

Ботанический журнал, 2021, T. 106, № 4, стр. 324-335
МОНИТОРИНГ ИЗМЕНЕНИЯ РАСТИТЕЛЬНОГО ПОКРОВА НА ОСУШЕННОМ МЕЗОТРОФНОМ ТРАВЯНО-СФАГНОВОМ БОЛОТЕ В ЮЖНОЙ КАРЕЛИИ
С. И. Грабовик 1, *, Л. В. Канцерова 1, **
1 Институт биологии Карельского научного центра РАН
185910 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: svetagrab@yandex.ru
** E-mail: Kancerova.L@mail.ru
Поступила в редакцию 26.06.2020
После доработки 13.12.2020
Принята к публикации 22.12.2020
Аннотация
На территории Южной Карелии в подзоне средней тайги на лесоболотном научном стационаре Карельского НЦ РАН “Киндасово” в заказнике Койву-Ламбасуо ведется многолетний мониторинг растительности болот. В данной работе представлены результаты 36-летних исследований постмелиоративной динамики видового состава и структуры растительного покрова мезотрофного травяно-сфагнового болота “Близкое” (61°45'10.28"с.ш., 33°28'12.73"в.д). Под влиянием осушения и сукцессий растительного покрова отмечается изменение видового состава растительного покрова, снижение его биоразнообразия в 30 метрах от мелиоративного канала и увеличение видового разнообразия на середине межканальной полосы. На середине межканальной полосы изменения в видовом составе происходят значительно медленнее, чем в 30 метрах от канала, так как удлинился период доминирования гигромезофильных и гигрофильных видов, но при дальнейшем действии осушения происходит уменьшение степени покрытия мезотрофного болотного разнотравья. С помощью бестрендового анализа выделено четыре периода увлажнения, каждый из которых обладает характерным для этого периода растительным покровом. Через 36 лет после осушения на месте мезотрофного травяно-сфагнового кочковато-топяного комплекса сформировался березово-травяной фитоценоз.
Болота имеют огромное значение, являясь резервуарами углерода, органических веществ, и пресной воды (P’yavchenko, 1985).
В республике Карелия болота занимают 3.63 млн га (21%), а заболоченные леса – 1.83 млн га (10.6%) (P’yavchenko, Kolomytsev, 1980). В связи с высокой заболоченностью, в Карелии в середине XX столетия были развернуты гидромелиоративные работы, которые существенно изменяли естественные ландшафты. Возросшее антропогенное влияние на болота приводит к нежелательным последствиям в состоянии естественных экосистем, которых можно избежать только при знании объективных законов внутренней организации и взаимосвязей этих систем с факторами среды. Результаты гидромелиорации могут быть положительными лишь при научно обоснованном прогнозе трансформации заболоченных лесов и болот, а это требует детального изучения растительности и продуктивности фитоценозов в процессе естественных и антропогенных смен растительного покрова разных типов болот.
Вопросы антропогенной динамики растительности болот довольно широко освещаются как в нашей стране, так и за рубежом. Подробный литературный обзор был сделан нами в работах Kuznetsov, Grabovik, 2010; Grabovik et al., 2019.
По мнению Б.С. Маслова “основой мелиоративных исследований должны вновь стать мелиоративно-болотные стационары в разных природно-хозяйственных регионах в сочетании с математическим и физическим моделированием” (Maslov, 2008: 68).
Целью данной работы явилось представление результатов 36-летних исследований постмелиоративной динамики видового состава и структуры растительного покрова в пространственно-временном аспекте на примере мезотрофного травяно-сфагнового болотного массива “Близкое”.
МАТЕРИАЛЫ И МЕТОДЫ
На территории Киндасовского лесо-болотного научного стационара Карельского научного центра РАН в 1970 г. были начаты комплексные научно-исследовательские работы по изучению структуры и динамики болот и заболоченных лесов в естественном состоянии и под влиянием мелиорации. В задачи исследования не входило измерение уровня грунтовых вод (УГВ) в течение вегетационного периода. УГВ на исследованных болотах был измерен только до осушения.
В климатическом отношении территория стационара характеризуется следующими средними многолетними данными: продолжительность вегетационного периода – 148 дней, температура воздуха за вегетационный период – 11.7°С, количество атмосферных осадков за год 565 мм, за вегетационный период – 316 мм (Chesnokov, 1979).
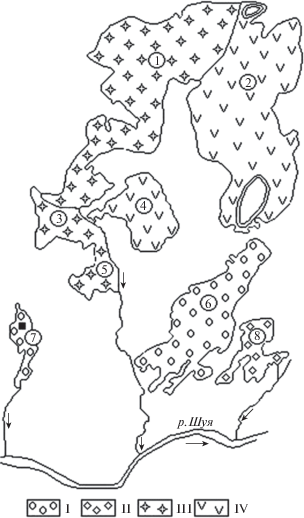
На территории стационара наиболее распространены четыре типа болотных массивов: сфагновый грядово-мочажинный олиготрофный, травяно-сфагновый мезотрофный, травяно-сфагново-гипновый (карельские кольцевые аапа болота) и мезотрофный сфагновый лесной (Elina et al., 1984). Значительная их часть до 1969 г. была в естественном состоянии, затем с 1969 по 1974 г. часть болот была осушена (рис. 1).
Рис. 1.
Схема болотных массивов стационара “Киндасово” (Elina, 1977).
Типы болотных массивов: I – сфагновый грядово-мочажинный олиготрофный; II – мезотрофный травяно-сфагновый; III – мезотрофный сфагновый лесной; IV – южнокарельский вариант кольцевого аапа типа. 7 – болотный массив “Близкое”, ◼ – пробная площадь.
Fig. 1. Scheme of mire ecosystems of the “Kindasovo” Experimental Research Station (Elina, 1977).
Types of mires: I – ombrotrophic ridge-hollow bog; II – mesotrophic herb-Sphagnum; III – mesotrophic Sphagnum-forest mire; IV – aapa mire. 7 – mire ecosystem “Blizkoye”, ◼ – sample plot.
Стационарные исследования проводили в два этапа. На первом этапе основное внимание уделяли изучению растительности естественных (неосушенных) болот. На основании анализа аэрофотоснимков устанавливали соотношение растительных комплексов на болотах (Galkina, 1969). На выделенных по аэрофотоснимкам болотных комплексах выбирали наиболее типичные для них участки (по форме микрорельефа и растительному покрову) – пробные площади (ПП), на которых в последующие годы вели постоянные наблюдения. Всего было заложено 11 ПП (площадь от 0.3 до 0.5 га), здесь выполнялись геоботанические описания, отбирались образцы торфа на ботанический состав и степень разложения, а также измерялся УГВ на коврах, мочажинах, кочках и грядах до осушения.
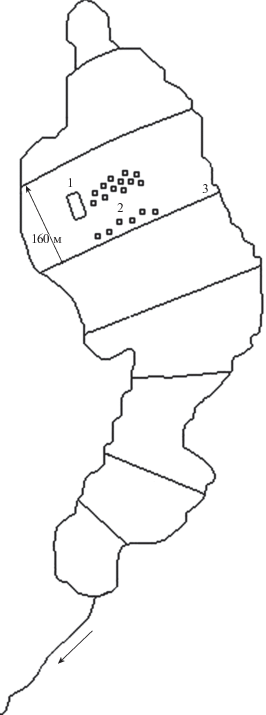
Горизонтальная структура растительного покрова изучалась методом крупномасштабного картирования (Elina et al., 1984) на постоянных пробных площадях (ППП), размеры которых (от 50 до 150 м2) зависели от сложности структуры болотного комплекса и величины отдельных элементов микрорельефа. Площадь исследуемой ПП болотного массива “Близкое” составляет 0.5 га (рис. 1), а площадь ППП – 150 м2 (рис. 2).
Рис. 2.
Схема размещения участков на болоте “Близкое”.
1 – постоянная пробная площадь; 2 – постоянные метровые площадки; 3 – мелиоративные каналы.
Fig. 2. Scheme of plots location in the mire “Blizkoye”.
1 – permanent sample plot; 2 – 1-m2 permanent sample plots; 3 – meliorative channels.
Второй этап начался в первое десятилетие после осушения и характеризовался детализацией ранее выполненных исследований, внимание акцентировалось на динамике растительного покрова, структуре и биологической продуктивности комплексов. Исследования динамики видового состава и структуры растительного покрова выполнялись апробированными и модифицированными методами (Grabovik, 1989, Grabovik et al., 2019). Наблюдения за динамикой видового состава растительного покрова вели на постоянных метровых площадках (1 м2) (ПМП) (рис. 2), расположенных в пределах ПП изучаемого болотного комплекса.
Названия сосудистых растений и мхов приводятся по сводкам (Czerepanov, 1995; Ignatov et al., 2006, 2017; Kravchenko, 2007; Ignatov, Milyutina, 2007). Основные таксационные показатели древостоя определялись методами, применяемыми в лесной таксации (Anuchin, 1982).
В работе используются эколого-ценотические группы (ЭЦГ) видов (Kuznetsov, 2005), выделенные эмпирически для болот Карелии по их отношению к условиям увлажнения, проточности, освещенности и трофности местообитаний, а также сопряженности в сообществах (Kuznetsov, 2002).
Для установления степени сходства видового состава растительных сообществ 1971 года (контроль) с растительными сообществами других лет (с 1972 по 2007 гг.) использовали коэффициент Жаккара (К) (Vasilevich, 1969), который вычисляли по формуле:
где а – число видов в сообществах 1971 года, b – число видов в сообществах других лет, с – число общих видов в сообществах a и b.Для установления экологических особенностей выделенных сообществ и определения их положения в экологическом пространстве был использован бестрендовый анализ соответствия (Detrended correspondence analysis, или DCA ординация), реализованный Hill, Gauch (1980) в прикладном пакете для обработки геоботанических данных DECORANA. В качестве анализируемого параметра использовался показатель проективного покрытия видов (в %) для растительных сообществ ПМП. Нагрузки на оси в ординации рассчитаны с использованием коэффициента Съеренсена.
Мезотрофный травяно-сфагновый болотный массив “Близкое”, площадью 10 га, глубиной торфяной залежи около 1 метра. До осушения центральная часть болота была занята мезотрофным травяно-сфагновым кочковато-топяным комплексом, в котором кочки занимали 30%, мочажины – 70%. В понижениях в течение вегетационного периода УГВ колебался от 0 до +5 см, а на повышениях – 15–25 см ниже поверхности сфагнового ковра. Для более детального изучения горизонтальной структуры растительного покрова в центральной части участка было сделано крупномасштабное картирование. Кочки были заняты кустарничково-осоково-сфагновыми сообществами, с единичными низкими (до 2 м) Betula pubescens и Pinus sylvestris. В неглубоких мочажинах до осушения были сообщества ассоциации Carex lasiocarpa – Menyanthes trifoliata и Carex lasiocarpa – Sphagnum subsecundum. Видовое богатство болотного участка до осушения было невелико и включало 29 видов: из них деревья – 2, кустарнички – 5, травы – 16 и мхи – 6.
В 1970 г. болото было осушено редкой сетью открытых каналов через 160 метров. В настоящее время каналы заросли травянистой растительностью. Мониторинг изменений растительного покрова велся на постоянных метровых площадках (ПМП) 1 × 1 м, заложенных на коврах и мочажинах на разном удалении от мелиоративных каналов (в более интенсивно осушенной части болотного участка, в 30-ти метровой полосе от канала, а также на середине межканальной полосы. Всего заложено 18 ПМП (рис. 2). Первые наблюдения сделаны в год проведения гидромелиорации болотного массива (контроль).
РЕЗУЛЬТАТЫ
Динамика видового состава растительного покрова в 30-метровой полосе вдоль мелиоративного канала
Бестрендовый анализ соответствия был проведен для растительных сообществ ПМП в 30-метровой полосе. Ординационная диаграмма (рис. 3) демонстрирует наличие четко выраженного хронологического градиента (с 1971 по 2007 годы исследования), который в растительных сообществах соответствует градиенту увлажнения от более влажных местообитаний к менее влажным. Анализ показал высокую нагрузку на ось 1, которая объясняет 76% изменчивости. Вдоль градиента нами выделено четыре группы (скопления) растительных сообществ, характеризующиеся степенью увлажнения за разные годы исследования. На ось 2 приходится очень низкая нагрузка, всего 1%.
Рис. 3.
Ординационная диаграмма растительных сообществ на постоянных метровых площадках, расположенных в 30 метрах от мелиоративного канала в разные годы исследования.
Римские цифры – периоды увлажнения.
Fig. 3. Ordination chart of plant communities in permanent 1-m2 sample plots located 30 m from the ditch in different years.
Roman numerals – drainage periods.
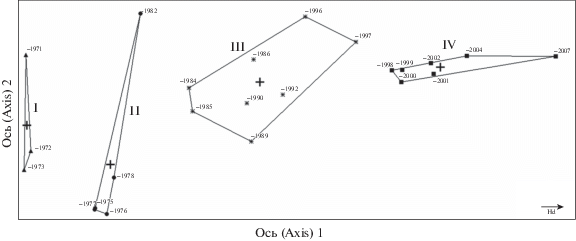
В первую (I) группу (период осушения с 1971 по 1973 год) входят кустарничково-осоково-травяно-моховые сообщества мезоолиготрофных, гидрофильных водно-болотных видов сосудистых растений и мхов. За данный период осушения исчезли виды эколого-ценотических групп, таких как Scheuchzeria palustris и Carex livida (табл. 1), которые характерны для сильно обводненных застойных мочажин: Carex chordorrhiza, С. canescens, C. limosa, C. livida, и мхов Sphagnum angustifolium, S. subsecundum. Доминантными видами сообществ данного периода являются Carex lasiocarpa и Menyanthes trifoliata. Они по-прежнему сохраняют высокую константность, но проективное покрытие C. lasiocarpa снижается с 35 до 15%, а Menyanthes trifoliata увеличивается с 20 до 30%.
Таблица 1.
Постмелиоративная динамика видового состава растительного покрова мезотрофного травяно-сфагнового болота в 30 метрах от мелиоративного канала Table 1. Post-drainage dynamics of the species composition of the vegetation cover of the mesotrophic herb-Sphagnum mire located 30 m from the ditch
| Видовой состав (Plant species) | Периоды осушения (Drainage periods)/Годы (Years) | |||||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | |||||
| 1971 | 1973 | 1975 | 1982 | 1984 | 1997 | 1998 | 2007 | |
| Группа (Group) Chamaedaphne calyculata* | ||||||||
| Chamaedaphne calyculata (L.) Moench | IV3 | IV7 | IV6 | – | – | – | – | – |
| Andromeda polifolia L. | IV2 | III1 | II+ | – | – | – | – | – |
| Oxycoccus palustris Pers. | II+ | III1 | II+ | – | – | – | – | – |
| Sphagnum angustifolium (С.E.O. Jensen ex Russow) С.E.O. Jensen | IV5 | – | – | – | – | – | – | – |
| S. divinum Elatberg et Hassel. | – | I+ | II2 | – | I+ | – | III2 | I1 |
| Группа (Group) Scheuchzeria palustris | ||||||||
| Carex limosa L. | I+ | I+ | – | – | – | – | – | – |
| Группа (Group) Carex lasiocarpa | ||||||||
| Carex lasiocarpa Ehrh. | V33 | V17 | IV6 | IV9 | III7 | II+ | II1 | – |
| Группа (Group) Carex livida | ||||||||
| Carex livida (Wahlenb.) Willd. | II+ | – | – | – | – | – | – | – |
| Sphagnum subsecundum Nees | V18 | V6 | – | – | – | – | – | – |
| Warnstorfia exannulata (Bruch et al.) Loeske | IV5 | – | – | IV38 | – | – | – | – |
| Группа (Group) Menyanthes trifoliata | ||||||||
| Menyanthes trifoliata L. | V22 | V26 | V16 | IV11 | III7 | I1 | – | – |
| Eriophorum angustifolium Honck. | V2 | IV+ | II+ | I+ | – | – | – | – |
| Carex chordorrhiza Ehrh. | V5 | II+ | – | – | – | – | – | – |
| Equisetum fluviatile L. | IV2 | V+ | IV+ | – | – | I2 | I+ | – |
| Группа (Group) Calla palustris | ||||||||
| Salix myrtilloides L. | IV5 | IV5 | IV2 | II1 | – | – | – | – |
| Carex canescens L. | IV7 | I+ | – | – | – | – | – | – |
| Scutellaria galericulata L. | – | – | – | – | – | – | I+ | I+ |
| Pseudobryum cinclidioides (Huebener) T.J. Kop. | – | – | – | – | – | – | I+ | – |
| Climacium dendroides (Hedw.) F. Weber et D. Mohr | – | – | – | – | – | – | I+ | – |
| Группа (Group) Carex acuta | ||||||||
| Calamagrostis neglecta (Ehrt.) Gaertn., B. Mey. et Scherb. | V10 | V5 | V35 | V33 | IV18 | III2 | IV2 | – |
| Comarum palustre L. | IV7 | V4 | V4 | V23 | IV29 | IV10 | V7 | II3 |
| Naumburgia thyrsiflora (L.) Reichenb. | IV+ | V4 | V4 | IV4 | IV5 | III2 | IV4 | IV+ |
| Galium palustre L. | IV3 | V+ | III+ | V1 | III+ | V2 | IV2 | I+ |
| Thyselium palustre (L.) Rafin. | IV+ | IV+ | IV+ | III+ | III1 | I+ | II+ | – |
| Группа (Group) Saxifraga hirculus | ||||||||
| Epilobium palustre L. | – | – | – | – | III1 | – | – | – |
| Группа лесных видов (Group of forest species) | ||||||||
| Rubus idaeus L. | – | – | – | – | – | – | II5 | II20 |
| Angelica sylvestris L. | – | – | – | – | – | I+ | II1 | – |
| Deschampsia cespitosa (L.) P. Beauv. | – | – | – | – | – | I2 | IV14 | V9 |
| Dryopteris carthusiana (Vill.) Y.P. Fuchs | – | – | – | – | – | – | IV2 | IV20 |
| Equisetum sylvaticum L. | – | II+ | II+ | III+ | I+ | III12 | V9 | V5 |
| Geum rivale L. | – | – | – | – | – | I2 | I3 | I+ |
| Trientalis europaea L. | – | – | – | – | – | I1 | I+ | – |
| Brachythecium rivulare Bruch et al. | – | – | – | – | – | – | – | IV4 |
| Polytrichum longisetum Sw. ex Brid. | – | – | – | – | – | – | I+ | – |
| Sciuro-hypnum curtum (lindb.) Ignatov. | – | – | – | – | – | I1 | I2 | I+ |
| Dicranum scoparium Hedw. | – | – | – | – | – | – | – | II+ |
Примечание.*– Эколого-ценотические группы. Единично встречающиеся виды в таблицу не включены. В таблицу не вошли все годы исследования из-за достаточно большого объема материала.
Note.*– Ecological-cenotic groups. Species occurring as single individuals are not included in the Table. Not all years of the study were included in the Table due to a considerable amount of the material.
Во вторую (II) группу (период осушения с 1975 по 1982 год) входят кустарничково-разнотравные сообщества мезотрофных, гигрофильных болотных видов. Различные виды болотных растений реагируют на осушение неодинаково. Одни быстро отмирают (виды ЭЦГ Chamaedaphne calyculata), а другие испытывают явное угнетение и отмирают постепенно (виды ЭЦГ Menyanthes trifoliata). Встречаются виды, которые развиваются даже лучше и роль их в покрове заметно увеличивается после осушения, особенно Calamagrostis neglecta и Comarum palustre (табл. 1). Их проективное покрытие в растительных сообществах, в отличие от первых лет осушения, резко возрастает в среднем с 5–10% до 25–40%. Данные виды относятся к ЭЦГ Carex acuta, произрастающих в топяных местообитаниях, ежегодно заливаемых на довольно длительное время. Количество осадков за данный период было выше средней нормы (в среднем 450 мм).
Сравнение коэффициента сходства первого года осушения с последующими годами I и II периодов показало снижение сходства видового состава растительных сообществ с 74% в 1972 году до 45% в 1982 году. Это связано с тем, что в первые годы осушения резко проявляется дренажный эффект на ПМП, находящихся ближе к мелиоративному каналу и происходит смена более влаголюбивых видов на менее влаголюбивые. Ослабление фитоценотической роли болотных растений создало наиболее благоприятные условия для естественного облесения болот уже в первые 7–10 лет после осушения. В это время гигромезофильные и мезофильные виды (конкуренты всходов древесных пород) еще не получили широкого распространения. Происходит массовый занос семян Betula pubescens, которые попадают в благоприятную для прорастания среду. Число особей Betula pubescens высотой от 0.5 до 5.5 м достигает 3400 экз./га.
В третью (III) группу (период осушения с 1984 по 1997 год) входят березово-травяные сообщества мезотрофных, гигромезофильных болотных видов. Растения гигрофильные угнетаются, конкуренция с их стороны практически устраняется. В напочвенном покрове ковров и бывших мочажин за этот период осушения большинство болотных видов продолжает сохраняться, но жизненность их снижается, они не цветут и не плодоносят (ЭЦГ Carex lasiocarpa, Menyanthes trifoliata, Carex acuta). Происходит изреживание травяного покрова, появляются виды (относящиеся к ЭЦГ лесных видов), хотя и в незначительном количестве, отсутствовавшие здесь до осушения. На низких приствольных кочках в сообществах появляются Deschampsia cespitosa, Polytrichum longisetum и Dicranum scoparium.
Сравнение коэффициента сходства первого года осушения с последующими годами III периода показалo, что коэффициент Жаккара снижается, но незначительно: с 39% в 1984 году до 35% в 1997 и число общих видов колеблется от 7 до 9.
В сформировавшемся березово-травяном фитоценозе общее число стволов составило 1850 экз./га, в том числе Betula pubescens 1700. Средний диаметр 5.6 см, высота – 6.9 м. Значительная часть Betula pubescens появилась здесь уже после осушения.
В четвертую (IV) группу (период осушения с 1998 по 2007 год) входят березово-травяные сообщества, мезотрофных, мезофильных лесо-болотных и преимущественно лесных видов.
В напочвенном покрове ковров болотные растения еще сохранились, но проективное покрытие их значительно снизилось, продолжается внедрение лесных видов (табл. 1) и в сложении напочвенного покрова они уже играют значительную роль. ЭЦГ лесных видов указывают на переменность водного режима и динамические связи болот и лесов. Доминантными видами сообществ данного периода являются Equisetum sylvaticum, Deschampsia cespitosa, Dryoptheris cartusiana. Из сообществ исчезли Carex lasiocarpa и Menyanthes trifoliata, которые в первом периоде осушения были доминантными видами. Также исчезли Equisetum fluviatile и Tyselium palustre. В сообществах появились встречающиеся ранее 1998 г. виды мхов: Climacium dendroides, Polytrichum longisetum, Brachythecium rivulare, Pseudobryum cinclidioides.
В целом произошло значительное снижение сходства видового состава (коэффициент Жаккара снизился с 24% в 1998 г. до 10% в 2007 г.).
Следует отметить, что такие светолюбивые растения, как Calamagrostis neglecta и Deschampsia cespitosa образуют здесь злаковые сообщества, которые на открытых местах (в окнах) препятствуют лесовозобновительному процессу. А такие виды, как Angelica sylvestris, Dryoptheris cartusiana, Equisetum sylvaticum, Trientalis europaea, Rubus idaeus и др., составляют группу теневыносливых видов, наибольшее распространение которых отмечается под пологом Betula pubescens.
За этот период осушения в составе древостоя произошли существенные изменения. Состав древостоя 9Б35 1С55, относительная полнота 0.9. Общее число стволов древостоя сократилось до 1658 экз./га, в том числе Betula pubescens до 1516.
Растительный покров до осушения был представлен 20 видами сосудистых растений и мхов, который через 36 лет после осушения представлен 13 видами, и только три вида являются общими: Comarum palustre, Galium palustre и Naumburgia thyrsiflora. Они обладают широкой экологической пластичностью, что позволяет им существовать в различных эколого-фитоценотических условиях.
Динамика видового состава растительного покрова на середине межканальной полосы
Бестрендовый анализ соответствия также был проведен для растительных сообществ ПМП на середине межканальной полосы (рис. 4). Здесь также значительна нагрузка на ось 1 (79%), интерпретируемая, как хронологический градиент, соответствующий градиенту увлажнения. Нагрузка на ось 2 низкая, 1%. Нами выделены четыре группы растительных сообществ, характеризующиеся степенью увлажнения за разные годы исследования.
Рис. 4.
Ординационная диаграмма растительных сообществ на постоянных метровых площадках, расположенных на середине межканальной полосы в разные годы исследования.
Римские цифры – периоды увлажнения.
Fig. 4. Ordination chart of plant communities in permanent 1-m2 sample plots located in the middle of a drained row in different years.
Roman numerals – drainage periods.
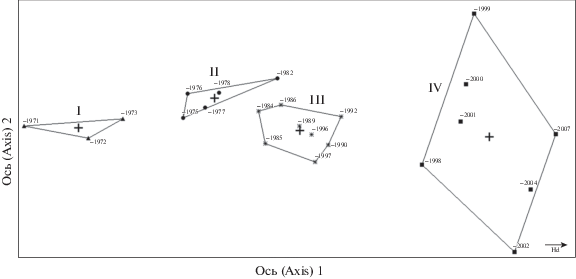
На середине межканальной полосы (табл. 2) изменения в видовом составе происходят в том же направлении, что и в приканальной полосе, но значительно медленнее, удлинился период доминирования гигромезофильных и гигрофильных видов, но при дальнейшем действии осушения происходит уменьшение степени покрытия мезотрофного болотного разнотравья. Внедрение лесных видов здесь отмечается только с 1998 года, но доля их участия в напочвенном покрове совсем незначительна.
Таблица 2.
Постмелиоративная динамика видового состава растительного покрова мезотрофного травяно-сфагнового болота на середине межканальной полосы Table 2. Post-drainage changes of the species composition of the vegetation cover in the mesotrophic herb-Sphagnum mire in the middle of a drained row
| Видовой состав (Plant species) | Периоды осушения (Drainage periods)/Годы (Years) | |||||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | |||||
| 1971 | 1973 | 1975 | 1982 | 1984 | 1997 | 1998 | 2007 | |
| Группа (Group) Chamaedaphne calyculata* | ||||||||
| Chamaedaphne calyculata (L.) Moench | IV3 | III3 | IV3 | I3 | – | – | – | – |
| Andromeda polifolia L. | V8 | IV5 | IV5 | – | – | – | – | – |
| Oxycoccus palustris Pers. | IV4 | IV4 | V3 | – | – | I+ | I+ | – |
| Sphagnum angustifolium (С.E.O. Jensen ex Russow) С.E.O. Jensen | – | – | I1 | II+ | I1 | I+ | – | – |
| S. divinum Elatberg et Hassel. | – | – | – | – | – | – | I+ | I1 |
| Группа (Group) Scheuchzeria palustris | ||||||||
| Carex limosa L. | I+ | I+ | – | – | – | – | – | – |
| Группа (Group) Carex lasiocarpa | ||||||||
| Carex lasiocarpa Ehrh. | IV25 | V14 | V11 | V16 | V31 | III5 | II+ | – |
| Группа (Group) Carex livida | ||||||||
| Carex livida (Wahlenb.) Willd. | I+ | I+ | – | – | – | – | – | – |
| Sphagnum subsecundum Nees | IV29 | III5 | III5 | – | – | – | – | – |
| Warnstorfia exannulata (Bruch et al.) Loeske | – | I1 | I1 | II1 | – | – | – | – |
| Группа (Group) Menyanthes trifoliata | ||||||||
| Menyanthes trifoliata L. | IV6 | V13 | V9 | V9 | V15 | V9 | IV4 | II1 |
| Eriophorum angustifolium Honck. | II+ | II+ | II+ | – | – | – | – | – |
| Carex chordorrhiza Ehrh. | V20 | I1 | I+ | I+ | – | – | – | – |
| Equisetum fluviatile L. | – | V1 | II+ | – | II+ | II1 | – | – |
| Группа (Group) Calla palustris | ||||||||
| Salix myrtilloides L. | IV13 | IV8 | IV9 | IV4 | I+ | – | – | – |
| Carex canescens L. | I+ | II+ | I2 | – | – | – | III2 | – |
| Scutellaria galericulata L. | – | – | – | – | – | – | I1 | I1 |
| Pseudobryum cinclidioides (Huebener) T.J. Kop. | – | – | – | – | – | – | I1 | I1 |
| Climacium dendroides (Hedw.) F. Weber et D. Mohr | – | – | – | – | – | – | I+ | I+ |
| Группа (Group) Carex acuta | ||||||||
| Calamagrostis neglecta (Ehrt.) Gaertn., B. Mey. et Scherb. | – | IV5 | IV11 | V27 | III18 | V7 | V7 | V9 |
| Comarum palustre L. | V11 | V6 | V8 | V21 | V43 | V6 | V6 | IV6 |
| Galium palustre L. | III3 | IV+ | IV+ | V2 | III1 | IV5 | III1 | III+ |
| Naumburgia thyrsiflora (L.) Reichenb. | III+ | V1 | V4 | V3 | III1 | IV3 | V5 | III+ |
| Thyselium palustre (L.) Rafin. | IV+ | IV+ | V3 | V1 | IV1 | II+ | III+ | I+ |
| Группа (Group) Saxifraga hirculus | ||||||||
| Epilobium palustre L. | II+ | – | – | II+ | III1 | I+ | – | I+ |
| Группа лесных видов (Group of forest species) | ||||||||
| Equisetum sylvaticum L. | – | III+ | II+ | III+ | – | III4 | IV1 | I+ |
| Drepanocladus polygamus (Bruch et al.) Hedenäs | – | – | II1 | II1 | – | – | – | – |
| Pleurozium schreberi (Brid.) Mitt. | – | – | – | – | – | I1 | I1 | I1 |
| Geum rivale L. | – | – | – | – | – | III3 | – | I+ |
| Angelica sylvestris L. | – | – | – | – | – | I1 | I1 | I1 |
| Deschampsia cespitosa (L.) P. Beauv. | – | – | – | – | – | I+ | III6 | |
| Trientalis europaea L. | – | – | – | – | – | I2 | III2 | II+ |
| Dryopteris carthusiana (Vill.) Y.P. Fuchs | – | – | – | – | – | – | III1 | III6 |
| Sciuro-hypnum curtum (lindb.) Ignatov. | – | – | – | – | – | – | I+ | I+ |
| Dicranum scoparium Hedw. | – | – | – | – | – | – | I+ | I+ |
| Brachythecium rivulare Bruch et al. | – | – | – | – | – | – | I+ | V5 |
| Polytrichum longisetum Sw. ex Brid. | – | – | – | – | – | – | I+ | I+ |
Примечание. *– Эколого-ценотические группы. Единично встречающиеся виды в таблицу не включены. В таблицу не вошли все годы исследования из-за достаточно большого объема материала.
Note. *– Ecological-cenotic groups. Species occurring as single individuals are not included in the Table. Not all years of the study were included in the Table due to a considerable amount of the material.
При слабой степени осушения Calamagrostis neglecta сохраняет очень высокий коэффициент участия в растительном покрове, он разрастается, продуцирует мощную дернину и слой сухой ветоши 5–7 см, в результате чего облесения не наблюдается, так как происходит вымокание семян древесных пород.
Сравнение коэффициента сходства первого года осушения с последующими годами показало снижение сходства видового состава с 65% в 1972 г. до 19% в 2007 г. Растительный покров до осушения был представлен 17 видами сосудистых растений и мхов, который через 36 лет после осушения сократился до 22 видов, из которых 6 являются общими: Comarum palustre, Galium palustre, Epilobium palustre, Menyanthes trifoliata, Naumburgia thyrsiflora и Thyselium palustre.
ЗАКЛЮЧЕНИЕ
Мониторинг изменения растительного покрова на осушенном мезотрофном травяно-сфагновом болоте “Близкое” Южной Карелии позволил проследить ход сукцессий растительности, выражающийся в обобщенных временных сериях. Эти серии являются моделью, отображающей направление происходящих смен фитоценозов во времени под влиянием осушения.
Под влиянием осушения и сукцессий растительного покрова на месте мезотрофного травяно-сфагнового кочковато-топяного комплекса через 36 лет после осушения сформировался березово-травяной фитоценоз. В древесном ярусе общее число стволов составляет 1658 экз./га, в том числе Betula pubescens 1516 экз./га.
В напочвенном покрове виды эколого-ценотических групп Chamaedaphne calyculata, Scheuchzeria palustris и Carex livida, которые произрастают в сильно обводненных застойных мочажинах, чутко реагируют на изменение гидрологического режима и выпадают из растительного покрова в первый период осушения. При дальнейшем действии осушения (II и III периоды) происходит угнетение видов таких ЭЦГ, как Carex lasiocarpa, Menyanthes trifoliata, Carex acuta, характеризующихся высокой степенью увлажнения. Через 30 лет (IV период) в сложении напочвенного покрова значительную роль играют виды ЭЦГ лесных видов, индицирующие переменность водного режима, а роль большинства видов в растительном покрове незначительна, так как они находятся на границах своих фитоценотических амплитуд.
Сравнение коэффициента сходства первого года осушения с последующими годами показало снижение сходства видового состава, как в 30-метровой полосе от канала (с 24% в 1998 году до 10% в 2007), так и на середине межканальной полосы (с 65% в 1972 году до 19% в 2007 году). Под влиянием осушения в 30-метровой полосе отмечается не только изменение растительного покрова, но и снижение его биоразнообразия (с 20 видов в 1972 году до 13 – в 2007), из которых только три вида являются общими: Comarum palustre, Galium palustre и Naumburgia thyrsiflora. На середине межканальной полосы в течение 36 лет происходит увеличение видового разнообразия с 17 до 22 видов, в сообществах на ПМП еще сохраняются болотные виды растений, а также появляются и лесные, при этом шесть видов являются общими: Comarum palustre, Galium palustre, Epilobium palustre, Menyanthes trifoliata, Naumburgia thyrsiflora и Thyselium palustre.
Главным экологическим фактором, обеспечивающим свыше 2/3 всей изменчивости растительности на постоянных метровых площадках, как на середине межканальной полосы, так и в 30‑метровой полосе является хронологический градиент, который в растительных сообществах соответствует градиенту увлажнения от более влажных местообитаний к менее влажным. На ординационной диаграмме нами выделено четыре периода увлажнения, каждый из которых определяется характерным для этого периода растительным покровом. В первые три года осушения растительный покров был представлен кустарничково-осоково-травяно-моховыми сообществами мезоолиготрофных, гидрофильных водно-болотных видов сосудистых растений и мхов. В период осушения с 1975 по 1982 год входят кустарничково-разнотравные сообщества мезотрофных, гигрофильных болотных видов. С 1984 по 1997 год сформировался березово-травяной фитоценоз, в растительный покров которого входят мезотрофные, гигромезофильные болотные виды, а в последнее десятилетие наших исследований на постоянных метровых площадках доминируют березово-травяные сообщества мезотрофных, мезофильных лесо-болотных и преимущественно лесных видов.
Список литературы
[Anuchin] Анучин Н.П. 1982. Лесная таксация. М. 552 с.
[Czerepanov] Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб. 992 с.
[Chesnokov] Чесноков В.А. 1979. Изменение стока с заболоченных водосборов южной Карелии под влиянием лесоосушения: Автореф. дис. … канд. геогр. наук. Л. 25 с.
[Elina] Елина Г.А. 1977. Типы болот Шуйской равнины. – В кн.: Стационарное изучение болот и заболоченных лесов в связи с мелиорацией. Петрозаводск. С. 5–19.
[Elina et al.] Елина Г.А., Кузнецов О.Л., Максимов А.И. 1984. Структурно-функциональная организация и динамика болотных экосистем Карелии. Л. 128 с.
[Galkina] Галкина Е.А. 1969. Аэрометоды и их значение в развитии ландшафтного болотоведения. – В кн.: Доклады комиссии аэросъемки и фотограмметрии. Л. 6: 26–35.
[Grabovik] Грабовик С.И. 1989. Динамика растительного покрова болотных массивов мезотрофного травяно-сфагнового типа под влиянием осушения. – Бот. журн. 74 (12): 1757–1768.
[Grabovik et al.] Грабовик С.И., Канцерова Л.В., Кутенков С.А. 2019. Влияние осушения на динамику растительного покрова мезотрофных травяно-сфагновых болот южной Карелии. – Бот. журн. 104 (6): 888–899.
Игнатов М.С., Афонина O.M., Игнатова E.A., Aболиня A.A., Акатова T.В., Баишева Э.З., Бардунов Л.В., Барякина E.A., Белкина O.A., Безгодов A.Г., Бойчук М.А., Черданцев В.Я., Чернядьева И.В., Дорошина Г.Я., Дьяченко A.П., Федосов В.Э., Гольдберг И.Л., Иванова E.И., Юкониене И., Каннукене Л., Казановский С.Г., Харзинов З.Х., Курбато-ва Л.Е., Максимов А.И., Маматкулов У.K., Манакян В.A., Масловский O.M., Напреенко M.Г., Отнюкова T.Н., Партыка Л.Я., Писаренко O.Ю., Попова Н.Н., Рыковский Г.Ф., Тубанова Д.Я., Железнова Г.В., Золотов В.И. 2006. Check-list of mosses of East Europe and North Asia. – Arctoa. 15: 1–130.
[Ignatov, Milyutina] Игнатов М.С., Милютина И.А. 2007. Ревизия рода Sciuro-hypnum (Brachytheciaceae, Bryophyta) в России. – Arctoa. 16: 63–86.
[Ignatov et al.] Игнатов М.С., Игнатова Е.А., Федосов В.Э., Иванова Е.И., Блом Х.Х., Муньос И., Беднарек-Охыра Х., Афонина О.М., Курбатова Л.Е., Чернядьева И.В., Черданцева В.Я. 2017 Флора мхов России. Т. 2. Oedipodiales – Grimmiales. М. 560 с.
Hill M.O., Gauch H.G. 1980. Detrended correspondence analysis: an improved ordination technique. – Vegetatatio. 42: 47–58.
[Kravchenko] Кравченко А.В. 2007. Конспект флоры Карелии. Петрозаводск. 403 с.
[Kuznetsov] Кузнецов О.Л. 2002. Использование эколого-ценотических групп видов для классификации болотной растительности. – Вестн. Томского ун-та. 2: 111–115.
[Kuznetsov] Кузнецов О.Л. 2005. Тополого-экологическая классификация растительности болот Карелии (омбротрофные и олиготрофные сообщества). – Тр. КарНЦ РАН. 8: 15–46.
[Kuznetsov, Grabovik] Кузнецов О.Л., Грабовик С.И. 2010. Мониторинг флоры и растительности болотных экосистем. – В кн: Мониторинг и сохранение биоразнообразия таежных экосистем Европейского Севера России. Петрозаводск. С. 19–31.
[Lopatin] Лопатин В.Д. 1983. Экологические ряды растительности болот. – В кн.: Структура растительности и ресурсы болот Карелии. Петрозаводск. С. 5–38.
[Maslov] Маслов Б.С. 2008. Вопросы истории, мелиорации торфяных болот и развитие науки. – Вестн. ТГПУ. 4 (78): 64–69.
[P’yavchenko] Пьявченко Н.И. 1985. Торфяные болота, их природное и хозяйственное значение. М. 152 с.
[P’yavchenko, Kolomytsev] Пьявченко Н.И., Коломыцев В.А. 1980. Влияние осушительной мелиорации на лесные ландшафты Карелии. – В кн.: Болотно-лесные системы Карелии. Л. С. 52–71.
[Vasilevich] Василевич В.И. 1969. Статистические методы в геоботанике. Л. 232 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал