

Ботанический журнал, 2021, T. 106, № 3, стр. 229-238
SCUTELLARIA OREOPHILA (LAMIACEAE): ЖИЗНЕННАЯ ФОРМА, ПОБЕГООБРАЗОВАНИЕ, ОНТОГЕНЕЗ
В. А. Черемушкина 1, *, А. Ю. Асташенков 1, **
1. Центральный сибирский ботанический сад СО РАН
630090 Новосибирск, ул. Золотодолинская, 101, Россия
* E-mail: cher.51@mail.ru
** E-mail: astal@bk.ru
Поступила в редакцию 23.04.2020
После доработки 29.09.2020
Принята к публикации 14.10.2020
Аннотация
В статье приводятся результаты исследования онтогенеза и особенностей развития особей эндемика Кавказа Scutellaria oreophila L. (Lamiaceae). На основании изучения побегообразования установлен биоморфологический портрет вида, который характеризует S. oreophila как явнополицентрический симподиально нарастающий длиннокорневищный травянистый поликарпик, гемикриптофит. Установлено, что на низкотравном альпийском лугу особи S. oreophila в основном представляют собой совокупность клонов, состоящих из рамет разных онтогенетических состояний. Онтогенез особей сложный и состоит из онтогенеза семенной особи (генеты) и дочерних особей вегетативного происхождения (рамет). Взрослая особь S. oreophila представляет собой явнополицентрическую систему парциальных кустов и побегов, связанных между собой длинными симподиально сочлененными гипогеогенными коммуникационными корневищами, их интенсивное ветвление, приводящее к пространственному захвату территории, характеризует жизненную форму изученного вида как длиннокорневищную.
Комплексное изучение флоры и растительности Кавказа является важной проблемой сохранения биоразнообразия. Уникальные почвенно-климатические условия и геологическая история этого региона способствуют процессам видообразования, приводящим к значительному числу эндемичных растений (Grossheim, 1936; Ivanov, 1998). Род Scutellaria L., один из крупных родов сем. Lamiaceae, характеризуется значительным полиморфизмом видов и недостаточно изучен. Биоморфологические исследования с целью уточнения морфологии и экологических особенностей видов почти не проводились, тогда как они также могут быть использованы в систематике рода. Практически не изучены онтогенез и жизненные формы представителей Scutellaria, которые раскрыли бы внутриродовые взаимоотношения и в конечном итоге могли бы быть учтены при создании более совершенной системы рода. Изучение биоморфологии растений позволит выявить механизмы адаптации, обеспечивающие их участие в структуре разнообразных растительных сообществ. Среди кавказских представителей Scutellaria значительная их часть относится к эндемикам Кавказа, которые встречаются во всех горных поясах от степного до альпийского.
Scutellaria oreophila Grossh. относится к подроду Euscutellaria Briq., секции Lupulinaria A.Ha-milt. Этот вид имеет близкое родство к S. orientalis L. и рядом авторов понимается либо как самостоятельный таксон, либо как раса (Yuzepchuk, 1954; Grossheim, 1967; Menitsky, 1992). Описан А.А. Гроссгеймом по экземплярам, собранным в окрестностях села Крыз Кубинского (в настоящее время Губинского) района Азербайджана. Тип в Баку.
S. oreophila – кавказский эндемик (Grossheim, 1967), относится к эукавказскому геоэлементу (Portnier, 2000; Ivanov, 2019). Его ареал охватывает район Большого Кавказа и горного Предкавказья в пределах России и северо-востока Азербайджана. В Восточном Закавказье S. oreophila распространен в основном в субальпийском и альпийском поясах, произрастая на горных лугах, щебнистых склонах, известняковых обнажениях и осыпях (Grossheim, 1949, 1967; Isaev, 1957) на высотах 2000–2500 м над ур. м. В других районах Кавказского хребта и горного Предкавказья он встречается на сухих склонах среднего пояса гор (Yuzepchuk, 1954; Galushko, 1980; Murtazaliev, 2009; Tsepkova, 2012), в травяном покрове сосновых лесов с участием Pinus kochiana Klotzsch ex C. Koch на высотах 1200–1600 м над ур. м., где отмечен на крутых склонах северной и северо-восточной экспозиций на маломощных скелетных почвах, на выходах известняковых скал (Abdurakhmanova, Neshataeva, 2017; Abdurakhmanova et al., 2018).
Существуют разные точки зрения на особенности жизненной формы Scutellaria oreophila. А.А. Гроссгейм (Grossheim, 1945; 1949) понимает S. oreophila как многолетнее травянистое растение. Его точку зрения разделяют С.Ю. Юзепчук (Yuzepchuk,1954), А.И. Галушко (Galushko, 1980) и Р.А. Муртазалиев (Murtazaliev, 2009). Однако Ю.Н. Карпун (Karpun, 2002) и А.Л. Иванов (Ivanov, 2019) считают, что S. oreophila – это полукустарничек, хамефит, у которого почки возобновления находятся выше поверхности почвы. Неоднозначность в определении жизненной формы и отсутствие сведений об онтогенезе особей S. oreophila побудили нас провести детальный биморфологический анализ для решения этого спорного вопроса.
Цель работы – изучение жизненной формы и онтоморфогенеза эндемика Кавказа Scutellaria oreophila.
МЕТОДЫ ИССЛЕДОВАНИЯ
Материал для изучения жизненной формы и онтоморфогенеза был собран в locus classicus в окрестностях с. Крыз Губинского района Республики Азербайджан в сообществе низкотравного альпийского луга с выходами крупных гранитных камней на высоте 2170 м над ур. м. (координаты N 41°13359; E 48°15254).
В работе использованы структурный и эколого-морфологический подходы. Жизненная форма описана согласно классификации И.Г. Серебрякова (Serebryakov, 1964). При изучении побегообразования применяли терминологию И.Г. Серебрякова (Serebryakov, 1959), Л.Е. Гатцук (Gatzuk, 1974), Е.Л. Нухимовского (Nukhimovskiy, 1997). Цикличность побега определяли с момента развертывания почки и до отмирания надземной части побега. При изучении онтогенеза была принята концепция дискретного описания онтогенеза (Rabotnov, 1950; Uranov, 1975). Фазы морфогенеза выделены согласно обобщению, сделанному О.В. Смирновой с соавт. (Smirnova et al., 1976), и последующим добавлениям (Savinykh, Cheryomushkina, 2015).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Прорастание семян надземное. Проросток представляет собой однопобеговое растение с двумя овальными семядолями и 1–2 парами супротивных, простых длинночерешковых настоящих листьев с пластинкой эллиптической формы и городчатым краем. Длина пластинки листа 0.6–0.8 см, ширина не более 0.5 см. В пазухах всех листьев закладываются почки. Гипокотиль хорошо выражен, достигает 1 см длины, эпикотиль короткий и не превышает 0.4–0.6 см. Главный корень светло-коричневого цвета, слабоветвистый, его длина не более 1.5–2.0 см. Длительность состояния 2–3 недели.
Особи S. oreophila переходят в ювенильное состояние (j) в год прорастания семян. Побег продолжает нарастать моноподиально, образуя ортотропную ось, состоящую из 6–7 метамеров с ассимилирующими листьями (рис. 1.I). Длина метамеров постепенно уменьшается по направлению к верхушечной почке. Сближение метамеров в апикальной части побега приводит к формированию розеточной части, в результате чего образуется верхнерозеточный побег. В пазухах всех листьев закладываются почки. Длина такого побега не превышает 1.5–1.6 см. Главный корень втягивает гипокотиль в почву, и семядольный узел оказывается на ее поверхности. В конце вегетационного сезона первичный побег принимает плагиотропное положение. Его верхушечная розеточная часть располагается на поверхности почвы, терминальная почка прикрыта зачатками сильно опушенных настоящих листьев. Главный корень достигает длины от 2.5 до 4 см и ветвится по всей длине; возникает корневая система с боковыми корнями I–II порядков.
На второй год растения переходят в имматурное состояние (im). Сохранившаяся розеточная часть первичного побега принимает ортотропное положение и продолжает моноподиальное нарастание. Второй годичный прирост не превышает 1 см. Первый и второй годичные приросты отделены друг от друга участком сближенных метамеров. На втором годичном приросте формируются 3–4 коротких и 5–7 длинных метамеров. Первичный побег ветвится, из зимующей почки семядольного или первого метамера развертывается ортотропный побег II порядка до 0.5 см длиной, несущий 3–4 пары зеленых листьев и представляющий собой первый годичный прирост верхнерозеточного дициклического побега возобновления. В пазухах листьев закладываются почки. В конце вегетационного сезона первичный побег отмирает до узла с побегом возобновления. Терминальная почка побега возобновления располагается в подстилке. Главный корень удлиняется до 6–7 см, на нем развивается много эфемерных тонких боковых корней.
На третий год растения переходят в виргинильное онтогенетическое состояние (v). Моноподиальное нарастание особи сменяется на симподиальное. Второй годичный прирост побега возобновления восходящий, высотой не более 2.0–2.5 см, состоит из 1–2 коротких и 4–5 длинных метамеров с зелеными листьями. Заканчивается побег двумя сближенными метамерами, несущими ассимилирующие листья. Надземная часть побега ветвится слабо развитыми побегами обогащения. Одновременно с ростом второго годичного прироста побега II порядка развертываются зимующие почки, расположенные в узлах 2–3 длинных метамеров первого годичного прироста. Развиваются 2–3 побега возобновления III порядка. Нереализованные почки становятся спящими. Формируется первичный куст. По своему развитию побеги I–III порядков дициклические верхнерозеточные. Первый годичный прирост побега III порядка не превышает 1.5 см, состоит из одного короткого метамера с чешуевидными листьями и 2–3 длинных метамеров с ассимилирующими листьями. В конце вегетационного сезона надземная часть побега II порядка вместе со слаборазвитыми побегами обогащения отмирает до зоны с побегами возобновления III порядка. Вся сохранившаяся часть первичного куста втягивается в почву и утолщается. Формируется короткий каудекс. На поверхности почвы остаются только верхушечные почки побегов возобновления, рост которых продолжится на следующий год. Нереализованные почки, сохранившиеся в побеговой системе, становятся спящими. Новые побеги возобновления IV порядка также дициклические. Они развертываются весной из почек, расположенных на длинных метамерах (0.4–0.8 см) первого годичного прироста побега III порядка. Таким образом, зона возобновления в структуре дициклического побега представляет собой 2–3 длинных метамера первого годичного прироста.
Число побегов в кусте виргинильных особей насчитывается от 3 до 6, длиной 2–2.5 см. Подземная сфера особи компактная, образована симподиально сочлененными резидами побегов I–III порядков с почками возобновления и спящими почками. Гипокотиль и базальная часть главного корня утолщаются до 2 мм. Корневая система становится смешанной, тонкие придаточные корни (в числе 2–3) формируются в узлах зоны возобновления. Продолжительность состояния – 2 года.
Переход в молодое генеративное состояние (g1) происходит на 5-й год жизни особей. Первым зацветает один из дициклических побегов IV порядка, остальные побеги остаются вегетативными, их надземная часть отмирает в конце вегетационного сезона до зоны возобновления. За счет развертывания боковых почек в верхней части второго годичного прироста побег может ветвиться слаборазвитыми побегами обогащения, выполняющими функцию фотосинтеза. Заканчивается побег брактеозным, компактным, коротким, открытым соцветием кисть.
В этом состоянии у растений, кроме дициклических побегов возобновления, из спящих почек, расположенных на каудексе, развиваются дициклические корневищноудлиненные побеги, длина плагиотропной части которых может достигать 3.0–5.0 см. В первый год корневищноудлиненный побег развивается под землей, его верхушечная почка перезимовывает, плагиотропный годичный прирост становится длинным гипогеогенным корневищем. На второй год почка трогается в рост и развертывается в надземный удлиненный вегетативный побег высотой до 2.5–3.5 см. На геофильной (корневищной) части этого побега в пазухах чешуевидных листьев закладываются почки, большая часть из них остается спящими. Апогеотропная часть побега представлена 2–3 короткими и 1–2 длинными метамерами, ортотропная состоит из 3–4 длинных метамеров, несущих зеленые листья.
На основе корневищноудлиненного побега формируется парциальный куст. Он образуются за счет реализации почек возобновления, расположенных в узлах метамеров апогеотропной части побега. Эти почки трогаются в рост одновременно с ростом надземной части корневищного побега. Появившиеся из них побеги – моно- или дициклические. Надземная часть моноциклических побегов отмирает вместе с надземной частью корневищного побега. На их базальной части сохраняются нереализованные почки, из которых на следующий год могут развиться побеги следующего порядка. Дициклические побеги образуются выше сформированных моноциклических. Они представляют собой, слаборазвитые анизотропные побеги, которые на будущий год продолжат свое развитие. Дициклический корневищноудлиненный побег дает начало развитию двум моноциклическим и 2–3 дициклическим побегам возобновления. Такая разветвленная побеговая система состоит обычно из побегов 2–3 порядков ветвления. Она слабо укореняется, живет 3–4 года и отмирает до базальных частей, с расположенными на них спящими почками.
Таким образом, особи в молодом генеративном состоянии представляют собой куртину. Она состоит из первичного куста, 2–3 парциальных вегетативных побегов и парциальных кустов, соединенных с первичным кустом гипогеогенными коммуникационными корневищами. В первичном кусте развивается 1–2 анизотропных дициклических генеративных и 2–6 вегетативных разветвленных побегов, часть из них – это первый годичный прирост дициклических побегов возобновления следующих порядков, другая – моноциклические побеги, отмирающие полностью в конце вегетационного сезона. В этом состоянии продолжительность молодого генеративного состояния семенной особи составляет 3–4 года (рис. 1.I).
В подземной сфере первичный куст имеет разросшийся до 5 мм в толщину каудекс, несущий спящие почки и редкие придаточные корни, и главный корень.
Куртина средневозрастных генеративных особей (g2) состоит из первичного куста и 4–6 парциальных кустов и парциальных побегов. В первичном кусте из спящих почек развиваются корневищноудлиненные побеги, плагиотропные части которых расположены в верхнем слое почвы. Эти побеги приводят к образованию новых слабо укореняющихся парциальных кустов. Число цветущих побегов в куртине увеличивается до 8–10. Генеративные побеги анизотропные, высота надземной части не более 5–7 см. Надземная часть некоторых парциальных кустов отмирает, но их базальные участки с живыми почками сохраняются в составе куртины продолжительное время.
Каудекс состоит из базальных частей побеговой системы первичного куста, он разрастается до 1–1.5 см в диаметре и несет большое число спящих почек, увеличивается также диаметр базальной части главного корня и диаметр придаточных корней. Продолжительность состояния 7–10 лет.
В этом состоянии начинается вегетативное размножение. Перегнивание корневищ приводит к образованию клона и самостоятельному развитию парциальных кустов и побегов. Возникшие раметы находятся в разных онтогенетических состояниях (от ювенильного до средневозрастного генеративного состояния) и способны к неоднократной партикуляции (рис. 1.II).
Рис. 1.
Онтогенез особей Scutellaria oreophila.
Условные обозначения:
I – онтогенез семенной особи, II – онтогенез раметы.
Онтогенетические состояния: p – проросток, j1 – ювенильное, im –имматурное, v – виргинильное, g1 – молодое генеративное, g2 – средневозрастное, g3 – старое генеративное, ss – субсенильное, s – сенильное, ------- – уровень почвы, → – пути и возможные переходы из одного онтогенетического состояния в другое.
1 – побег верхнерозеточный, 2 – первый годичный прирост дициклического побега, 3 – побег дициклический вегетативный, 4 – побег дициклический генеративный, 5 – побег обогащения генеративный, 6 – побег обогащения вегетативный, 7 – побег корневищный, 8 – побег дициклический корневищноудлиненный (парциальный побег), 9 – корневище гипогеогенное, 10 – парциальный куст, 11 – главный корень, 12 – придаточный корень, 13 – спящая почка, 14 – каудекс.
Fig. 1. Ontogenesis of Scutellaria oreophila individuals.
Legend:
I – ontogenesis of a seed individual, II – ontogenesis of a ramet.
Ontogenetic states: p – seedling, j1 – juvenile, im – immature, v – virginal, g1 – young generative, g2 – mature generative, g3 – old generative, ss – subsenile, s – senile, ------- – soil level, → – paths and possible transitions from one ontogenetic state to another.
1 – upper-rosette shoot, 2 – first annual growth of a dicyclic shoot, 3 – dicyclic vegetative shoot, 4 – dicyclic generative shoot, 5 – generative enriching shoot, 6 – vegetative enriching shoot, 7 – rhizome shoot, 8 – dicyclic rhizome-long-shoot (partial shoot), 9 – hypogeogenic rhizome, 10 – partial bush, 11 – main root, 12 – adventitious root, 13 – dormant bud, 14 – caudex.
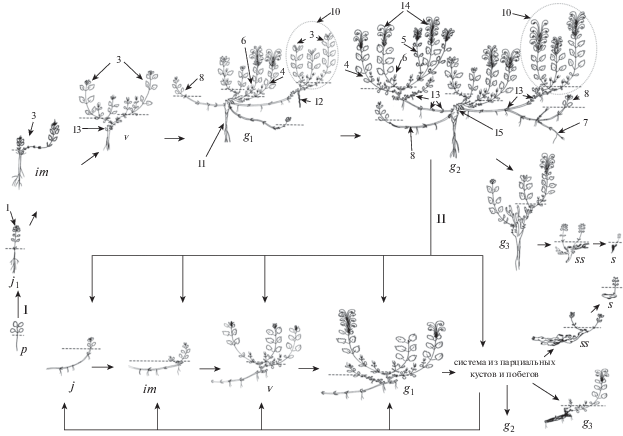
Старые генеративные особи (g3) семенного происхождения теряют способность к образованию корневищных побегов и вегетативному размножению. В первичном кусте побеги развертываются в основном из спящих почек, расположенных на каудексе и в основании побегов на коротких метамерах. Генеративные побеги единичны, они моноциклические, высотой 8.0–12.0 см, вегетативные побеги также моноциклические. Последние не переходят к цветению, остаются вегетативными и полностью отмирают в этот же год. В составе куста сохраняются отмершие остатки побеговых систем и базальные части корневищ. Длительность жизни семенных особей в этом состоянии не превышает 2–3 лет.
У особей постгенеративного периода (субсенильного (ss) и сенильного (s) состояний) семенного происхождения из спящих почек на каудексе развертываются только однолетние вегетативные побеги имматурного или ювенильного облика. После реализации всех почек онтогенез семенной особи завершается. Длительность состояния 1–3 года, а продолжительность жизни семенной особи составляет 20–23 года.
Особи семенного происхождения в сообществе встречаются очень редко. Они располагаются единично около крупных валунов, где ослаблена или отсутствует фитоценотическая конкуренция и приживание проростков не затруднено. В природных популяциях особи вида представляют собой, как правило, раметы разного онтогенетического состояния, относящиеся к разным поколениям.
Онтогенез парциальных образований (рамет) начинается с ювенильного состояния (рис. 1.II). Рамета ювенильного состояния – это одиночный побег, подземная часть которого образована участком плагиотропного корневищного побега, а надземная представляет собой анизотропный вегетативный побег до 2–3 см длины. Надземная часть побега состоит только из 2–3 удлиненных метамеров, несущих ассимилирующие листья. Придаточные корни развиваются в узлах любой части корневища. После отмирания надземной части осенью, рамета на следующий год начинает ветвиться и переходит в имматурное состояние. Из одной пары супротивно расположенных почек, сохранившихся в апогеотропной части материнского корневищного побега, развиваются два вегетативных побега замещения. Их геофильная часть состоит из 2–3 коротких метамеров и 1‒3 длинных, несущих чешуевидные и переходные листья. Надземная часть ортотропная несет 5–6 метамеров с зелеными листьями. Зоной возобновления становится первый верхний длинный метамер. На третий год рамета переходит в виргинильное онтогенетическое состояние. На базе корневищного побега формируется парциальный куст, состоящий из 4–6 вегетативных побегов. Корневищный побег утолщается до 0.2 мм, нереализованные почки на нем становятся спящими и сохраняются в течение всей жизни корневища.
На 4-й год один из дициклических побегов замещения зацветает. У молодых генеративных рамет увеличивается метамерная протяженность зоны возобновления побегов замещения. Она представляет собой геофильную часть анизотропного дициклического побега, несущую 2–3 длинных метамера. Впоследствии большинство образовавшихся побегов цветут, и рамета переходит в зрелое генеративное состояние. Геофильная (корневищная) часть каждого побега, состоящая из длинных метамеров, становится зоной возобновления побегов n + 1 порядка. Также развиваются моноциклические генеративные побеги из почек апогеотропной части материнского побега. Часть придаточных корней утолщается, особенно в апикальной части корневища. В этом состоянии полицентрическая система неоднократно партикулирует с образованием рамет от ювенильного до субсенильного состояния. В старом генеративном состоянии в рамете накапливается большое число отмерших побегов. Новые побеги развертываются в основном из спящих почек, расположенных на коротких метамерах в базальной части любого побега предыдущего порядка. Участок корневища покрывается толстой перидермой, часть придаточных корней отмирает. Парциальный куст раметы в субсенильном состоянии несет 2–3 вегетативных моноциклических побега. Сенильные раметы представлены небольшим участком сохранившегося утолщенного многолетнего корневищного побега с небольшим числом спящих почек. Как правило, развертывается только один моноциклический вегетативный побег ювенильного облика (из наиболее крупной почки), реже – два побега. Длительность жизни раметы от возникновения из верхушечной почки корневища и до отмирания в сенильном состоянии составляет 10–15 лет. Развитие корневищных побегов приводит к пространственному разрастанию особей S. oreophila, в результате чего формируется полицентрическая система из парциальных побегов и кустов, связанных разветвленными коммуникационными гипогеогенными корневищами с придаточной корневой системой (рис. 2).
Рис. 2.
Система парциальных кустов и побегов Scutellaria oreophila.
Условные обозначения:
Корневище раметы: a – ветвление акротонное с образованием рамет j состояния; b – ветвление мезотонное и базитонное с образованием рамет im–g1 состояний; c – ветвление из спящих почек многолетней части с образованием рамет ss–s состояний; d – фрагмент клона, состоящий из парциальных кустов и побегов.
1 – парциальный побег, 2 – разветвленный парциальный побег, 3 – парциальный куст, 4 – отмерший парциальный куст.
Fig. 2. The system of partial bushes and shoots of Scutellaria oreophila.
Legend:
Rhizome of a ramet: a – acrotonic branching with the formation of j state ramet; b – branching mesotonic and basitonic with the formation of im–g1 states ramets; c – branching from dormant buds of a perennial part with the formation of ss–s states ramets; d – fragment of a clone consisting of partial bushes and shoots.
1 – partial shoot, 2 – branched partial shoot, 3 – partial bush, 4 – dead partial bush.
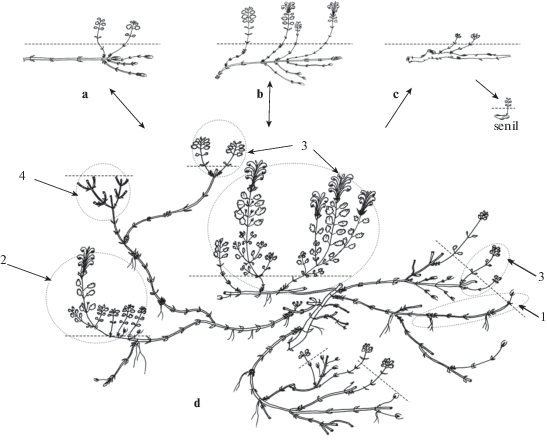
Корневище раметы, вне зависимости от онтогенетического состояния, способно ветвиться, что связано с реализацией любых почек, расположенных на нем (рис. 2). Одновременно может развиться от 4 до 6 новых корневищных побегов. Их длина варьирует от 3–4 см до 10–13 (19) см. В зависимости от возраста корневища, положения и времени раскрытия почек на нем образуются отличимые по онтогенетическому состоянию раметы следующих поколений. Наибольшее число растущих корневищных побегов сосредоточено в его апикальной части. Они дают начало ювенильным раметам, которые полностью проходят свой частный онтогенез. Корневищные побеги, возникшие из почек в средней и базальной частях, могут развиваться под землей 2–3 года и, выйдя на поверхность, формировать удлиненный ортотропный годичный генеративный побег. Раскрытие спящих почек на многолетних базальных участках корневища приводит к образованию субсенильных и сенильных рамет. Достаточно часто из-за высокой плотности и числа растущих корневищ в почве, раметы в ювенильном и имматурном состояниях могут сокращать свой онтогенез. Они, минуя генеративное состояние, переходят в постгенеративный период. Вегетативное размножение, связанное с естественным нарушением связи между парциальными побегами и парциальными кустами, может происходить на любом этапе развития полицентрических систем.
Итак, изучение онтогенеза семенной особи S. oreophila выявило, что уже на его ранних этапах почки возобновления за счет контрактильной деятельности главного корня остаются на поверхности почвы или в подстилке. Первичный и парциальные кусты формируются в результате базитонного ветвления анизотропных дициклических удлиненных побегов возобновления. Их второй годичный прирост ежегодно отмирает, а первый, несущий почки возобновления, полегает и укореняется придаточными корнями. Эти признаки, предложенные Л.Е. Гатцук (Gatsuk, 1976) для отнесения растений к травам сезонного климата, характеризуют S. oreophila как травянистое растение. Подробный биоморфологический анализ и особенности онтогенеза особей позволяют решать спорные вопросы жизненных форм видов (Cheryomushkina et al., 2012; 2019; Denisova et al., 2017). Так, у S. supina, родственного вида к S. oreophila, в различных экологических условиях формируются разные жизненные формы (Cheryomushkina, Guseva, 2015). Нами установлено, что длиннокорневищно-стержнекорневая биоморфа S. supina образуется в мезофильных условиях, что в общих чертах сходно c развитием особей S. oreophila. У обоих видов онтогенез особей сложный и состоит из онтогенеза генеты и рамет. Корневищные побеги образуются из спящих почек, что приводит к образованию куртины. Но в отличие от S. oreophila, фаза куртины у семенной особи S. supina длится до завершения онтогенеза, что характеризует жизненную форму этого вида как длиннокорневищно-стержнекорневую. В то время как у особей S. oreophila распад куртины происходит в середине онтогенеза (в средневозрастном генеративном состоянии). Стоит отметить, что в побегообразовании обоих видов участвуют спящие почки, что является одним из механизмов формирования длиннокорневищной жизненной формы (Astashenkov, Cheryomushkina, 2019).
Таким образом, существование длительное время особей S. oreophila в виде явнополицентрической системы парциальных кустов и побегов, связанных между собой длинными симподиально сочлененными гипогеогенными коммуникационными корневищами, их интенсивное ветвление, приводящее к пространственному захвату территории, характеризует жизненную форму изученного вида как длиннокорневищную.
ЗАКЛЮЧЕНИЕ
Изучение развития особей эндемичного вида S. oreophila, растущего на альпийских лугах, показало, что вид относится к явнополицентрическим растениям. Размножение особей вида в основном вегетативное за счет формирования длинных корневищ, которые развиваются из спящих почек. В результате вегетативного размножения образуется клон, состоящий из особи семенного происхождения (генеты), полицентрических систем, включающих парциальные побеги и кусты, а также из отдельных парциальных побегов и парциальных кустов (рамет). Выявлена высокая степень поливариантности сложного онтогенеза, выражающаяся в разной длительности частного онтогенеза рамет и пропусков отдельных онтогенетических состояний.
На основании наших исследований биоморфологический портрет особей S. oreophila можно охарактеризовать как – вегетативно-подвижный травянистый длиннокорневищный симподиально нарастающий многолетний поликарпик. Структура взрослых особей состоит из двух типов побегов: удлиненных и корневищноудлиненных. Удлиненные побеги моно-дициклические анизотропные, иногда ортотропные, вегетативные или генеративные, развертываются из почек зоны возобновления. Корневищноудлиненные побеги – ди-, полициклические вегетативные или генеративные, образуются из спящих почек. Корневая система генеративной особи смешанная, главный корень сохраняется в течение всей жизни особи. Придаточные корни развиваются на геофильных частях дициклических побегов и на корневище.
Список литературы
[Abdurakhmanova, Neshataeva] Абдурахманова З.И., Нешатаева В.Ю. 2017. Классификация сосновых лесов из сосны Коха (Pinus kochiana Klotzsch. ex C. Koch) республики Дагестан. – Бюл. гос. Никитского бот. сада. 123: 43–50.
[Abdurakhmanova et al. ] Абдурахманова З.И., Нешатаев В.Ю., Нешатаева В.Ю. 2018. Сосновые леса (Pineta kochianae) в Республике Дагестан. – Растительность России. 34: 3–46. https://doi.org/10.31111/vegrus/2018.34.3
Astashenkov A.Yu., Cheryomushkina V.A. 2019. The Role of Dormant Buds in Transformation of Herbaceous Biomorphs (by the Example of Nepeta L. sect. Spicatae (Benth.) Pojark.). – Biol. Bulletin. 46 (6): 562–569. https://doi.org/10.1134/S1062359019040046
[Cheryomushkina et al.]. Черемушкина В.А., Асташенков А.Ю., Бобоев М.Т. 2012. Морфогенез и состояние ценопопуляции Limonium chodshamumynense Lincz. et Czuk. в южном Таджикистане (г. Ходжа-Мумин). – Известия АН Республики Таджикистан. Отд. биол. и мед. наук. 1: 7–13.
Cheryomushkina V.A., Guseva A.A. 2015. Life forms of Scutellaria supina L. (Lamiaceae). – Contemp. Probl. Ecol. 8 (5): 624–635.
[Cheryomushkina et al.] Черёмушкина В.А., Таловская Е.Б., Асташенков А.Ю., Гусева А.А., Джуманов С. 2019. Биология Thymus dmitrievae Gamajun. (Lamiaceae) на заповедной территории (Заповедник Аксу-Жабаглы). – Вестн. Том. гос. ун-та. Биология. 47: 103–122. https://doi.org/10.17223/19988591/47/6
[Galushko] Галушко А.И. 1980. Флора Северного Кавказа. Определитель. Т. 3. Ростов-на-Дону. 328 с.
[Gatsuk] Гатцук Л.Е. 1974. К методам описания и определения жизненных форм в сезонном климате. – Бюл. МОИП. Отд. биол. 79 (3): 84–100.
[Gatsuk] Гатцук Л.Е. 1976. Содержание понятия “травы” и проблема их эволюционного положения. – Труды Московского общества испытателей природы. 42: 55–130.
[Grossheim] Гроссгейм А.А. 1949. Определитель растений Кавказа. Л. 746 с.
[Grossheim] Гроссгейм А.А. 1967. Род Scutellaria L. – Шлемник. – В кн.: Флора Кавказа. Т. 7. Л. С. 308–322.
[Denisova et al.] Денисова Г.Р., Черемушкина В.А., Асташенков А.Ю., Гусева А.А. 2017. Особенности развития Dracocephalum fragile Turcz. ex Benth. (Lamiaceae) в Туве. – Вест. ТвГУ. Серия: Биология и экология. 4: 144–157.
[Isaev] Исаев Я.М. 1957. Род Scutellaria L – Акулька. – В кн.: Флора Азербайджана. Т. 7. Баку. С. 232–245.
[Ivanov] Иванов А.Л. 1998. Флора Предкавказья и ее генезис. Ставрополь. 162 с.
[Ivanov] Иванов А.Л. 2019. Конспект флоры Российского Кавказа (сосудистые растения). Ставрополь. 323 с.
[Karpun] Карпун Ю.Н. 2002. Дикорастущие древесные растения Северного Кавказа. Сочи. 14 с.
[Menitsky] Менитский Ю.Л. 1992. Конспект видов семейства Lamiaceae (Labiatae) флоры Кавказа. – Бот. журн. 77 3 (6): 63–78.
[Murtazaliev] Муртазалиев Р.А. 2009. Конспект флоры Дагестана (Melantaceae – Acoraceae). Т. 3. Махачкала. 304 с.
[Nukhimovskiy] Нухимовский Е.Л. 1997. Основы биоморфологии семенных растений. Т. 1. М. 630 с.
[Portenier] Портениер Н.Н. 2000. Система географических элементов флоры Кавказа. – Бот. журн. 85 (9): 26–33.
[Rabotnov] Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах. – Тр. БИН АН СССР. Сер. 3. Геоботаника. 6: 179–196.
Savinykh N.P., Cheryomushkina V.A. 2015. Biomorphology: Current Status and Prospects. – Cont. Prob. Ecol. 8 (5): 541–549.
[Serebryakov] Серебряков И.Г. 1964. Жизненные формы высших растений и их изучение. – В кн.: Полевая геоботаника. Т. 3. Л. С. 146–208.
[Serebryakov] Серебряков И.Г. 1959. Типы развития побегов у травянистых многолетников и факторы их формирования. – Учен. Зап. Моск. гор. пед. ин-та им. В.П. Потемкина. Вопросы биологии растений. 100 (5): 3–38.
[Smirnova et al.] Смирнова О.В., Заугольнова Л.Б., Торопова Н.А., Фаликов Л.Д. 1976. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений различных биоморф. – В кн.: Ценопопуляции растений (основные понятия и структура). М. 215 с.
[Tsepkova] Цепкова Н.Л. 2012. Новая ассоциация остепненных лугов Джинальского хребта. – Известия Самарского научного центра РАН. 14 (1–4): 1149–1151.
[Uranov] Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов. – Биол. науки. 2: 7–34.
[Yuzepchuk] Юзепчук С.В. 1954. Род Шлемник – Scu-tellaria L. – В кн.: Флора СССР. Т. 20. М.–Л. С. 72–225.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал