

Ботанический журнал, 2021, T. 106, № 3, стр. 211-228
БОЛОТА ТАЕЖНОГО СЕВЕРО-ВОСТОКА ЕВРОПЕЙСКОЙ РОССИИ
Т. К. Юрковская *
Ботанический институт им. В.Л. Комарова РАН
197360 Санкт-Петербург, ул. Проф. Попова, 2, Россия
* E-mail: yurkovskayatat@gmail.com
Поступила в редакцию 05.08.2020
После доработки 09.11.2020
Принята к публикации 10.11.2020
Аннотация
Растительность болот на северо-востоке тайги Европейской России по площади уступает только лесам. Преобладают крупные болота и болотные системы, а среди них – верховые и аапа. Наши исследования разрушили сложившиеся представления о существовании печорской провинции аапа болот. На самом деле они распространены в пределах всей северной и средней тайги, а также в лесотундре. Соотношение верховых и аапа болот на севере тайги можно уподобить соотношению на этом пространстве еловых и сосновых лесов. На западе, в Кольско-Карельском регионе, сосновые леса преобладают над еловыми, а на северо-востоке господствуют еловые леса, сосняки уступают им по площади и запасам. Аналогично на западе аапа болота доминируют, верховые значительно им уступают. На северо-востоке верховые болота преобладают над аапа.
Рассматриваемая часть северо-востока европейской тайги России занимает северную и среднюю подзоны тайги в пределах восточноевропейского ботанико-географического сектора. Заметим, что между лесотундрой и северной тайгой протянулась еще одна широтная полоса – крайнесеверная тайга. Но в данной работе мы ее не рассматриваем в качестве особой подзональной категории, а объединяем с северной тайгой. Это связано с тем, что в отношении ее трактовки современными геоботаниками нет единства. В Архангельской области, несмотря на то что она была впервые выделена именно там А.П. Шенниковым (Shennikov, 1941), до сих пор ее границы не установлены. В республике Коми, по предложению Ю.П. Юдина (Yudin, 1954) – ученика и последователя А.П. Шенникова, все геоботаники выделяют подзону крайнесеверной тайги (Degteva et al., 2001). Исследователи Карелии рассматривают ее в качестве широтной полосы второго порядка и называют полосой редкостойной тайги, следуя М.Л. Раменской (Ramenskaya, Shubin, 1975) или крайнесеверной (Yurkovskaya, Payans-kaya-Gvozdeva, 1993).
Восточноевропейский ботанико-географический сектор хорошо отделяется от граничащего с ним западнорусского сектора. Границей между ними служит река Онега, впадающая в Белое море. Онега разделяет Восточную Фенноскандию (Кольско-Карельский регион) и таежный Север Восточно-Европейской равнины. На востоке граница проходит по Приуралью. Эти меридиональные сектора были показаны мною на карте (Yurkovskaya, 2010, 2011). В географических координатах граница рассматриваемого региона идет от 66° с.ш. до 60° с.ш. и от 39°в.д. до 59° в.д.
На западе, в Карелии, доминируют сосновые леса, а в нашем регионе – еловые. Larix sibirica11 впервые появляется в Кольско-Карельском регионе (Kozhin, Sennikov, 2016), но только на северо-востоке елово-лиственничные, сосново-лиственничные и лиственничные леса начинают встречаться довольно часто (Kucherov, Zverev, 2011; Kucherov, 2019). В Архангельской области, в верховьях реки Вага, левого притока Северной Двины, появляется Abies sibirica22 (по наблюдениям автора и Т.И. Исаченко в 1969 г). Западная граница пихты в регионе очень хорошо и четко описана во флоре Северо-Востока (Flora…, 1974). Леса с примесью пихты и пихтовые леса встречаются уже в Республике Коми. На Тиманском кряже (Республика Коми) отмечен Pinus sibirica, а затем уже в Приуралье и на Урале (Nepomilueva, 1974).
Везде в этой части северной и средней тайги много болот. Но в Карелии господствуют болота карельского аапа типа, грядово-мочажинные сфагновые верховые болота встречаются редко, как и болота других типов, тогда как на северо-востоке преобладают верховые болота, а аапа встречаются спорадически и лишь местами доминируют.
Общими для обоих секторов являются субокеанические дистрофные верховые болота южноприбеломорского типа, распространенные вдоль юго-восточного побережья Белого моря и далее вплоть до реки Мезень. Эти болота не определяют специфику секторов, к тому же о них имеется обширная литература (Katz, 1937, 1948; Bogda-novskaya-Guienéuf, 1949; Elina, Yurkovskaya, 1965; Elina, 1969, 1971; Yurkovskaya, 2003, 2004). Поэтому в данной статье я их не обсуждаю.
Итак, на таежном северо-востоке доминируют верховые и аапа болотные массивы. Классики русского болотоведения (Zinserling, 1929, 1938; Katz, 1948, 1971; Bogdanovskaya-Guienéuf, 1949) отнесли их к двум особым географическим типам: онежско-печорский верховой тип болотных массивов и печорский аапа тип. Мною было показано, что на самом деле у них общий ареал, простирающийся от реки Онега на западе до Урала (Yurkovskaya, 1992, 2004, 2010). Исследования верховых и аапа болот продолжаются в этом регионе и подтверждают общность ареалов обоих типов, а также уточняют границу между секторами и выявляют варианты этих типов (Kutenkov, 2006; Smagin et al., 2013; Goncharova, 2013, 2014 и др.). И в дальнейшем мы их назовем: северовосточноевропейские верховые континентальные (в отличие от упомянутых выше субокеанических южноприбеломорских) и северовосточноевропейские аапа. Переименование связано с тем, что во время работы над картой Европы мы столкнулись не только с трудностями перевода на английский язык, но и понимания наших географических терминов зарубежными коллегами.
Кассандрово-морошково-сфагновые со сфагновыми мочажинами на склонах и с заболоченным лесом по краю северовосточноевропейские континентальные верховые болота.
До конца 1970-х годов имелись лишь краткие геоботанические описания нескольких болотных массивов (Rabotnov 1929, 1930; Katz, Minkina 1936; Korchagin, 1940).
Опубликованы данные по стратиграфии торфяных залежей и районированию отдельных частей региона (Nikonov, 1958а, b; Kiryushkin, 1967; Abramova, Kiryushkin, 1968). Исследования активизировались в связи с составлением геоботанической карты европейской части СССР (Karta…, 1979). С этой целью мною проводились геоботанические исследования болот в Архангельской, Вологодской областях и республике Коми, в результате которых был собран материал по флоре, растительности, строению торфяной залежи, а также возрасту верховых болот этого типа (Yurkovskaya 1984, 1992; Elina, Yurkovskaya, 1980). Кроме того, мною были отдешифрированы материалы аэрофотосъемки на эту территорию. В дальнейшем исследования были продолжены как мною, так и ботаниками Петрозаводска, Сыктывкара, Архангельска и др. На сегодня мы имеем довольно полную картину о составе и структуре растительного покрова верховых болот этого типа, их морфологии, строении торфяной залежи и возрасте. Кроме того, теперь доступны материалы космической съемки, которые значительно облегчили процесс распознавания типов на отдельных полигонах и экстраполяции результатов исследования.
Верховые болота этого типа имеют слабовыпуклую поверхность. Превышение вершин над окраинами составляет 2–3 м. Многие массивы имеют асимметричную форму поверхности, т.е. их вершина и самая глубокая часть залежи сдвинуты к одному краю (эксцентрические). Морфологические части болот различаются нечетко. Вычленяется вершина с мелкомочажинным микрорельефом, поросшая низкорослой сосной, и склон с более или менее хорошо развитым грядово-мочажинным микрорельефом. Грядово-мочажинный комплекс занимает основную часть болотного массива, а на некоторых болотах регрессивный или грядово-озерковый комплекс.
Мною установлено, что степень выраженности того или иного комплекса на болотах этого типа или их отсутствие не связано ни с глубиной залежи, ни с возрастом массива, а является исключительно функцией его водосборной площади, т.е. зависит от размеров массива. Примером зависимости степени выраженности тех или иных комплексов от площади болот служит изучавшийся нами в течение ряда лет полигон, болотная система Себболото (Yurkovskaya, 1992 и др.).
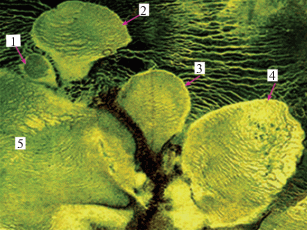
Самый маленький массив (5.4 га) в этой системе весь покрыт сообществами одной ассоциации (Pinus sylvestris f. Litwinowii-Chamaedaphne calyculata-Ledum palustre-Sphagnum fuscum) (рис. 1.1 ). В центре следующего по величине (24 га) массива сформировался грядово-мочажинный комплекс (рис. 1.2 ). Третий (32.5 га) почти весь занимают грядово-мочажинные комплексы, различающиеся размерами гряд и мочажин и их растительным покровом, а у вершины появляются первые признаки деградации и денудации мохового покрова в мочажинах (рис. 1.3 ). Явления деградации и денудации растительного покрова на грядах и в мочажинах очень характерны для четвертого по величине массива (88 га, рис. 1.4 ). Регрессивный комплекс на болоте № 4 вытянулся по профилю на 300 м. Мочажины достигают ширины 20–30 м, имеют неравномерный покров, состоящий из фрагментов разных ассоциаций и пятен обнаженного торфа. На грядах Sphagnum fuscum оттеснен лишайниками к краям. Среди содоминирующих видов на грядах и в мочажинах впервые появляется Trichophorum cespitosum. Регрессивный комплекс занимает подавляющую часть самого крупного болота (638 га). На грядах преобладают сообщества c Empetrum nigrum s.l., Rubus chamaemorus, Sphagnum fuscum и Cladina, редкой сильно искривленной сосной. Мы назвали такую сосну Pinus sylvestris f. pomorica, она очень характерна для регрессивных комплексов верховых болот этого типа и южноприбеломорских, но ботанического описания этой формы нет.
Рис. 1.
Пространственная организация болотной системы Себболото, аэрофотоснимок 64о31ʹ с.ш. 44о17ʹ в.д.
Fig. 1. Spatial organization of the Sebboloto miresystem, aerial view 64°31ʹ N 44°17ʹ E.
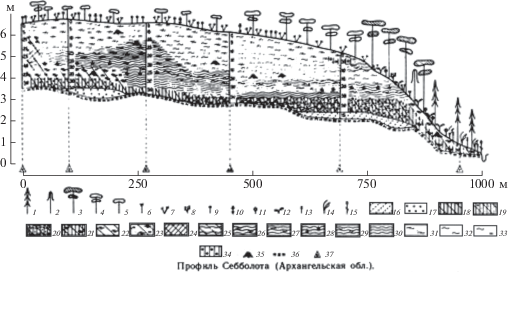
Морфология северовосточноевропейских верховых болот, характер растительности и строение торфяной залежи хорошо прослеживаются на профилях массивов, изученных в Архангельской области и опубликованных мною в 1992 г. (рис. 14–16 ). Глубина залежи на исследованных нами болотах колебалась от 2.5 до 5 м. Залежь верхового типа (рис. 2). Верхний пласт залежи образован комплексом верховых торфов – фускум – под грядами, сфагновым мочажинным – под мочажинами. На вершине, где нет грядово-мочажинного комплекса, – верхний пласт сложен фускум торфом. Ниже залежь образуют сфагново-пушицевые и пушицевые торфы. Несмотря на значительное удаление болот друг от друга (в нашем материале это левобережье Сев. Двины; правобережье Пинеги, Архангельская область; и бассейн реки Вымь, Республика Коми), строение залежи однотипно. Различается лишь мощность пластов, что связано с глубиной залежи и только в придонных слоях наблюдаются существенные различия. Это свидетельствует о несхожести начальных этапов развития болот. На одних болотах более или менее длительной была озерная фаза развития, они подстилаются сапропелем. Другие сформировались в мелководных бассейнах, об этом говорит гипновый низинный торф, приуроченный к самым глубоким участкам дна, а основная их часть сформировалась путем заболачивания суши, в таком случае на дне лежат древесные и древесно-пушицевые переходные торфы (Yurkovs-kaya, Elina, 1992). За исключением начальных этапов, развитие болот в дальнейшем шло по одному пути.
Рис. 2.
Профиль верхового болота. 1 – Picea obovata, 2 – Betula pubescens, 3 – Pinus sylvestris f. uliginosa, 4 – Pinus sylvestris f. litwinowii, 5 – Pinus sylvestris f. willkommii, 6 – Ledum palustre, 7 – Chamaedaphne calyculata, 8 – Betula nana, 9 – Eriophorum vaginatum, 10 – Scheuchzeria palustris, 11 – Rubus chamaemorus, 12 – Vaccinium myrtillus, 13 – Equisetum fluviatile, 14 – Calamagrostis canescens, 15 – Phalaroides arundinacea; низинные торфы: 16 – гипновый, 17 – хвощовый; переходные торфы: 18 – древесный, 19 – березовый, 20 – березово-пушицевый, 21 – древесно-сфагновый, 22 – сфагновый, 23 – шейхцериево-сфагновый, 24 – осоковый, 25 – пушицевый; верховые торфы: 26 – пушицево-сфагновый (Sphagnum fuscum), 27 – пушицево-сфагновый (S. magellanicum), 28 – пушицево-сфагновый (S. angustifolium), 29 – пушицево-сфагновый (мочажинный), 30 – пушицевый, 31 – фускум, 32 – сфагновый мочажинный, 33 – шейхцериево-сфагновый пушицево-сфагновый (мочажинный), 34 – сосновый, 35 – пень, 36 – глина, 37 – номера скважин.
Fig. 2. Profile of the raised bog. 1 – Picea obovata, 2 – Betula pubescens, 3 – Pinus sylvestris f. uliginosa, 4 – Pinus sylvestris f. litwinowii, 5 – Pinus sylvestris f. willkommii, 6 – Ledum palustre, 7 – Chamaedaphne calyculata, 8 – Betula nana, 9 – Eriophorum vaginatum, 10 – Scheuchzeria palustris, 11 – Rubus chamaemorus, 12 – Vaccinium myrtillus, 13 – Equisetum fluviatile, 14 – Calamagrostis canescens, 15 – Phalaroides arundinacea; eutrophic peat types: 16 – brown moss peat, 17 – Equiseteti peat; mesotrophic peat types: 18 – woody peat, 19 – Betuleti peat, 20 – Betulo-Eriophoreti peat, 21 – woody-Sphagnum moss peat, 22 – Sphagneti peat, 23 – Scheuchzerio-Sphagneti peat, 24 – Cariceti peat, 25 – Eriophoreti peat; oligotrophic peat types: 26 – Eriophoro-Sphagneti fuscum peat, 27 – Eriophoro-Sphagneti magellanicum peat, 28 – Eriophoro-Sphagneti angustifolium peat, 29 – Eriophoro-Sphagneti hollow peat, 30 – Eriophoreti peat, 31 – Sphagneti fuscum peat, 32 – Sphagneti hollow peat, 33 – Scheuchzerio-Sphagneti and Eriophoro-Sphagneti hollow peat, 34 – Pineti peat; other: 35 – stump, 36 – clay, 37 – well numbers.
Рассмотрим детальнее растительность комплексов, характерных для северовосточноевропейских верховых болот. Самыми распространенными являются грядово-мочажинные комплексы, на многих болотах занимающие почти всю поверхность болота, за исключением самой вершины и окраины. Эти комплексы довольно однообразные. Площадь мочажин обычно несколько превышает площадь гряд. Только у самой вершины обратное соотношение. Размеры мочажин от вершины болота к окраине постепенно увеличиваются. Гряды невысокие (0.3–0.4 м) и широкие (3–5 м). В их моховом покрове господствует Sphagnum fuscum, отличающийся не только постоянством, но и высоким покрытием. Наиболее частыми являются асс. Pinus sylvestris–Chamaedaphne calyculata–Sphagnum fuscum (табл. 1, 1), свойственная всему ареалу этих болот, и Pinus sylvestris–Chamaedaphne calyculata–Empetrum nigrum–Sphagnum fuscum (табл. 1, 2), встречающаяся в основном в северной части их ареала. Наличие сосны и ее значительное участие в растительном покрове не только гряд, но и вершины болота – также характерный признак болот этого типа. Они существенно отличаются этим от болот западнорусского, южноприбеломорского типов, на которых в центральных частях сосен значительно меньше и они ниже. Здесь обычны не только Pinus sylvestris f. willkommii, но и f. litwinowii, достигающая высоты 3.5–4 м. Часто много еще более высокого сухостоя.
Таблица 1.
Значения классов константности видов ассоциаций Sphagneta fusci северовосточноевропейских верховых болот Table 1. Values of species constancy classes in Sphagneta fusci associations of northeastern European raised bogs
| Вид Species | Aссоциации/Associations | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| Число описаний Number of releves | |||
| 11 | 20 | 18 | |
| Empetrum nigrum | r | V | V |
| Vaccinium myrtillus | 0 | II | II |
| V. uliginosum | II | V | V |
| V. vitis-idaea | 0 | II | II |
| Drosera rotundifolia | I | V | V |
| Dicranum affine | III | III | IV |
| Pleurozium schreberi | r | III | III |
| Mylia anomala | II | IV | IV |
| Cladina rangiferina | II | 3-IV | 15-V |
| Sphagnum nemoreum | 0 | III | V |
| Cetraria islandica | 0 | II | V |
| Cladina stellaris | 0 | III | V |
| Trichophorum cespitosum | 0 | r | IV |
| Sphagnum angustifolium | III | III | V |
| S. balticum | II | II | IV |
| S. lindbergii | 0 | I | III |
| Oxycoccus palustris | IV | IV | II |
| Pinus sylvestris | V | V | V |
| Andromeda polifolia | V | V | V |
| Betula nana | II | II | II |
| Chamaedaphne calyculata | V | V | V |
| Ledum palustre | V | V | V |
| Oxycoccus microcarpa | V | V | V |
| Rubus chamaemorus | V | V | V |
| Carex pauciflora | II | II | II |
| Eriophorum vaginatum | V | V | V |
| Sphagnum fuscum | V | V | V |
| Polytrichum strictum | II | II | I |
| Cladina arbuscula | II | III | III |
| Среднее число видов Average number of species | 13 | 18 | 22 |
Таблица 2.
Значения классов константности видов ассоциаций Sphagneta papillosi северовосточноевропейских аапа болот Table 2. Values of species constancy classes in Sphagneta papillosi associations of northeastern European aapa mires
| Вид Species | Ассоциации/Associations | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| Число описаний Number of relives | ||||
| 8 | 9 | 12 | 10 | |
| Betula pubescens | II | 0 | 0 | 0 |
| Carex pauciflora | 0 | II | III | II |
| Sphagnum subfulvum | II | 0 | 0 | 0 |
| S. magellanicum | III | V | II | II |
| Trichophorum cespitosum | II | 0 | V | I |
| Eriophorum angustifolium | V | V | II | IV |
| Juncus stygius | I | I | III | I |
| Loeskypnum badius | 0 | 0 | II | 0 |
| Carex chordorrhiza | I | II | III | V |
| Warnstorfia exannulata | 0 | 0 | 0 | I |
| W. fluitans | 0 | 0 | 0 | I |
| Carex lasiocarpa | V | V | V | I |
| Betula nana | V | V | IV | IV |
| Chamaedaphne calyculata | II | II | I | I |
| Equisetum fluviatile | III | III | II | II |
| Sphagnum papillosum | V | V | V | V |
| Pinus sylvestris | II | 0 | I | 0 |
| Andromeda polifolia | V | V | V | V |
| Oxycoccus palustris | V | V | IV | V |
| Trichophorum alpinum | I | 0 | II | II |
| Carex limosa | III | III | II | II |
| C. rostrata | IV | III | III | III |
| Eriophorum vaginatum | 0 | I | I | I |
| Drosera anglica | II | IV | III | II |
| D. rotundifolia | IV | V | IV | IV |
| Menyanthes trifoliate | V | V | V | V |
| Scheuchzeria palustris | III | V | V | III |
| Sphagnum fallax | I | I | I | 0 |
| S. flexuosum | V | III | IV | II |
| S. jensenii | I | II | III | I |
| S. lindbergii | 0 | II | I | 0 |
| S. majus | 0 | I | I | 0 |
| S. russowii | 0 | II | I | I |
| S. subsecundum | 0 | I | 0 | I |
| S. warnstorfii | I | I | 0 | 0 |
| Straminergon stramineum | III | III | III | IV |
| Hepaticae | 0 | 0 | II | II |
| Среднее число видов Average number of species | 14 | 15 | 14 | 13 |
Растительность мочажин более разнообразна, но еще беднее флористически. Наиболее характерны сообщества асс. Eriophorum vaginatum–Sphagnum balticum, Eriophorum vaginatum–Scheuchzeria palustris–Sphagnum balticum, Scheuchzeria palustris–Sphagnum balticum. Каждая из них представлена типичной и субассоциацией с Andromeda polifolia, характерной для менее увлажненных, по сравнению с типичными, местообитаний. Данные ассоциации отличаются друг от друга доминирующими видами травяного покрова. Они представляют собой последовательный сукцессионный ряд и связаны с определенными уровнями болотных вод и размерами мочажин. Первые самые небольшие (не более 2–5 м ширины, УБВ от –5 до –10 см), последние встречаются в мочажинах до 10–15 м шириной, с УБВ от 0 до –3 см. Широко распространены также мочажины со Sphagnnum majus, асс. Eriophorum vaginatum–Scheuchzeria palustris–Sphagnum majus и Scheuchzeria palustris–Sphagnum majus (рис. 3). Последние приурочены к самым крупным мочажинам шириною от 15 до 20 м и длиною до 100–300 м с высоко стоящей водой (–3, 0, +3 см), а на севере региона часто встречаются также мочажины со Sphagnum lindbergii.
Рис. 3.
Мочажина Scheuchzeria palustris–Sphagnum majus.
Fig. 3. Hollow Scheuchzeria palustris–Sphagnum majus.

Значительные площади на крупных болотных массивах в северной тайге занимают регрессивные дистрофные комплексы, для которых характерны гряды с обилием лишайников и большие денудированные мочажины шириною до 20–50 м. Они сосредоточены в верхних участках склонов, а на болотах площадью более 500 га занимают часто всю центральную часть массивов, включая их вершину.
В регрессивном комплексе на грядах преобладают сообщества асс. Pinus sylvestris–Empetrum nigrum–Rubus chamaemorus–Cladina-Sphagnum fuscum (табл. 1, 3). Даже в регрессивном комплексе гряды бывают сильно облесены. Сообщества со значительным участием Pinus sylvestris f. litwinowii характерны для наименее регрессивных участков грядово-мочажинных комплексов. Увеличение низкорослых форм связано с развитием регрессивных комплексов и увеличением размеров массивов. В центре крупных болот с обширными участками регрессивных комплексов гряды почти безлесны.
Таблица 3.
Значения классов константности видов ассоциаций Sphagneta magellanici северовосточноевропейских аапа болот Table 3. Values of species constancy classes in Sphagneta magellanici associations of northeastern European aapa mires
| Вид Species | Aссоциации/Associations | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| Число описаний Number of releves | |||
| 8 | 9 | 12 | |
| Ledum palustre | II | 0 | 0 |
| Drosera rotundifolia | III | V | V |
| Carex rostrata | IV | I | II |
| Sphagnum angustifolium | III | 0 | 0 |
| S. papillosum | I | III | III |
| S. russowii | III | I | 0 |
| Chamaedaphne calyculata | V | 0 | III |
| Trichophorum cespitosum | I | IV | 0 |
| Straminergon stramineum | II | 0 | V |
| Carex pauciflora | II | 0 | V |
| Eriophorum angustifolium | II | I | IV |
| Andromeda polifolia | V | V | V |
| Betula nana | IV | V | V |
| Carex lasiocarpa | IV | III | V |
| Menyanthes trifoliata | V | V | V |
| Warnstorfia exannulata | 0 | 0 | 0 |
| W. fluitans | 0 | 0 | 0 |
| Sphagnum magellanicum | V | V | V |
| Empetrum nigrum | II | 0 | I |
| Oxycoccus microcarpus | I | II | 0 |
| O. palustris | V | V | V |
| Rubus chamaemorus | I | II | I |
| Eriophorum vaginatum | I | II | II |
| Carex chordorrhiza | I | I | I |
| C. limosa | II | II | I |
| Equisetum fluviatile | I | 0 | 0 |
| Scheuchzeria palustris | II | III | II |
| Sphagnum balticum | 0 | I | I |
| S. fallax | I | 0 | I |
| S. flexuosum | II | I | II |
| S. fuscum | II | I | II |
| S. warnstorfii | I | 0 | 0 |
| Pohlia nutans | I | 0 | 0 |
| Polytrichum strictum | II | II | II |
| Среднее число видов Average number of species | 13 | 13 | 13 |
Другая особенность растительного покрова гряд в регрессивном комплексе – появление микропонижений с растениями, чуждыми грядам. Это прежде всего влаголюбивые мхи – Sphagnum balticum, S. lindbergii. Предшественниками их являются участки с синузиями психрофильных мхов (Dicranum affine, Pleurozium schreberi), печеночников (Mylia anomala), лишайниками (Cladonia arbuscula, C. rangiferina), они сменяются упомянутыми сфагнами, которые разрастаются и вместе с сопутствующими им сосудистыми растениями образуют микроценозы или фрагменты фитоценозов, чуждые сообществам гряд. Наблюдались разные стадии формирования таких микроценозов, которые начинались с вымокания и осклизнения лишайников, поселения печеночников, а затем сфагновых мхов. Некоторые из них потом вновь заполнялись лишайниками или Sphagnum fuscum, площадь других увеличивалась, и на многих грядах регрессивного комплекса они занимали до 10% площади, сосредоточиваясь в понижениях размером от 0.75–1 м ширины до 1–3 м длины. Подобные микроценозы уже представляют зачатки будущих мочажин, а их состав по набору видов близок к составу аналогичных мочажин. “Грядовые” виды находятся в малом обилии и угнетенном состоянии. В дальнейшем такие микроценозы отшнуровываются от гряды и присоединяются к рядом расположенной мочажине или гряда распадается на части и формируется новая мочажина. Таким путем происходит увеличение площади мочажин и уменьшение площади гряд.
Денудированные мочажины имеют пестрый, т.е. очень неравномерный и разнородный покров, состоящий из отдельных, часто сменяющихся пятен – фрагментов разных фитоценозов, перемежающихся с участками обнаженного торфа. В их моховом покрове чередуются Sphagnum balticum, S. lindbergii, S. majus, S. papillosum, Cladopodiella fluitans, лишайник Cetrariella delisei, в травяном – Trichophorum cespitosum, Carex limosa, Drosera anglica, Eriophorum vaginatum, Scheuchzeria pa-lustris, а на севере региона местами и Eriophorum russeolum. Иногда каждый из фрагментов занимает лишь небольшое пятно, в некоторых мочажинах преобладают 1–2 фрагмента, а остальные вкраплены в этот фон. Порой вся мочажина – сплошной обнаженный торф, а фрагменты с растительным покровом прижаты к краям и углам ее. Длина пятен обнаженного торфа достигает иногда свыше 100 м при ширине 15–25 м (рис. 4). На севере региона для мочажин характерны фитоценозы асс. Trichophorum cespitosum–Sphagnum lindbergii. Они особенно свойственны молодым небольшим мочажинам, а также крупным денудированным мочажинам, сосредотачиваясь в их углах. На севере часто встречаются и мочажины асс. Eriophorum russeolum–Sphagnum balticum, Eriophorum russeolum–Sphagnum lindbergii, Eriophorum russeolum–Sphagnum majus.

Во флоре этих болот, по неполным данным, насчитывается 55 видов сосудистых растений, 25 видов листостебельных мхов (из них 17 сфагновых), 10 лишайников, 3 Hepaticae. Флора специально не изучалась, особенно ущербны данные о Hepaticae, недостаточно изучены лишайники, нет данных о водорослях, которые свойственны мочажинам в регрессивных комплексах и встречаются в озерках. Виды-эпифиты и эпиксилы игнорировались намеренно, так как они связаны с породой дерева или минерала и идентичны в лесу и на болоте. Более половины всех сосудистых растений (35 видов) характерны для нетипичных местообитаний, в том числе 27 для краевого заболоченного леса, а также топей. В олиготрофных и дистрофных фитоценозах центральной части насчитывается 19 видов сосудистых растений, 16 – листостебельных мхов (из них 12 сфагновых), 5 видов лишайников (Cladonia arbuscula, C. rangiferina, C. stellaris, Cetrariella delisei, C. islandica), все 3 Hepaticae (Mylia anomala, Cladopodiella fluitans, Gymnocolea inflata). Среди трав и кустарничков господствуют Eriophorum vaginatum, Scheuchzeria palustris, Carex limosa (реже), Chamaedaphne calyculata, Rubus chamaemorus, Empetrum nigrum s.l., Andromeda polifolia, Oxycoccus microcarpus, O. palustris. В северотаежной подзоне часто доминирует Trichophorum cespitosum, а к северо-востоку от реки Пинега в мочажинах активной становится Eriophorum russeolum. Следует отметить частое присутствие в олиготрофных центральных частях массивов Carex pauciflora, а местами Melampyrum pratense. В фитоценозах болот, расположенных в бассейне реки Сев. Двина еще встречаются иногда западные виды, характерные для болот субокеанической и северо-западной групп: Sphagnum cuspidatum, S. rubellum, S. tenellum. Несколько раз мною были описаны фитоценозы с Calluna vulgaris, который обычно отсутствует на болотах этого типа.
Широко распространена сосна, представленная на северовосточноевропейских верховых болотах в основном тремя формами: Pinus sylvestris f. litwinowii, P. sylvestris f. willkommii, P. sylvestris f. pomorica. Изучение и описание болотных форм сосны является одной из прерогатив русского болотоведения со времен В.Н. Сукачева и его ученика, талантливого геоботаника Р.И. Аболина (Su-kachev, 1905; Abolin, 1915; Solonevich, 1962; Volkova, 2000). Последняя выделяемая мною форма “pomorica” не была никем ранее описана, так как эти болота до начала наших исследований были изучены очень слабо. Мое же описание чисто физиономическое и требует, безусловно, серьезной корректировки специалистом. Эта экологическая форма сосны характерна для центральных частей крупных болот данного типа, особенно для гряд регрессивного комплекса. Она образуется под влиянием сильных ветров и мороза. При этом отмирает (отмерзает) верхушечный побег, крона становится флагообразной, а ствол перекручивается, приобретая иногда причудливые извилистые формы. Высота ее обычно чуть более 1 м. Она напоминает формы сосны, растущие на южноприбеломорских болотах (Elina, Yurkovskaya, 1965; Elina, 1971). И потому я ей присвоила название pomorica.
Итак, на огромной площади ареала выявлена географическая неоднородность морфологии, структуры и состава растительного покрова, мощности (глубины) торфяной залежи северовосточноевропейских верховых болот. Ранее описанный тип (Katz, 1948) является типичным вариантом этих болот, встречающимся по всему региону и заходящему на восток Карелии, но особенно характерным для южной полосы. Для этого варианта отмечаем безраздельное господство грядово-мочажинных комплексов со Sphagnum fuscum и Chamaedaphne calyculata на грядах, Sphagnum balticum и S. majus в мочажинах. Северотаежный вариант этих болот представлен крупными массивами с обилием лишайников на грядах, а в мочажинах наряду со Sphagnum balticum и S. majus заметным доминантом становится S. lindbergii. Существенное значение на севере приобретает в покрове гряд Empetrum nigrum s.l., а Trichophorum cespitosum и на грядах, и в мочажинах.
Травяно-сфагново-гипновые с грядово-мочажинно-озерковыми комплексами в вогнутой части северовосточноевропейские аапа болота
Напомню главные особенности аапа болот, отличающие их от верховых: 1. вогнутая форма поверхности (у верховых выпуклая); 2. наличие гетеротрофных грядово-мочажинных или грядово-озерковых комплексов в вогнутой части болотного массива (у верховых они располагаются на вершине и склонах, т.е. выпуклых частях болота и гомотрофны по своей структуре); 3. торфяная залежь в центре болота низинного или переходного типа, а по периферии смешанного (у верховых – преобладает верховой тип залежи, только по краю могут быть переходные и низинные); 4. водно-минеральное питание складывается за счет атмосферных, делювиальных и подземных вод (у верховых – только атмосферное); 5. в отличие от верховых в растительном покрове аапа три доминантных синузии – травяная, сфагновая и гипновая (у верховых доминируют сфагновые мхи).
Подчеркнем также значительную проточность вод в связи с сильным уклоном болота по траверзу, впервые это отчетливо было показано О.Л. Кузнецовым (Kuznetsov, 2017). Главное, что обуславливает резкое различие состава и структуры растительного покрова аапа и верховых болот – это различие водного режима. Это было блестяще показано в работе финского исследователя Т. Tohvanainen (2011). Многие особенности структуры и развития аапа болот выявлены Кузнецовым в Карелии (Kuznetsov, 1980, 1982; Kuznetsov, Yurkovskaya, 2016).
Анализ и дешифрирование космических снимков, как и исследования последней четверти ХХ века и начала ХХI-го окончательно подтвердили утверждение о панбореальном распространении аапа болот, высказанное a priori Rauno Ruuhijarvi еще в 1960 г. (Ruuhijarvi, 1960). Нанесены на карту местонахождения аапа болот в Канаде (Zoltai, Pollett, 1983; Wells, Zoltai, 1985). Они обнаружены на севере США, в штатах Мен и Миннесота (Hofstetter, 1983; Davis, Anderson, 2001 и др.). Они показаны мною на картах растительности России и Европы. Первые данные об аапа болотах в Сибири стали появляться с 60-х годов (Storozheva, 1960; Romanova, Usova, 1969; Preys, 1978). В России они распространены в лесотундре, северной и средней тайге; и от западной границы России до Тихого океана (Yurkovskaya, 2010, 2011).
Долгое время, с тех пор как их впервые описали Н.Я. Кац (Katz, 1928) и Ю.Д. Цинзерлинг (Zinserling, 1929), бассейн средней Печоры считался изолированным местонахождением аапа болот этого типа. И только с началом исследований в связи с составлением Карты растительности европейской части СССР (Karta…, 1979) и просмотром аэрофотоснимков мною была установлена сплошная полоса распространения аапа болот на протяжении всего таежного северо-востока, смыкающаяся на западе с карельскими аапа болотами (Yurkovskaya, 1977). Первоначально граница между карельскими и северовосточноевропейскими аапа болотами проводилась мною по реке Выг и Онежскому озеру, но позднее исследования карельских болотоведов выяснили (Kutenkov, 2006), что эту границу следует проводить восточнее. Одним словом, как было сказано выше, ареал верховых и аапа болот на северо-востоке Европы совпадает, только в отличие от Карелии аапа болота на северо-востоке уступают верховым, хотя в некоторых местах, а не только по Средней Печоре, они занимают значительные площади. В целом же можно сказать, что соотношение верховых и аапа болот аналогично соотношению еловых и сосновых лесов в тайге и зависит это в основном от геоморфологии и водного питания болот.
Исследования в Республике Коми проводила Р.Н. Алексеева (Alekseeva, 1974, 1988), а позднее вплоть до настоящего времени ведет Н.Н. Гончарова (Goncharova, 2014). Мною с начала 70-х годов велись исследования аапа болот в Архангельской области и Республике Коми, а в настоящее время под руководством О.Л. Кузнецова продолжают их изучение в Архангельской области карельские и местные специалисты (Kuznetsov et al., 2013).
Аапа болота занимают проточные котловины, но нередко, особенно в Архангельской области, они встречаются в сочетании с верховыми внутри болотных систем, подавляющую площадь которых занимают верховые. В этих системах чаще всего они располагаются на месте транзитных топей. При этом на юге ареала аапа болот (вплоть до севера южной тайги) сохраняются лишь те из них, которые входят в состав болотных систем. Южнее севера южной тайги по всей европейской тайге они отсутствуют, а проточные топи в системах приобретают свой классический характер, так называемые гладкие транзитные топи, т.е. без грядово-мочажинного комплекса с евтрофной травяной, травяно-моховой или мезотрофной травяно-сфагновой растительностью. Впрочем, гладкие топи встречаются в системах, на протяжении всего ареала верховых болот, вплоть до их северной границы.
Размеры болотных массивов сильно варьируют, но много крупных болот площадью несколько тысяч гектаров (Nikonov, 1953; Alekseeva, 1974; Goncharova, 2014). Но более часто встречаются и по количеству преобладают аапа болота небольших размеров.
По макро- и микроморфологии они очень сходны с аапа болотами Карелии; также имеют вогнутую поверхность и уклон по продольному траверзу. В вогнутой части массивов развит грядово-мочажинный микрорельеф, а по окраинам кочковатый или окраины ровные. Наряду с грядово-мочажинным и грядово-озерковым комплексами, характерными для аапа болот Карелии, широко распространены грядово-озерково-топяные. Любопытно, что на некоторых болотах гряды и мочажины имеют концентрическую форму и физиономически напоминают расположение гряд и мочажин на верховых болотах. Это впервые было отмечено Н.Н. Гончаровой на космоснимке болота Пурганюр, а впоследствии этот болотный массив был изучен ею в натуре (Goncharova, Dubrovskiy, 2014).
В заключение повторим следующее: аапа болота имеют панбореальный ареал и в России они встречаются от ее западных границ до восточных.
Залежь этих аапа болот неглубокая (2–4 м), максимум 6 м (Nikonov, 1953; Yurkovskaya, 1984). Они отличаются большим разнообразием слагающих торфов. Характерны залежи переходного и низинного типа, топяного и топяно-лесного вида строения. Нередко и в низинном типе залежи верхний слой образован переходными торфами.
Растительный покров пестр и разнообразен. Преобладают многочисленные грядово-мочажинно-озерковые и озерково-топяные комплексы. На положительных элементах микрорельефа, как на всех типах аапа, господствуют сфагновые сообщества, а к отрицательным приурочены травяные и гипновые. Но в отличие от карельских аапа болот доля участия в мочажинах травяно-гипновых сообществ значительно сократилась, а роль сфагновых сообществ увеличилась.
Сообщества формации Sphagneta papillosi являются самыми распространенными, формируя покров гряд, кочек, ковров (табл. 2). Высота гряд варьирует от 20 до 40 см, но в основном равна 25 см, а высота кочек всего 10–15 см, среди них много крупных плоских кочек-подушек.
УБВ в межень под грядами составляет 10–20 см ниже поверхности мохового покрова, а под кочками и в сообществах ковра – –5...–10 см. Эдификаторная роль Sphagnum papillosum хорошо выражена, он редко уступает под натиском травяно-кустарничкового покрова. Местами усиливается роль более влаголюбивых видов секции Cuspidata, реже более евтрофных (Sphagnum subfulvum, S. warnstorfii), которые вообще редко отмечают на болотах этого типа, иногда усиливается роль Sphagnum magellanicum и S. russowii. Только 3 вида имеют V класс константности: Andromeda polifolia, Oxycoccus palustris и Menyanthes trifoliata. Carex lasiocarpa в сравнении с карельскими встречается значительно реже, но по-прежнему является характерным содоминантом узких извилистых гряд аапа и в ветреные дни колышется словно ковыль в степи. Нет Juniperus, Molinia caerulea, Calluna vulgaris, которые в сообществах ряда ассоциаций карельских аапа болот были содоминантами. Остальные содоминанты те же: Betula nana, Carex lasiocarpa, Trichophorum cespitosum, но добавляются еще Carex chordorrhiza и C. rostrata. Очень широко распространена здесь Eriophorum angustifolium (IV класс константности, тогда как в Карелии в этой формации она относилась к I классу).
Асс. Betula nana–Carex lasiocarpa–Sphagnum papillosum формирует покров гряд. Основную массу травяно-кустарничкового яруса образует Carex lasiocarpa, местами ее покрытие достигает 50–65%. Общее покрытие травяно-кустарничкового яруса колеблется от 25 до 85%, составляя в среднем 50%. Число видов в сообществах варьирует от 12 до 18 (табл. 2, 1).
Асс. Сarex lasiocarpa–Sphagnum papillosum (табл. 2, 2) также образует покров гряд и ковров. Но гряды ниже, от 10–15 до 20 см. Покрытие и обилие Carex lasiocarpа ниже, чем в предыдущей ассоциации, а Betula nana хотя и присутствует во всех сообществах, но совсем теряет свою доминирующую роль, встречаясь рассеянно или единично. Значительно ниже и общее покрытие травяно-кустарничкового яруса, оно изменяется от 10 до 40%, а в среднем около 20%. По составу ассоциация близка к предыдущей, но увеличилось присутствие более влаголюбивых и светолюбивых видов, например Carex chordorrhiza, Drosera anglica, Scheuchzeria palustris, Sphagnum magellanicum.
Асс. Trichophorum cespitosum–Sphagnum papillosum является самой распространенной (табл. 2, 3). Ее фитоценозы характерны для узких низких гряд, встречаются на плоских кочках-подушках и образуют ковры. Высота гряд и кочек 10–20 см. Ширина гряд от 0.5 до 4 м, но в основном не шире 1–1.2 м. УБВ колеблется от –5 до –15 см. Можно выделить субассоциацию с Betula nana, которая характерна для более высоких и широких гряд. Обилие и покрытие Trichophorum варьирует от 10 до 70%; во многих ценозах разрастается Andromeda polifolia, покрытие которой местами достигает 20%. Растения более высокие, как Carex lasiocarpа, C. rostrata, Eriophorum angustifolium, встречаются единично или рассеянно и яруса не образуют. Trichophorum cespitosum формирует собственный ярус высотой 20–30 см, где постоянно присутствует Scheuchzeria palustris. Данная ассоциация отличается от предыдущих не только доминантами травяно-кустарничкового яруса, но и значениями константности ряда видов. Только в этой ассоциации встретился Loeskypnum badium, характерный для сообществ с повышенной трофностью. Выше средней константность у Juncus stygius.
Асс. Carex chordorrhiza–Sphagnum papillosum по принципу выделения некое исключение, так как я ее называю по дифференцирующему виду, встречающемуся с высоким постоянством, но не всегда с высоким обилием и доминирующая роль его сомнительна (табл. 2, 4). Во всех сообществах этой ассоциации хорошо выражен ярус, образованный Menyanthes trifoliata. В этой ассоциации можно выделить 3 субассоциации: типичную, с Carex rostrata и с Carex lasiocarpa.
Таблица 4.
Значения классов константности видов ассоциаций Sphagneta jensenii северовосточноевропейских аапа болот Table 4. Values of species constancy classes in Sphagneta jensenii associations of northeastern European aapa mires
| Вид Species | Aссоциации/Associations | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| Число описаний Number of relives | ||||
| 10 | 10 | 10 | 11 | |
| Eriophorum russeolum | V | 0 | I | II |
| Scheuchzeria palustris | II | V | III | III |
| Carex limosa | V | V | V | IV |
| C. rostrata | II | II | I | V |
| Drosera anglica | III | II | IV | 0 |
| Sphagnum fallax | I | 0 | I | III |
| S. lindbergii | I | I | I | III |
| Oxycoccus palustris | I | I | I | V |
| Andromeda polifolia | I | II | II | II |
| Chamaedaphne calyculata | I | 0 | 0 | I |
| Carex chordorrhiza | I | I | I | 0 |
| Carex lasiocarpa | I | 0 | I | I |
| Eriophorum angustifolium | II | II | II | II |
| E. vaginatum | II | 0 | 0 | II |
| Drosera rotundifolia | I | II | I | I |
| Equisetum fluviatile | 0 | II | I | 0 |
| Menyanthes trifoliata | V | V | IV | V |
| Utricularia intermedia | I | I | I | 0 |
| Sphagnum flexuosum | II | III | I | III |
| S. jensenii | V | V | V | V |
| S. majus | 0 | I | I | II |
| S. obtusum | I | I | I | I |
| S. papillosum | 0 | II | II | 0 |
| S. platyphyllum | 0 | I | I | 0 |
| Straminergon stramineum | I | 0 | II | I |
| Warnstorfia exaтnulata | 0 | I | I | 0 |
| W. fluitans | 0 | II | II | I |
| Среднее число видов Average number of species | 14 | 15 | 14 | 13 |
В процессе сукцессии типичная субассоциация Carex chordorrhiz–Sphagnum papillosum является начальной стадией образования травяно-сфагновых сообществ с господством Sphagnum papillosum. Она занимает небольшие плоские кочки. В ее покрове, помимо Carex chordorrhiza, велика роль других травянистых видов, характерных для мочажин. Субассоциации с Carex rostrata и с Carex lasiocarpa указывают путь от плоских кочек к грядам. При этом гряды с Carex rostrata более типичны для периферийной части грядово-озерково-мочажинного комплекса и чаще сочетаются со сфагновыми мочажинами, а гряды с Carex lasiocarpa – для центральных частей этого комплекса и входят в состав комплексов с озерками и травяными мочажинами.
Наиболее продвинутыми в сукцессионном ряду являются 2 первые ассоциации, они особенно характерны для гряд, составляющих основу грядово-мочажинно-озеркового комплекса. Сообщества ассоциации с Trichophorum cespitosum характерны для узких молодых гряд, они нередко располагаются поперек основного направления гряд и, по-видимому, возникают в результате изменения гидрологического режима в процессе развития болота. Фитоценозы Carex lasiocarpa–Sphagnum papillosum занимают более высокие и широкие гряды, направленные поперек основного стока и часто перпендикулярно грядам с Trichophorum cespitosum. Я предполагаю, что гряды с Trichophorum cespitosum стали развиваться значительно позднее, чем гряды с Carex lasiocarpa.
Сообщества Sphagneta fusci на основной части ареала встречаются значительно реже чем в Карелии. Они чаще всего встречаются на кочках посреди гряд со Sphagnum papillosum или образуют самостоятельные более крупные кочки, иногда собранные в грядообразные цепочки, изредка формируют покров гряд. Обычно в комплексах, где имеются сообщества со Sphagnum fuscum, сосуществуют и преобладают по площади сообщества со Sphagnum papillosum и S. magellanicum, которые также приурочены к положительным элементам микрорельефа. Но господствуют во всех комплексах травяные или травяно-сфагновые сообщества топяных местообитаний или вторичные озерки, почти лишенные растений. Высота кочек и гряд колеблется от 35 до 60 см. УБВ стоит на 25–30 см ниже поверхности мохового покрова.
Всего выделено 4 ассоциации: Empetrum nigrum–Menyanthes trifoliata–Sphagnum fuscum, Chamaedaphne calyculata–Empetrum nigrum–Menyanthes trifoliata–Sphagnum fuscum, Andromeda polifolia–Menyanthes trifoliata–Sphagnum fuscum, Chamaedaphne calyculata–Menyanthes trifoliata–Sphagnum fuscum.
Но к северу роль фитоценозов со Sphagnum fuscum в растительном покрове возрастает, а самое интересное сообщает об этих сообществах Н.Н. Гончарова, которая много работала на аапа болотах в республике Коми в полосе крайнесеверной тайги (Goncharova, 2014). На этих болотах наряду с минеротрофными мочажинами, вторичными озерками и мочажинами-римпи часты очень длинные и высокие гряды со Sphagnum fuscum, на которых отсутствуют минеротрофные виды, их растительность абсолютно аналогична растительности гряд верховых болот.
Фитоценозы формации Sphagneta magellanici на северовосточноевропейских аапа болотах занимают второе место среди сфагновых сообществ, характерных для положительных форм микрорельефа, т.е. то, которое на карельских аапа болотах отведено ценозам Sphagneta warnstorfii. Данная совокупность фитоценозов должна быть выделена в минеротрофную субформацию, так как около 10 видов, характерных для них, вообще не встречаются в олиготрофных фитоценозах Sphagneta magellanici. Это Carex chordorrhiza, C. lasiocarpa, C. limosa, C. rostrata, Equisetum heleocharis, Eriophorum angustifolium, Menyanthes trifoliata, Scheuchzeria palustris, Sphagnum warnstorfii, Straminergon stramineum (табл. 3). Некоторые из этих видов являются субдоминантами: Menyanthes trifoliata, Carex lasiocarpa, C. rostrata.
Сообщества ассоциации Chamaedaphne calyculata–Menyanthes trifoliata–Sphagnum magellanicum встречаются на кочках и грядах аапа болот (табл. 3, 1). Они особенно свойственны для окраинного кочковато-равнинного комплекса и для периферической части центрального грядово-мочажинного комплекса. В последнем они сочетаются с травяно-сфагновыми мочажинами со Sphagnum jensenii, Eriophorum russeolum, Carex limosa, Scheuchzeria palustris. Высота кочек и гряд колеблется в пределах 0.3–0.5 м. УБВ от –15 до ‒35 см.
Высота кустарничкового яруса 20–35 см, а Menyanthes образует приземистый ярус высотой 5–10 см. Высокие травы и Betula nana соcтавляют разреженный полог высотой 50–60 см, изредка они формируют более или менее выраженный верхний ярус. Гряды и кочки безлесны. Иногда на окраинах встречается сухостой сосны.
Общее покрытие травяно-кустарничкового покрова сильно меняется – от 10 до 70%, как и у господствующих видов: Chamaedaphne calyculata – от 10 до 60%, Menyanthes – от 10 до 40%. Различаем 2 субассоциации: типичную и с Carex rostrata.
Сообщества ассоциации Andromeda polifolia–Menyanthes trifoliata–Sphagnum magellanicum (табл. 3, 2) располагаются на небольших высоких кочках или грядах, проективное покрытие Andromeda достигает 20–30%.
Асс. Betula nana–Carex lasiocarpa–Sphagnum magellanicum характерна для широких гряд аапа болот (табл. 3, 3). Высота их 20–35 см, а ширина – 5 м. Уровень болотных вод стоит на 15–20 см ниже сфагнового покрова. Carex lasiocarpa и Betula nana образуют высокий (до 70 см) и густой (покрытие до 40–70%) ярус. Покрытие и обилие Menyanthes сильно варьирует (от 5 до 60%).
Все 3 ассоциации хорошо расчленяются по видовому составу и месту в сукцессионном ряду. Chamaedaphne calyculata–Menyanthes trifoliata–Sphagnum magellanicum является самой продвинутой в сукцессионном ряду и характеризуется наибольшей олиготрофизацией. Только в ней встречаются Ledum palustre и Sphagnum angustifolium. Betula nana–Carex lasiocarpa–Sphagnum magellanicum представляет переход от Sphagneta papillosi к Sphagneta magellanici. Andromeda polifolia–Menyanthes trifoliata–Sphagnum magellanicum – начальный этап в образовании сообществ Sphagneta magellanici путем зарастания небольших кочек или, как предыдущая, переход от Sphagneta papillosi к Sphagneta magellanici.
Сообщества Sphagneta jensenii (табл. 4), как и Sphagneta magellanici, подчеркивают своеобразие северовосточноевропейских аапа болот. Они широко распространены на аапа болотах этого типа и в отличие от всех перечисленных выше сфагновых, характерны для отрицательных форм микрорельефа – мочажин и топей. Участие сообществ со Sphagnum jensenii, как характерную особенность северовосточноевропейских аапа болот, отмечают все их исследователи (Katz, 1948, 1971; Alekseeva, 1974; Boch, Vasilevich, 1980; Goncharova, 2014).
Сообщества Sphagneta jensenii характерны для периферийной части грядово-мочажинного комплекса, а также образуют топяные ковры между грядово-мочажинным комплексом и краевым заболоченным лесом, реже встречаются в виде бордюра вдоль озерков или образуют заливы в их углах и тогда могут быть обнаружены и в центре болота.
Асс. Eriophorum russeolum–Sphagnum jensenii (табл. 4, 1) встречается во всех перчисленных местообитаниях, но наиболее характерна для грядово-топяных комплексов, в которых мочажины-топи достигают ширины 50 м и занимают 70–85% площади комплекса. УБВ колеблется от 0 до ‒5 см. В травяном покрове постоянны Eriophorum russeolum, Menyanthes trifoliata и почти постоянна Carex limosa и нередко обильнее, чем Eriophorum. Эти 2 вида образуют верхний ярус высотой 20–30 см, покрытие его колеблется от 15 до 40%. Почти всегда имеется хорошо выраженный ярус Menyanthes высотой 10–15 см. Моховой покров часто монодоминантный, местами с примесью Sphagnum fallax или Sphagnum flexuosum.
Асс. Scheuchzeria palustris–Sphagnum jensenii (табл. 4, 2) наиболее характерна для мочажин периферийной части грядово-мочажинного комплекса или образует бордюр по краю мочажин-озерков. УБВ колеблется от +1 до –3 см. Помимо шейхцерии в ней, как и в предыдущей, постоянны только Carex limosa и Menyanthes trifoliata. Доминирует всюду Scheuchzeria. Она и Carex limosa формируют первый ярус высотой 15–30 см. Во втором ярусе более или менее обильна Menyanthes. Общее покрытие в фитоценозах этой ассоциации обычно ниже, чем в предыдущей и варьирует от 15 до 30%. Моховой покров рыхлый и эдификаторная роль Sphagnum jensenii во многих сообществах менее выражена. Значительна примесь то мезоолиготрофных видов: Sphagnum papillosum, Sphagnum flexuosum, Loeskipnum badium, то более евтрофных: Sphagnum platyphyllum, Warnstorfia exannulata.
Асс. Carex limosa–Sphagnum jensenii (табл. 4, 3) встречается во всех перечисленных местообитаниях. Carex limosa одна, лишь с незначительной примесью Scheuchzeria в некоторых ценозах, образует ярус; постоянство вахты снижается, но в некоторых сообществах она очень обильна, покрывая до 40% поверхности, в таких фитоценозах относительно велико и общее покрытие, достигая 45%, обычно же оно не превышает 15%. Как и в предыдущей ассоциации, не столь выражена доминирующая роль Sphagnum jensenii, значительна примесь разных видов: от Sphagnum fallax до Warnstorfia exannulata.
Асс. Carex rostrata–Sphagnum jensenii (табл. 4, 4) особенно характерна для топяных ковров с наиболее низким уровнем воды (–5 до –10 см, иногда до –15 см). Это сразу отразилось на постоянстве Oxycoccus palustris, здесь оно 100%, тогда как в других она редка. Menyanthes почти всегда обильна и имеет 100%-е постоянство, покрытие иногда достигает 40–50%. Иногда она и необыкновенно высокая, до 25 см, но обычно 10–15 см. Высокий ярус Carex rostrata (65–75 см) часто весьма густой, также до 40–50% покрытия, но преимущественно 15–20%. Местами хорошо выражен ярус высотой 25–30 см, образованный Carex limosa и Scheuchzeria. Однако в этой ассоциации постоянство Carex limosa снизилось до 70% (в предыдущих оно было 100%). Доминирующее положение Sphagnum jensenii не всегда явное, местами значительно участие близкого по экологии и характерного для топяных ковров Sphagnum fallax.
Только последняя ассоциация имеет группу дифференцирующих видов, остальные различаются только доминантами. Эту особенность болотных синтаксонов с маловидовым составом отмечали неоднократно (Tüxen, 1978; Boch, Vasile-vitch, 1980; Rybniček, 1981 и др.).
Травяные сообщества аапа болот этого типа характеризуются крайней бедностью видового состава и однообразием. Широко распространены почти чистые озерки, в которых встречаются единичные экземпляры Menyanthes trifoliata, Eriophorum angustifolium, Carex limosa, C. lasiocarpa, C. rostrata.
Сообщества ассоциации Carex limosa–Me-nyanthes trifoliata являются самыми распространенными. Они встречаются в обширных сильно обводненных озерках (мочажины-озерки или римпи), в заливах вторичных озерков или образуют бордюр по краю озерков. Размеры мочажин очень варьируют: от 4–5 до 15–30 м в ширину и от 10–15 до 100–150 м в длину. Они занимают значительные площади (до 70%) в грядово-мочажинно-озерковых комплексах и сосредоточены в центральных участках болотных массивов. От близкой ассоциации Carex limosa–Utricularia intermedia карельских аапа она отличается отсутствием Carex livida. Carex livida вообще отсутствует в этом регионе, тогда как для аапа болот Кольско-Карельского региона она чрезвычайно характерна.
Сообщества четко разделяются на 2 яруса: первый с Carex limosa высотой 25–35 см, а второй с Menyanthes – 10–15 см. Обилие и покрытие их сильно колеблются в зависимости от УБВ. Обилие изменяется от 3 до 5 баллов, а покрытие от 15 до 35% каждого из двух видов. Вместе с Carex limosa растут C. chordorrhiza, Scheuchzeria palustris, Pedicularis palustris. Высокие травы (Carex lasiocarpa, C. rostrata, Equisetum fluviatile, Eriophorum angustifolium) образуют разреженный полог, возвышаясь до 60–70 см. Общее покрытие травяного покрова варьирует от 10–15 до 45–50%. УБВ в мочажинах-озерках колеблется от 0 – +5 до +25 – +35, чаще всего +10 – +15 см. Обилие, покрытие и густота стояния того или иного вида изменяется и по годам в зависимости от увлажнения. Так, в 1985 засушливом году на повторно исследуемых болотах в Архангельской области значительно уменьшилось обилие Utricularia intermedia и увеличилась густота стояния Carex limosa. Мхи в этих сообществах редки, более часто отмечены Sphagnum jensenii, Warnstorfia exannulata и W. fluitans.
Травяно-гипновые фитоценозы на этих аапа болотах редки. Это отличает их от карельских аапа болот и свидетельствует об их общей бедности и слабой минерализации вод. Warnstorfia exannulata самый характерный из гипновых мхов вид для северовосточноевропейских аапа болот, но и он редко доминирует.
Асс. Carex limosa–Warnstorfia exannulata – единственная, которую я смогла выделить (табл. 5). Ее фитоценозы встречаются как бордюр и заливы в травяных мочажинах-озерках или в проточных топях. УГВ колеблется от –3 до +3 см. В моховом покрове часто встречается примесь Sphagnum jensenii и S. subsecundum, но Warnstorfia exannulata всегда господствует, составляя 60–80% покрытия. Покров его рыхлый, например в одном фитоценозе обильно росла Utricularia intermedia. Хорошо выражены 3 яруса: первый образован Carex limosa с участием Scheuchzeria palustris, иногда C. chordorrhiza высотою 20–30 см; второй ярус образует Menyanthes, третий ярус – мхи.
Таблица 5.
Значения классов константности видов ассоциации Carex limosa–Warnstorfia exanulata северовосточноевропейских аапа болот Table 5. Values of species constancy classes in Carex limosa–Warnstorfia exanulata association of northeastern European aapa mires
| Вид Species | Константность Constancy |
|---|---|
| Carex limosa | V |
| Menyanthes trifoliata | V |
| Scheuchzeria palustris | II |
| Utricularia intermеdia | III |
| Eriophorum angustifolium | III |
| Carex chordorrhiza | I |
| Pedicularis palustris | II |
| Juncus stygius | I |
| Warnstorfia exanulata | V |
| Sphagnum subsecundum | IV |
| S. jensenii | III |
| S. obtusum | II |
| S. platyphyllum | II |
| Warnstorfia fluitans | II |
| Straminergon stramineum | III |
| Среднее число видов Average number of species | 15 |
| Число описаний Number of releves | 15 |
На северовосточноевропейских аапа болотах хорошо выражен экологический ряд центр–окраина, как и на всех аапа болотах. Вдоль суходолов тянется заболоченный лес, его сменяет кустарничково-сфагновое сосновое редколесье, за которым следует полоса травяно-сфагновых ковров с Eriophorum angustifolium, E. gracile, E. vaginatum, Carex lasiocarpa, C. rostrata, C. pauciflora, Trichophorum cespitosum. Всю центральную часть занимают грядово-мочажинные, грядово-озерковые и озерково-топяные комплексы, нередко практически непроходимые.
Флористический состав северовосточноевропейских аапа болот изучен недостаточно. К настоящему времени известно около 50 видов сосудистых растений и 36 видов листостебельных мхов, из них 21 сфагновые. К числу преобладающих относятся Betula nana, Chamaedaphne calyculata, Carex lasiocarpa, C. limosa, Trichophorum cespitosum, Equisetum heleocharis, Eriophorum angustifolium, Menyanthes trifoliata, Utricularia intermedia, Sphagnum papillosum, S. magellanicum, S. jensenii, Warnstorfia exannulata, W. fluitans, Straminergon stramineum. В восточной части их ареала частой становится Eriophorum russeolum, а на болотах верхнего и среднего течения Печоры появляется Pinus sibirica (Alekseeva, 1988; Boch, Olenskaya, 1980; Goncharova, 2014).
Помимо этих 2-х самых распространенных типов болотных массивов в этом регионе спорадически встречаются сфагновые переходные, травяные низинные, своеобразные ключевые висячие болота, особые оригинальные небольшие болота в карстовых воронках. Все они еще мало изучены и ждут своих исследователей. На самом северо-востоке, на границе с лесотундрой появляются бугристые болота.
Список литературы
[Abolin] Аболин Р.И. 1915. Болотные формы сосны Pinus silvestris L. – Тр. Ботан. музея Акад. наук. 15: 62–84.
[Abramova, Kiryushkin] Абрамова Т.Г., Кирюшкин В.Н. 1968. Районирование болот Архангельской области. – В кн.: Северо-Запад европейской части СССР. Л. Т. 6. С. 81–113.
[Alekseeva] Алексеева Р.Н. 1974. Аапа болота среднего течения р. Печоры. – В кн. Типы болот СССР и принципы их классификации. Л. С. 62–68.
[Alekseeva] Алексеева Р.Н. 1988. Болота Припечорья. Л. 135 с.
[Boch, Olenskaya] Боч М.С., Оленская Н.М. 1980. Болота Якшинского участка. – В кн.: Взаимосвязь компонентов лесных и болотных экосистем северного Приуралья. Л. С. 65–87.
[Boch, Vasilevich] Боч М.С., Василевич В.И. 1980. Болота окрестностей Сегежского озера (южная Карелия). – Бот. журн. 65 (1): 27–38.
[Bogdanovskaya-Guienéuf] Богдановская-Гиенэф И.Д. 1949. Типы верховых болот СССР. – Тр. 2-го Всесоюз. геогр. съезда. М. 3: 144–152.
Czerepanov S.K. 2007. Vascular plants of Russia and adjacent states (the former USSR). Cambridge. 517 p.
Davis R.B., Anderson D.S. 2001. Classification and distribution of freshwater peatlands in Maine. – Northeastern Naturalist. 8(1): 1–50.
[Degteva et al.] Дегтева С.В., Железнова Г.В., Пыстина Т.Н., Шубина Т.П. 2001. Ценотическая и флористическая структура лиственничных лесов Европейского Севера. СПб. 269 с.
[Elina] Елина Г.А. 1969. К истории развития болот юго-восточной части побережья Белого моря. – Бот. журн. 54 (4): 545–552.
[Elina] Елина Г.А. 1971. Типы болот Прибеломорской низменности. – В кн.: Болота Карелии и пути их освоения. Петрозаводск. С. 51–79.
[Elina, Yurkovskaya] Елина Г.А., Юрковская Т.К. 1965. О прибеломорских болотах Карелии. – Бот. журн. 50 (4): 545–552.
[Elina, Yurkovskaya] Елина Г.А., Юрковская Т.К. 1980. Верховые болота на левобережье Северной Двины. – Бот. журн. 65 (7): 958−970.
[Flora…] Флора Северо-Востока европейской части СССР. 1974. Л. Т. 1. 274 с.
[Goncharova] Гончарова Н.Н. 2013. Флора болот бассейна реки Печора. – Вестник института биологии Коми научного центра Уральского отделения РАН. 1 (177): 18–20.
[Goncharova] Гончарова Н.Н. 2014. Болотный заказник “Чукчинское”. – В кн.: Кадастр особо охраняемых природных территорий республики Коми. Сыктывкар. С. 41–42.
[Goncharova, Dubrovskiy] Гончарова Н.Н., Дубровский Ю.А. 2014. Болотный заказник “Пурганюр”. – В кн.: Кадастр особо охраняемых природных территорий республики Коми. Сыктывкар. С. 108.
Hofstetter R.H. 1983. Wetland in the United States. – In: Mires, swamp, bog, fen and moor. Amsterdam. P. 201–244. (Ecosystem of the world; Vol. 4B).
Ignatov M.S., Afonina O.M., Ignatova E.A. et al. 2006. Check-list of mosses of East Europe and North Asia. – Arctoa. 15: 1–130.
[Karta…] Карта растительности европейской части СССР. М 1 : 2 500 000. 1979. М. 6 листов.
[Katz] Кац Н.Я. 1928. О типах олиготрофных сфагновых болот Европейской России и их широтной и меридиональной зональности. – Тр. Ботан. н.-и. ин-та при физ.-мат. фак. Моск. ун-та. С. 1–60.
[Katz] Кац Н.Я. 1937. Типы болот и их размещение на территории европейской части СССР. – Землеведение. 39 (4–5): 388–456.
[Katz] Кац Н.Я. 1948 Типы болот СССР и Западной Европы и их географическое распространение. М. 320 с.
[Katz] Кац Н.Я. 1971. Болота земного шара. М. 295 с.
[Katz, Minkina] Кац Н.Я., Минкина Н.И. 1936. Торфяные болота северной части бассейна реки Печора. – Тр. ЦТОС. 1: 103–125.
[Kiryushkin] Кирюшкин В.Н. 1967. Болотная система “Вадье” и пути ее развития. – В кн.: Природа болот и методы их исследований. Л. С. 47–55.
[Korchagin] Корчагин А.А. 1940. Растительность северной половины Печоро-Илычского заповедника. – Тр. Печоро-Илычского заповедника. 2: 5–412.
Kozhin M.N., Sennikov A.N. 2016. The Russian larch (Larix archangelica, Pinaceae) in the Kola Peninsula. – Memoranda – Societatis pro Fauna et Flora Fennica. 92: 79–91.
[Kucherov] Кучеров И.Б. 2019. Ценотическое и экологическое разнообразие светлохвойных лесов северной и средней тайги Европейской России. СПб. 568 с.
[Kucherov, Zverev] Кучеров И.Б., Зверев А.А. 2011. Лиственничные леса северо-востока Европейской России. – Вестн. Томского гос. ун-та. Биология. 13 (1): 28–50.
[Kutenkov] Кутенков С.А. 2006. Болотные экосистемы. – В кн.: Природа и историко-культурное наследие Кожозерья. Архангельск. С. 39–50.
[Kuznetsov] Кузнецов О.Л. 1980. О развитии аапа болот северной Карелии. – В кн.: Болота Европейского Севера СССР. Петрозаводск. С. 92–113.
[Kuznetsov] Кузнецов О.Л. 1982. Структура и динамика аапа болот северной Карелии. – Бот. журн. 67 (10): 1304–1400.
[Kuznetsov] Кузнецов О.Л. 2017. Разнообразие типов болот таежной зоны европейского севера России. – В кн.: Западно-Сибирские торфяники и цикл углерода: прошлое и настоящее. Матер. Пятого Международного полевого симпозиума. Новосибирск. С. 26–28.
[Kuznetsov et al.] Кузнецов О.Л., Кутенков С.А., Чуракова Е.Ю. 2013. Аапа болота Архангельской области, их разнообразие и генезис. – В кн.: Водно-болотные угодья и пути миграции птиц в Баренцовом / Евроарктическом регионе и вдоль Зеленого пояса Фенноскандии. Матер. Международной научно-практической конференции. С. 122–128.
[Kuznetsov, Yurkovskaya] Кузнецов О.Л., Юрковская Т.К. 2016. Аапа болота на севере Голарктики. – Карельский экологический журнал. С. 18–22.
[Nepomilueva] Непомилуева Н.И. 1974. Кедр сибирский (Pinus sibirica Du Tour) на северо-востоке европейской части СССР. Л. 184 с.
[Nikonov] Никонов М.Н. 1953. Торфяники средней Печоры. – Тр. ин-та леса АН СССР. 13: 148–157.
[Nikonov] Никонов М.Н. 1958а. Торфяники Архангельской области. – В кн.: Торфяной фонд Архангельской области. М. С. III–XVI.
[Nikonov] Никонов М.Н. 1958b. Торфяники Коми АССР. – В кн.: Торфяной фонд Коми АССР. М. С. III–XV.
[Preys] Прейс А.А. 1978. Бугристые болота бассейна реки Хантайки и их динамика. – В кн.: Генезис и динамика болот. Т. 2. М. С. 43–50.
[Rabotnov] Работнов Т.А. 1929. Болота б. Никольского уезда Северо-Двинской губ. – Изв. Лугового ин-та. 4–6: 34–67.
[Rabotnov] Работнов Т.А. 1930. О болотах Архангельского округа. – Торфяное дело. 5: 232–235.
[Ramenskaya, Shubin] Раменская М.Л., Шубин В.И. 1975. Природное районирование в связи с вопросами лесовосстановления. – Лесовосстановление в СССР и Мурманской области. Петрозаводск. С. 180–189.
[Romanova, Usova] Романова Е.А., Усова Л.И. 1969. Геоботаническая и краткая гидрологическая характеристика болотных ландшафтов водораздела рек Вах и Вахтинский Еган Западной Сибири. – Тр. ГГИ. 157: 98–122.
Ruuhijarvi R. 1960. Über die regionale Einteilung der nordfinnischen Moore. – Ann. Bot. Soc. Vanamo. 31 (1): 1–360.
Rybniček K. 1981 Problematika klasifikace rašelinnych spolecenstev. – Zpr. Čs. Bot. Společ. 16 (2): 65–70.
[Shennikov] Шенников А.П. 1941. К ботанической географии лесного северо-востока европейской части СССР. – Тр. БИН АН СССР. Сер. III. Геоботаника. Вып. 4. С. 35–46.
[Smagin et al.] Смагин В.А., Носкова М.Г., Денисенков В.П. 2013. Нахождение аапа-болот в Архангельской области. – Бот. журн. 98 (9): 1095–1108.
[Solonevich] Солоневич Н.Г. 1962. Корневая система болотной сосны. – Проблемы ботаники. Т. 6. М.-Л. С. 297–307.
[Storozheva] Сторожева М.M. 1960. Материалы к характеристике болот восточного склона Северного Урала и Зауралья. – Тр. ин-та биологии УФ АН СССР. 20: 1–54.
[Sukachev] Сукачев В.Н. 1905. О болотной сосне. – Лесной журнал. 35 (3): 354–372.
Tohvanainen T. 2011. Abrupt ombrotrophication of a boreal aapa mire triggered by hydrological disturbance in the catchment. – J. Ecol. 99: 404–415.
Tüxen R. 1978. Remarques sur la synsystématique de la classe Oxycocco-Sphagnetea (I). – Colloque phytosociologuique: Sols tourbeux. Lille. T. 7. P. 383–391.
[Volkova] Волкова E.М. 2000. Морфолого-биологические особенности и изменчивость сосны обыкновенной, произрастающей в условиях олиготрофных болот средней и южной тайги: Автореф. дис. … канд. биол. наук. М. 19 с.
Wells E.D., Zoltai S. 1985. Canadian system of wetlands classification and its application to circumboreal wetlands. – Aquilo. Ser. Bot. 21: 45–52.
[Yudin] Юдин Ю.П. 1954. Темнохвойные леса. – Производительные силы Коми АССР. Т. 3. Ч. 1. М.-Л. С. 42–125.
[Yurkovskaya] Юрковская Т.К. 1977. Болота бассейна реки Сулы (геоботанический очерк). – В кн.: Проблемы экологии, геоботаники, ботанической географии и флористики. Л. С. 59–68.
[Yurkovskaya] Юрковская Т.К. 1984. Виды торфа бореальных болот Архангельской области. – В кн.: Матер. VII Всесоюзн. совещ. Калинин. С. 48–55.
[Yurkovskaya] Юрковская Т.К. 1992. География и картография растительности болот Европейской России и сопредельных территорий. СПб. 256 с.
[Yurkovskaya] Юрковская Т.К. 2003. Верховые болота восточного Прибеломорья как часть единого природно-исторического Поморского региона. – В кн.: Природное и историко-культурное наследие северной Фенноскандии. Петрозаводск. С. 51–57.
Yurkovskaya T.K. 2004. Raised bogs on the North-East of Europe. – Annali di Botanica: nuova serie. IV: 19–28.
[Yurkovskaya] Юрковская Т.К. 2010. Меридиональная зональность и широтная дифференциация растительности болот России. – Направления исследований в современном болотоведении России. СПб. С. 165–178.
[Yurkovskaya] Юрковская Т.К. 2011. Карта растительности. М 1 : 15 000 000 – Национальный атлас почв Российской Федерации. С. 46–51.
[Yurkovskaya, Elina], Юрковская Т.К., Елина Г.А. 1992. Крупномасштабное картографирование палеорастительности голоцена. – Геоботаническое картографирование 1991. СПб. С. 3−12.
[Yurkovskaya] Юрковская Т.К., Паянская-Гвоздева И.И. 1993. Широтная дифференциация растительности вдоль российско-финской границы. – Бот. журн. 78(12): 72−98.
[Zinserling] Цинзерлинг Ю.Д. 1929. Очерк растительности болот по среднему течению р. Печоры. – Изв. Главн. ботан. сада. 29 (1–2): 95–128.
[Zinserling] Цинзерлинг Ю.Д. 1938. Растительность болот. – В кн.: Растительность СССР. Л. Т. 1. С. 355–428.
Zoltai S.C., Pollett F.C. 1983. Wetlands in Canada: their classification, distribution and use. – In: Mires, swamp, bog, fen and moor. Amsterdam. P. 28–43. (Ecosystem of the world; Vol. 4B).
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал