытянутые и плотно прилегают друг к другу. Кроме того, пыльца A. maxima немного крупнее пыльцы A. turczaninowii.
Основные палиноморфологические характеристики видов из большинства секций сходны, что не позволяет использовать полученные данные для разграничения секций. Исключение составляет секция Megista. Пыльца видов этой секции отличается от пыльцы всех остальных изученных видов Androsace, прежде всего, мелко бугорчатой скульптурой. Некоторые авторы помещают A. maxima в секцию Andraspis (Wang et al., 2004), однако морфологические признаки пыльцы поддерживают выделение этого вида в самостоятельную секцию. Особенности строения пыльцы подтверждают самостоятельность A. turczaninowii (Shishkin, Bobrov, 1952), который по данным IPNI считается синонимом A. maxima.
Таким образом, сравнительное палиноморфологическое исследование показало, что важными диагностическими признаками пыльцы разных видов рода Androsace являются: форма и очертания пыльцевых зерен, особенности скульптуры экзины, строение апертур и прежде всего морфологические особенности эндоапертур (их очертания, размеры, расположение).
Следует отметить, что при изучении пыльцы проломников важнейшее значение имеют данные светооптической микроскопии, из всех важных палиноморфологических признаков только для изучения скульптуры нужен СЭМ.
Род Douglasia (табл. I , 36, табл. IV , 1–2). Изучена пыльца 7 видов, описания приведены в табл. 1.
Таблица IV . Морфология пыльцы представителей родов Douglasia, Vitaliana, Primula (СЭМ)
1–2 – D. ochotensis; 3–4 – V. primuliflora; 5–6 – P. darialica; 7–9 – P. forbesii; 10–12 – P. longiscapa; 13–15 – P. malacoides. 1, 3, 8, 11, 14 – общий вид пыльцевого зерна с экватора. 5, 7, 10, 13 – общий вид пыльцевого зерна с полюса. 2, 4, 6, 9, 12, 15 – скульптура поверхности.
Масштабные линейки, мкм: 1–15 – 1.
Plate IV . Pollen morphology of the Douglasia, Vitaliana, Primula (SEM)
1–2 – D. ochotensis; 3–4 – V. primuliflora; 5–6 – P. darialica; 7–9 – P. forbesii; 10–12 – P. longiscapa; 13–15 – P. malacoides. 1, 3, 8, 11, 14 – equatorial view of pollen grain. 5, 7, 10, 13 – polar view of pollen grain. 2, 4, 6, 9, 12, 15 – detail of ornamentation.
Scale bar, μm: 1–15 – 1.
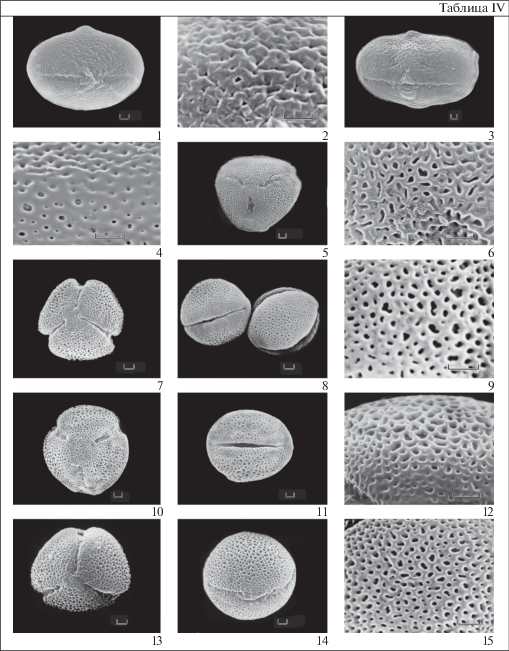
Пыльцевые зерна радиально-симметричные, изополярные, 3-бороздно-оровые. Самые крупные пыльцевые зерна у D. laevigata (полярная ось 24.1–25.6, экваториальный диаметр 12.9–13.5), у остальных зерен полярная ось не превышает 22 мкм, самые мелкие зерна выявлены у D. arctica (полярная ось 18.3 мкм) и у D. ochotensis (полярная ось 18.4 мкм). Пыльцевые зерна вытянуто-эллипсоидальные. P/E у разных видов колеблется от 1.6 у D. ochotensis до 1.95 у D. arctica. Очертания с полюса 3-лопастные или округло-треугольные. С экватора эллиптические (D. arctica, D. ochotensis) или округло-4-угольные.
Борозды длинные, узкие с четкими краями и острыми концами. Оры от почти округлых или слегка вытянутых по экватору у пыльцы D. arctica, D. montana, D. nivalis до эллиптических у остальных исследованных видов. Экзина слегка разрастается над бороздой в месте расположения эндоапертуры, образуя небольшой двузубчатый замок. Замок возможно рассмотреть только при использовании СЭМ. Скульптура не просматривается при использовании светооптического микроскопа. СЭМ позволяет выявить 2 типа скульптуры: перфорированную у пыльцевых зерен D. montana и складчато-перфорированную у пыльцы остальных изученных видов.
Род Vitaliana, V. primuliflora (табл. IV , 3–4).
Пыльцевые зерна радиально-симметричные, изополярные, 3-бороздно-оровые. Средних размеров. Полярная ось 26.3–27.8 мкм, экваториальный диаметр 14.4–15.5 мкм. P/E = 1.8. В очертании с экватора округлопрямоугольные, с полюса округлотреугольные. Борозды длинные, узкие с четкими краями и острыми концами. Оры крупные (табл. 1), округлые, четко очерченные. Скульптура перфорированная с мелкими неглубокими складками. Перфорации разноразмерные, не более 0.2 мкм в диаметре, округлые, погруженные.
Род Pomatosace, P. filicula.
Пыльцевые зерна радиально-симметричные, изополярные, 3-бороздно-оровые, мелкие (полярная ось не более 19 мкм), P/E = 1.7, в очертании с экватора округлопрямоугольные, с полюса слабо трехлопастные. Борозды длинные, узкие с четкими краями и острыми концами. Оры часто нечеткие, вытянуты по экваториальной оси. Скульптура перфорированная.
Род Primula (табл. IV , 5–15).
Для палиноморфологического сравнения родов Androsace и Primula была изучена пыльца 6 видов Primula с 3-бороздно-оровой пыльцой (P. darialica, P. farinifolia, P. forbesii, P. longiscapa, P. malacoides, P. zeylamica).
Изученные пыльцевые зерна радиально-симметричные, изополярные, 3-бороздно-оровые, у большинства видов мелкие (длина полярной оси не более 17 мкм). Только у P. malacoides пыльцевые зерна средних размеров (полярная ось 28.4–30.2 мкм, экваториальный диаметр 23.5–25.5 мкм). Большей частью эллипсоидальные (P/E > 1), реже почти сфероидальные (P/E = 1) или широкоэллипсоидальные (P/E < 1). В очертании с экватора эллиптические, почти округлые или широкоэллиптические, с полюса слабо 3-лопастные. Борозды длинные, узкие. Оры большей частью нечеткие. Скульптура микросетчатая.
Сравнительный палиноморфологический анализ родов Androsace, Douglasia, Pomatosace, Vita-liana, Primula.
По мнению многих систематиков, род Androsace считается не менее полиморфным чем близкородственный ему род Primula. Сравнительный палинологический анализ показал, что пыльца Androsace менее разнообразна, чем пыльца разных видов Primula, для которой описано 3 основных палинотипа и 20 дополнительных подтипов (Punt et al., 1974). Изученный материал и литературные данные позволяют утверждать, что для рода Androsace характерен один 3-борозно-оровый палинотип. Хотя в пределах этого палинотипа пыльца разных видов довольно разнообразна. Палиноморфологический анализ полученных данных и имеющихся в литературе сведений показал, что и в роде Androsace есть виды, пыльца которых сходна с пыльцой некоторых видов Pri-mula. Большинство видов Primula характеризуется сплющенными 3-слитнобороздными пыльцевыми зернами, однако есть небольшое число видов примул (например, P. darialica, P. farinifolia, P. forbesii, P. longiscapa, P. malacoides, P. zeylamica и др.) с эллипсоидальной 3-бороздно-оровой пыльцой, нечеткими орами и перфорированной или мелкосетчатой скульптурой экзины. Сходными палиноморфологическими чертами обладают и некоторые виды Androsace, такие как, например, A. gmelini, A. henryi, A. rotundifolia. В дисперсном состоянии определить принадлежат такие пыльцевые зерна видам Androsace или Primula не представляется возможным. Существование видов с такой пыльцой (3-бороздно-оровые, с нечеткими орами и перфорированной скульптурой) не позволяет провести четкую границу между родами, несмотря на то что пыльца большинства видов Primula хорошо отличается от пыльцы большинства видов Androsace. Таким образом, полученные нами данные не согласуются с утверждением Wendelbo (1961) об отсутствии какого-либо палинологического сходства между родами Primula и Androsace.
Также определенный интерес представляет палиноморфологическое сравнение родов Androsace и Douglasia. Большинство авторов, особенно североамериканских, считают Douglasia самостоятельным родом (Constance, 1938; Robbins, 1944; Kelso, 1992; Kelso et al., 1994), тогда как многие европейские исследователи рассматривают Douglasia лишь в ранге секции в пределах рода Androsace (Shishkin, Bobrov, 1952; Wendelbo, 1961; Kress, 1965; Smith, Lowe, 1997). Сравнительный палиноморфологический анализ Androsace и Douglasia показал, что по основным признакам пыльцы (форма, очертания с экватора, апертуры, скульптура) эти два рода не различаются. В литературе отмечают, что пыльцевые зерна видов Douglasia крупнее пыльцы Androsace (Xu et al., 2016). Однако, наше исследование показало, что ряд видов Androsace (например, A. alaschanica, A. bungeana, A. coccinea, A. fedtschenkoi и некоторые другие) имеют пыльцу, близкую по размерам пыльце Douglasia (табл. 1). Молекулярные исследования также указывают на положение Douglasia среди Androsace (Trift et al., 2002; Martins et al., 2003). Таким образом, палинологические сведения полностью согласуются с современными молекулярно-генетическими данными.
До сих пор продолжается обсуждение статуса рода Vitaliana. Благодаря наличию гетеростилии этот род считается уникальным в трибе Androsaceae и на основании этого некоторые авторы трактуют его как самостоятельный род (Ferguson, 1972; Smith, Lowe, 1997 и др.), другие как секцию в роде Androsace (Wendelbo, 1961; Kress, 1965, 1967). Современные молекулярно-генетические исследования свидетельствуют о том, что виды рода Vitaliana должны быть включены в состав Androsace (Trift et al., 2002; Martins et al., 2003; Dixon et al., 2016). Полученные нами палиноморфологические данные показывают, что по форме, очертаниям пыльцевых зерен и по перфорированной скульптуре экзины пыльца Vitaliana сходна с пыльцой многих видов Androsace. Главным отличительным признаком пыльцевых зерен Vitaliana являются крупные (около 5 мкм в диаметре), четко очерченные округлые оры. Таких эндоапертур у пыльцы исследованных видов Androsace не было обнаружено. Кроме того, от пыльцы большинства Androsace пыльца Vitaliana отличается значительно большими размерами (табл. 1).
Пыльцевые зерна рода Pomatosace не отличаются от пыльцевых зерен исследованных видов рода Androsace, что согласуются с ранее полученными палиноморфологическими данными (Wendelbo, 1961; Spanowsky, 1962). Нами не обнаружено никаких палиноморфологических черт (форма зерна, апертуры, скульптура экзины), которые, по мнению некоторых авторов (Xu et al., 2016) позволяют отличить морфотип пыльцы Pomatosace от других подгрупп Androsace. Последние молекулярно-филогенетические исследования поддерживают слияние Pomatosace и Androsace (Schneeweiss et al., 2004; Wang et al., 2004; Boucher et al., 2012; de Vos et al., 2014) и палинологические данные, на наш взгляд, не противоречат такому объединению родов в один таксон.
Таким образом, сравнительное исследование пыльцы видов Androsace показало, что род характеризуется одним (3-бороздно-оровым типом пыльцы). В пределах этого типа пыльцевые зерна разных видов хорошо различаются размерами, формой, очертанием, особенностями эндоапертур и скульптурой экзины. Во всех секциях рода, кроме секции Megista, встречаются виды с похожей пыльцой, что затрудняет использование признаков пыльцы для разграничения секций, установленных по макроморфологическим признакам.
Обнаруженное большое сходство пыльцы родов Androsace, Douglasia, Pomatosace и Vitaliana, вероятно, свидетельствует о их близком родстве и не противоречит объединению этих родов в один таксон.
Список литературы
Anderberg A.A., Kelso S. 1996. Phylogenetic implications of endosperm cell wall morphology in Douglasia, Androsace and Vitaliana (Primulaceae). – Nord. J. Bot. 16 (5): 481–486. https://doi.org/10.1111/j.1756-1051.1996.tb00262.x
Boucher F.C., Thuiller W., Roquet C., Douzet R., Aubert S., Alvarez N., Lavergne S. 2012. Reconstructing the origins of high-alpine niches and cushion life form in the genus Androsace s. l. (Primulaceae). – Evolution. 66 (4): 1255–1268. https://doi.org/10.1111/j.1558-5646.2011.01483.x
[Britskii et al.] Брицкий Д.А., Иванова А.Н., Григорьева В.В. 2019. Морфология многобороздных пыльцевых зерен некоторых видов Primula (Primulaceae). – Бот. журн. 104 (10): 1610–1621.
Constance L. 1938. A revision of the genus Douglasia Lindl. – Am. Midl. Nat. 19 (1): 249–259.
Dixon C.J., Gutermann W., Schönswetter P., Schneeweiss G.M. 2016. Taxonomy and nomenclature of the polymorphic European high mountain species Androsace vitaliana (L.) Lapeyr. (Primulaceae). – PhytoKeys 75: 93–106. https://doi.org/10.3897/phytokeys.75.10731
Erdtman G. 1952. Pollen morphology and taxonomy. Angiosperms. Stockholm. 539 p.
Ferguson I.K. 1972. Vitaliana Sesler, Androsace L. – In: Flora Europaea. Vol. 3. Cambridge. P. 20–23.
[Grigoryeva et al.] Григорьева В.В., Брицкий Д.А., Иванова А.Н., Коробков А.А. 2018. Морфология пыльцевых зерен некоторых видов секции Farinosae рода Primula (Primulaceae). – Бот. журн. 103 (5): 645–654.
Hu C.M. 1994. On the geographical distribution of the Primulaceae. – J. Trop. Subtrop. Bot. 2 (4): 1–14.
Hu C.M., Yang Y.C. 1986. A revision of the genus Androsace L. in China. – Acta Phytotaxon. Sin. 24 (3): 215–232.
Hu C.M., Kelso S. 1996. Primulaceae. – In: Flora of China. Vol. 15. St. Louis. P. 80–99.
Kelso S.1992. Conspectus of the genus Douglasia (Primulaceae) with comments on Douglasia alaskana, an Alaska-Yukon alpine endemic. Can. J. Bot. 70 (3): 593–596. https://doi.org/10.1139/b92-076
Kelso S., Yurtsev B.A., Murray D.F. 1994. Douglasia beringensis (Primulaceae): A new species from northwestern Alaska. – Novon. 4 (4):381–385. https://doi.org/10.2307/3391448
Kim Y.S., Kim H.K. 1988. A taxonomic study on genus Androsace in Korea. – Kor. J. Plant Tax. 18 (3): 233–262.
Kress A. 1965. Zur Zytotaxonomie der Androsace-Vitaliana-Douglasia-Verwandtschaft. – Mitt. Bot. Staatssamml. München. 5: 653–674.
Kress A. 1967. 104. Familie Primulaceae. Nachträge, Berichtigungen und Ergänzungen. – In: Illustrierte Flora von Mitteleuropa. Vol. 5. München. P. 2246b–2250b.
[Kupriyanova, Aleshina]. Куприянова Л.А., Алешина Л.А. 1967. Палинологическая терминология покрытосеменных растений. Л. 84 с.
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1972. Пыльца и споры растений флоры европейской части СССР. Т. 1. Л. 184 с.
[Kupriyanova, Aleshina]. Куприянова Л.А., Алешина Л.А. 1978. Пыльца двудольных растений флоры европейской части СССР. Л. С. 84.
Mabberley D.J. 2009. Mabberley’s plant-book: a portable dictionary of plants, their classification and uses. Cambridge. 920 p.
Martins L., Oberprieler C., Hellwig F.H. 2003. A phylogenetic analysis of Primulaceae s. l. based on internal transcribed spacer (ITS) DNA sequence data. – Plant Syst. Evol. 237 (3): 75–85. https://doi.org/10.1007/s00606-002-0258-1
Mast A.R., Kelso S., Richards A.J., Lang D.J., Feller D.M.S., Conti E. 2001. Phylogenetic relationships in Primula L. and related genera (Primulaceae) based on noncoding chloroplast DNA. – Int. J. Plant Sci. 162 (6): 1381–1400.
Nasir Y.J. 1986. Pollen morphological studies in the Androsace (Primulaceae) species found in Pakistan. – Pak. J. Bot. 18 (2): 301–307.
Pax F. 1889. Monographische Übersicht über die Arten der Gattung Primula. Botanische Jahrbücher fur Systematik, Pflanzengeschichte und Pflanzengeographie. 10: 75–241.
Punt W., Weenen D.L.V., Van Dostrum W.A.P. 1974. The Northwest European Polen Flora 3. Primulaceae. – Rev. Paleobot. Palynol. 17 (3/4): 31–70.
Robbins G.T. 1944. North American species of Androsace. – Am. Midl. Nat. 32 (1): 137–163. https://doi.org/10.2307/2421367
[Shishkin, Bobrov] Шишкин Б.К., Бобров Е.Г. 1952. Род 1118. Androsace L. – В кн.: Флора СССР. Т. 18. М.–Л. С. 221–243.
Smith G., Lowe D.B. 1997. The genus Androsace: a monograph for gardeners and botanists. Pershore, Worcestershire. 208 p.
Schneeweiss G.M., Schönswetter P., Kelso S., Niklfeld H. 2004. Complex Biogeographic Patterns in Androsace (Primulaceae) and Related Genera: Evidence from Phylogenetic Analyses of Nuclear Internal Transcribed Spacer and Plastid trnL-F Sequences. – Syst. Biol. 53 (6): 856–876. https://doi.org/10.1080/10635150490522566
Spanowsky W. 1962. Die Bedeutung der Pollenmorphologie für die Taxonomie der Primulaceae-Primuloideae. – Feddes Rep. 65 (3): 149–214.
Trift I., Källersjö M., Anderberg A.A. 2002. The monophyly of Primula (Primulaceae) evaluated by analysis of sequences from the chloroplast gene rbcL. – Systematic Botany. 27 (2): 396–407. https://www.jstor.org/stable/3093879
de Vos J.M., Hughes C.E., Schneeweiss G.M., Moore B.R., Conti E. 2014. Heterostyly accelerates diversification via reduced extinction in primroses. – Proc. Biol. Sci. 281 (1784): 20140075. https://doi.org/10.1098/rspb.2014.0075
Wang Y.J., Li X.J., Hao G., Liu J.Q. 2004. Molecular phylogeny and biogeography of Androsace (Primulaceae) and the convergent evolution of cushion morphology. – J. Syst. Evol. 42 (6): 481–499. http://www.jse.ac.cn/EN/Y2004/V42/I6/481
Wendelbo P. 1961. Studies in Primulaceae III. On the genera related to Primula with special reference to their pollen morphology. – Arb. Univ. Bergen Mat. Nat. 19: 1–31.
Xu Y., Hu C.-M., Hao G. 2016. Pollen morphology of Androsace (Primulaceae) and its systematic implications. – J. Syst. Evol. 54 (1): 48–64. https://doi.org/10.1111/jse.12149
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал