

Ботанический журнал, 2021, T. 106, № 8, стр. 769-787
МИКРОМОРФОЛОГИЯ СЕМЯН У НЕКОТОРЫХ ВИДОВ ATOCION, IXOCA, MINJAEVIA И VISCARIA (SILENEAE, CARYOPHYLLACEAE)
Т. И. Кравцова 1, *, В. О. Романова 1, **
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
* E-mail: kraveleon@yandex.ru
** E-mail: veronique71@mail.ru
Поступила в редакцию 28.01.2021
После доработки 24.04.2021
Принята к публикации 27.04.2021
Аннотация
С использованием сканирующего электронного микроскопа изучены морфология семени и микроморфология семенной кожуры у 7 видов из родов Atocion, Minjaevia и Viscaria, составляющих на молекулярно-филогенетических деревьях трибы Sileneae кладу Viscaria, не соответствующую представлениям традиционной систематики, а также рода Ixoca, ранее входившего в эту кладу. Выяснено, что признаки семени, прежде всего форма и ориентация клеток экзотесты, имеют в исследуемой группе важное таксономическое и диагностическое значение. Полученные результаты подтвердили, что род Ixoca не входит в кладу Viscaria group и занимает обособленное положение в трибе Sileneae. Обращает на себя внимание точное повторение характерного признака семян Ixoca – коронки из нескольких рядов плоских волосков по периферии семени у некоторых видов рода Minuartia, изученных Song et al. и Zaychenko, Zernov. Проведенное исследование подтвердило также близкое родство Atocion и Minjaevia, выявленное молекулярными методами. Вместе с тем, наши результаты расходятся с молекулярными данными о сестринских отношениях этих родов с родом Viscaria. В роде Viscaria выявлены два типа семени (V. alpina и V. viscosa), которые значительно различаются по микроморфологии. Оба типа заметно отличаются от семян Atocion и Minjaevia и обнаруживают сходные черты по этим признакам с родом Lychnis. Разнотипность семян Viscaria согласуется с предложенным С.С. Иконниковым (1987) делением этого рода на секции.
Молекулярно-филогенетические исследования трибы Sileneae DC., проведенные с использованием разных молекулярных маркеров, ядерного и хлоропластного происхождения (Oxelman, Lidén, 1995; Oxelman et al., 1997), показали, что одной из базальных клад на филогенетическом дереве трибы является смолковая клада (Viscaria group), объединяющая четыре морфологически различающихся мелких рода: Atocion Adans., Ixoca Raf. (=Heliosperma (Rchb.) Rchb.), Minjaevia Tzvel. и Viscaria Bernh. Позже было показано, что положение здесь рода Ixoca является ошибочным (Fraiman, Oxelman, 2007; Fraiman et al., 2009a). Этот род, происхождение которого связывают с процессами гибридизации, обособлен в трибе Sileneae и занимает в ней неясное положение. Родственные связи Ixoca на молекулярно-филогенетических деревьях трибы зависят от методов исследования и выбранных генных маркеров; по некоторым данным, он составляет отдельную базальную кладу (Mikhailova, 2016).
Молекулярные методы выявили близкое родство родов Atocion и Minjaevia (Oxelman et al., 2001, Fraiman et al., 2009b; 2013). Вид Minjaevia rupestris (L.) Tzvel.) (= Silene rupestris L.) был включён в род Atocion, как Atocion rupestre (L.) Oxelman. Сестринские отношения между Atocion и Viscaria являются одним из неожиданных результатов, полученных Oxelman и Lidén (1995), подтвержденным всеми последующими молекулярными исследованиями (Fraiman et al., 2009b; Jafari et al., 2020). Ранее род Atocion сближали с Silene L. s. str., a род Viscaria – с родом Lychnis L. (например, Chowdhuri, 1957; Tzvelev, 2001). Противоречия с традиционной систематикой трибы Sileneae связывают с возможным сетчатым характером их эволюции (Erixon, Oxelman, 2008; Fraiman et al., 2009b; Mikhailova, 2016).
Морфология семян у представителей трибы Sileneae издавна, начиная с работ E. Boissier (1867) и P. Rohrbach (1868), используется в целях его систематики. В.И. Кожанчиков (Kozhanchikov, 1969) и V. Melzheimer (1977, 1980, 1988) одними из первых авторов обратили внимание на важное таксономическое значение признаков микроморфологии семенной кожуры в этой группе, в частности, формы клеток экзотесты. За последние десятилетия, после появления техники сканирующей электронной микроскопии, число исследований микроморфологии семян у таксонов трибы Sileneae, как и других триб семейства Caryophyllaceae, заметно возросло (Ghazanfar, 1983; El-Oqlah, Karim, 1990 – цит. по: Fawzi et al., 2010; Nersesian, 1990; Kovtonjuk, 1995; Yildiz, Cirpici, 1998; Hong et al., 1999; Yildiz, 2002; Zareh, 2005; Perveen, 2009; Fawzi et al., 2010; Camelia, 2011; Ocaña et al., 2011; Arman, Gholipour, 2013; Tabaripour et al., 2013; Gholipour, Kuhdar, 2014; Martynyuk et al., 2015; Keshavarzi et al., 2015; Dadandi, Yildiz, 2015; Abdel-Maksoud, Fawzi, 2016; Romanova, Kravtsova, 2016, 2019; Atazadech et al., 2017; Hoseini et al., 2017; Kuh et al., 2017). Использованные для сравнительного анализа признаки семени несколько различаются в этих работах, и нет унифицированной общепринятой их классификации и терминологии. Различные признаки были выбраны для выделения типов семян: наличие или отсутствие бороздки на спинке (Hong et al., 1999), очертания клеток экзотесты, форма и скульптура их наружной периклинальной стенки, характер волнистости антиклинальных стенок (Fawzi et al., 2010), очертания семени, строение и орнаментация латеральных его сторон (Ocaña et al., 2011), строение семени, орнаментация дорсальной и латеральных его сторон (Candáu, Talavera, 1978). В некоторых работах для определения степени родства изученных таксонов проведен статистический анализ признаков семени (Zareh, 2005; Fawsi et al., 2010; Arman, Gholipour, 2013; Tabaripour et al., 2013; Keshavarzi et al., 2015; Abdel-Maksoud, Fawzi, 2016). Семена значительного числа видов трибы Sileneae (56), в том числе входящие в Viscaria group, кратко описаны в атласе семян центрально- и восточноевропейской флоры (Bojňanský, Fargašová, 2007). Перечисленные исследования носят в основном региональный характер, выполнены нередко для стран Юго-Западной Азии и Средиземноморья, где находятся основные центры разнообразия рода Silene s.l. (Greuter, 1995). Полученные в них результаты и обзор этих работ (например, Ocaña et al., 2011, Hoseini et al., 2017) показали, что признаки семени важны для таксономии рода Silene s.l., но имеют ограниченное использование – для таксономии и диагностики лишь некоторых видов. Во многих исследованиях отмечено, что признаки семени, включая орнаментацию семенной кожуры, не могут быть применены для разграничения секций рода, так как семена нередко морфологически очень разнородны в пределах одной секции, и сходные типы семян имеются у видов из разных секций (Candáu, Talavera, 1978; Ghazanfar, 1983; El-Oqlack, Karim, 1990 (цит. по: Fawsi et al., 2010); Hong et al., 1999; Fawsi et al., 2010; Ocaña et al., 2011; Hoseini et al., 2017). Из изученных E. Hoseini с соавторами 22 секций рода Silene (по классификации Melzheimer, 1988) только три имели однотипные семена. M.E. Ocaña et al. (2011) в обсуждении полученных результатов, со ссылкой на работы D. Jeanmonod (1985) и V. Melzheimer (1987), указали, что это явление может быть связано либо с предполагаемой искусственностью границ между секциями, либо с процессами гибридизации или влиянием окружающей среды. Процессами гибридизации объясняют также гетерогенность семян в пределах одного вида или подвида (Aeschimann, 1984).
Хотя морфология семян в трибе Sileneae довольно широко изучалась, в том числе и у интересующих нас видов, сравнительные исследования структур семени в рамках Viscaria group ранее не проводились. Задача работы – проведение сравнительно-морфологического изучения семян у представителей родов, входящих в кладу Viscaria group, степень родства которых установлена методами молекулярной филогении. Ее цель – предварительная оценка таксономического и диагностического значения признаков семени в данной кладе. В сравнительный анализ включены также два вида рода Ixoca, ранее относимого к кладе Viscaria.
МАТЕРИАЛ И МЕТОДЫ
Материалом послужили семена 7 видов (23 образца), полученные из Гербариев LE (БИН РАН), KW (Институт ботаники им. Н.Г. Холодного НАН Украины, Киев), карпологической коллекции Музея БИН РАН (Lec), по обмену семян, а также собранные в природе. Таксономия группы соответствует классификации Н.Н. Цвелева (Tzvelev, 2001); учитывали также более позднюю ревизию родов Atocion и Viscaria (Fraiman et al., 2013).
Изученные виды и образцы
Изучены 3 вида рода Atocion Adans. из 6 (согласно классификации B. Fraiman et al., 2013):
A. armeria (L.) Raf.: Парк БИН, 4 IX 2004, Н.Н. Цвелев 26 (LE); Гомельская губ., ст. Добруш, 25 VII 1926, Ю. Кречетович 24 (LE); Санкт-Петербург, газон на улице С. Ковалевской, сентябрь 2014, Г.Ю. Конечная; Дальний Восток, г. Благовещенск, Амурский филиал БСИ ДВО РАН, сентябрь 2016 г., Я.В. Болотова. Вид произрастает в южной, центральной и восточной Европе на щебнистых сухих лугах, скалах, лесных опушках и полянах (Fraiman et al., 2013); часто культивируется как декоративный.
A. lithuanicum (Zapał.) Tzvelev: Украина, Киевская обл. 25 V 75, М.В. Клонов, № 033131 (KW); Украина, Волынская обл., 13 VIII 2007, Д. Екушенко, № 092475 (KW); Украина, Я.П. Дидух, № 072256 (KW). Вид известен из восточной Европы. Согласно V. Martynyuk et al. (2015), он эндемик Полесья (Украина), Fraiman et al. (2013) указывают более широкое его распространение. Вид обычно считается синонимом или разновидностью A. armeria.
Minjaevia rupestris (L.) Tzvelev (= Atocion rupestre (L.) Oxelman): Карелия, берег Онежского оз., мыс Черест, 26 VII 2013, Г.Ю. Конечная (LE); Нюландская губерния, между ст. Оггельбю и Фредериксберг, 15.07.1911, Н.В. Шипчинский 1138 (LE); Finland, Lojo, Rothert, 31.07. (LE); Finland, Lojo, Rothert 2563 (LE). Вид обитает в горных районах Скандинавии, Пиренеев, других горных систем Испании, на Корсике, в Альпах, Апеннинах, в Восточной Европе заходит в Карелию, Ленинградскую и Мурманскую области. Предпочитает известняковые почвы, скалы, трещины кремнистых пород, в Скандинавии встречается также в открытых сосновых лесах (Fraiman et al., 2013).
Изучены 2 вида рода Ixoca Rafin. (Heliosperma (Rchb.) Rchb. Род в обработке разных авторов насчитывает 4–18 видов, распространенных в горах Европы от Пиренеев до Карпат.
I. arcana (Zapał.) Ikonn. (= H. arcanum Zapał.): Ботанический музей БИН РАН, LEc № 9833; Зап. Украина, Станиславская (Ивано-Франковская) обл., 21 VII 1940, А.И. Пояркова и др. (LE). Вид обитает на известняковых скалах в Карпатах, в бассейне Днестра, эндемик (Ikonnikov, 1984).
I. carpatica (Zapał.) Ikonn. (= H. quadrifidum (L.) Reichenb.): Herb. Mus. Univ. Leopoldinensis N 104699, Carpathians, Charnohora, VIII. 1927, J. Mądalski (LE); Иваново-Франковская обл., 28 VI 1963, И.В. Артемчук (LE). Эндемик восточных Карпат, растет на каменистых склонах и скалах (Ikonnikov, 1984).
Изучены 2 вида рода Viscaria Bernh. из 3 (согласно классификации Fraiman et al., 2013).
V. alpina (L.) G. Don.: Кольский п-ов, басс. р. Покой, 29 VIII 60, Н.И. Орлова и др., № 925 (LE); Кольский п-ов, басс. р. Покой, 26 VIII 60, Н.И. Орлова и др., № 850 (LE); ю.-в. часть Кольского п-ва 16 VIII 27, Ю.Д. Цинзерлинг № 1034 (LE); Кольский п-ов, Хибины, 27 VIII 1945, Голубкова №123 (LE); Мурманская губ., Имандра, 39 VII 27, Г. Кренг, Т. Вобликова (LE). Вид встречается на очень различных почвах в тундрах, лесотундрах, высокогорьях, на лужайках, песках и скалах, в горах от Скандинавии до Альп и Пиренеев, а также в Северной Америке, включая восточную Гренландию (Ikonnikov, 1987; Fraiman et al., 2013).
V. viscosa Bernh. (= V. vulgaris Bernh.): Украина, Великолукский уезд, 22 VI 1921, Булавнина и др. (KW); Псковская обл., дер. Опарино, 14 VIII 1915, Кузнецов, № 9832 (LE); Москва, ГБС, А.Н. Швецов. Вид предпочитает песчаные и каменистые почвы, встречается на лугах, скалах, лесных опушках в большинстве районов Европы, в Предкавказье и в Западной Сибири (Ikonnikov, 1987; Fraiman et al., 2013).
Семена исследовали с помощью сканирующего электронного микроскопа Jeol JSM-6390 LA и стереомикроскопа МБС-10. У большинства видов они изучены в 2–3 или более повторностях, по 10 семян из каждого образца. Мы руководствовались результатами исследования изменчивости морфологических признаков семян гвоздичных Кожанчикова (Kozhanchikov, 1975), согласно которым минимальной выборкой для большинства видов могут служить 5–10 семян одного образца. На латеральных сторонах семени наблюдали и измеряли клетки, расположенные концентрическими рядами. В работе принята терминология, используемая при описании морфологии растений и микроморфологии их поверхности (Murley, 1951; Barthlott, 1981; Stern, 2004; Barthlott et al., 2017).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Семена у изученных видов Viscaria group очень мелкие (табл. 1; рис. 1–5), 0.5–0.8 мм дл., 0.4–0.7 мм шир., 0.2–0.5 мм толщ., латерально уплощенные, в плане округло-почковидные, почковидные, иногда неравно-, то есть, асимметрично почковидные и формы запятой (V. alpina), округло-треугольные, на продольно-медианном срезе неправильно узко-трапециевидные (виды рода Atocion), широко-трапециевидные (Minjaevia), обратнояйцевидные, узко-обратнояйцевидные, прямоугольные (Viscaria), в основании слабо выемчатые с выступом (Atocion, Minjaevia) или без него (Viscaria), с вогнутой, ровной или слегка выпуклой дорсальной стороной, коричневые или тёмно-коричневые, у рубчика в основном черноватые, иногда такого же цвета, что и семя (Minjaevia), блестящие (Atocion) или без блеска. Латеральные стороны семени слабо изогнутые или от слабо выпуклых до слабо вогнутых, у Atocion с полукольцевым периферическим ребром; их поверхность шероховатая, сглажено-бугорчатая (Viscaria alpina) или бугорчатая с концентрическими рядами бугорков (V. viscosa). Рубчик без латеральных складок и околорубчиковых выступов (открытый) либо переходного типа, с едва развитыми выступами или складками.
Таблица 1.
Морфология семени у видов Viscaria group (Sileneae) Table 1. Seed morphology in the species of Viscaria group (Sileneae)
| Вид Species |
Длина Length | Ширина Width | Толщина Thickness | Форма Shape | Основание
семени Seed base |
Дорсальная
сторона Dorsal side |
Латеральные
стороны Lateral sides |
Поверхность
латеральных
сторон Surface of lateral sides |
Маргиналь-
ные выступы Marginal projections |
Цвет Color |
Блеск/Shine | Рубчик Hilum |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Atocion armeria | 0.4–0.5 | 0.5–0.7 | 0.3–0.4 | RR | сл. выемчатое
с выступом sl. recessed with projection |
вогнутая concave |
сл. изогнутые или ровные sl.curved/ flat |
шероховатая rough |
ребра ribs |
br/ dark-br, bl | + | открытый/переходный к лат. выступам open/passing to lateral protrusions |
| Atocion lithuanicum | 0.5–0.6 | 0.5–0.6 | 0.3–0.4 | RR | сл. выемчатое
с выступом sl. recessed with projection |
вогнутая concave |
сл. изогнутые или ровные sl. curved/ flat |
шероховатая rough |
ребра ribs |
br, bl | + | открытый/переходный к лат. выступам open/passing to lateral protrusions |
| Minjaevia rupestris | 0.5–0.6 | 0.5–0.7 | 0.3–0.5 | TR | почти ровное
с выступом almost straight with projection |
ровная или
сл. вогнутая flat sl. concave |
ровные flat |
шероховатая rough |
– | br/ dark-br | – | открытый open |
| Viscaria alpina | 0.5–0.7 | 0.6–0.8 | 0.3–0.4 | AR C R RR |
сл. выемчатое sl. recessed |
ровная или
сл. выпуклая flat sl.convex |
сл. выпуклые
или ровные sl. convex/ flat |
сглаженно-
бугорчатая smoothed- tuberculate |
– | br, bl | – | переходящий
от открытого к
лат. складкам passing from open to lateral folds |
| Viscaria viscosa | 0.4–0.5 | 0.5–0.7 | 0.2–0.3 | RR | сл. выемчатое sl. recessed |
сл. вогнутая или ровная sl. concave/flat |
сл. выпуклые
или ровные sl. convex/flat |
бугорчатая tuberculate |
– | br, bl | – | открытый open |
| Ixoca arcana | 0.6–0.7 (1.2–1.3) |
0.9–1.0 (1.4–1.5) |
0.3–0.4 | R RR |
с выступающим валиком with protrud-ing roll |
с коронкой crowned |
сл. выпуклые
или ровные sl. convex/ flat |
шероховатая, ин.
с неотчетливыми бугорками rough smt. with indistinct tubercles |
3 ряда крупных волосков 3 rows of large hairs |
br | + | окружен валиком surrounded by a roll |
| Ixoca
carpatica с коронкой incl. crown |
0.5–0.7 (1.1–1.4) |
0.6–0.8 (1.4–1.6) |
0.2–0.4 | R RR |
с выступающим валиком with protruding roll |
с коронкой crowned |
сл. выпуклые
или ровные sl. convex/ flat |
шероховатая ин.
с неотчетливыми бугорками rough smt. with indistinct tubercles |
3 ряда крупных волосков 3 rows of large hairs |
br | + | окружен валиком surrounded by a roll |
Примечания. Форма семени (очертания): AR – асимметрично-почковидное, C – форма запятой, R – почковидное, RR – округло-почковидное, TR – треугольно-почковидное; цвет: bl – черноватый (возле рубчика), br – коричневый, dark-br – тёмно-коричневый; ин. – иногда, лат. –латеральный, сл. – слабо.
Notes. Seed shape: AR – asymmetrically reniform, C – comma-like, R – reniform, RR – rounded reniform, TR – triangular reniform; color: bl – blackish (near hilum), br – brown, dark-br – dark-brown; sl. – slightly, smt. – sometimes.
Рис. 1.
Внешний вид и фрагменты поверхности семени у Atocion armeria (SEM).
A – семя с латеральной стороны, B – с дорсальной стороны (со спинки), C – с вентральной стороны, D, F – клетки экзотесты (вид сверху) на латеральной стороне семени при разном увеличении, E – клетки экзотесты на спинке, G – поверхность наружной перикалинальной стенки и граница смежных клеток экзотесты; cr – кристалл воска, h – рубчик семени, hr – область рубчика, lf – латеральные складки семенной кожуры около рубчика, p – папиллы на наружной стенке экзотестальной клетки; звездочкой обозначены углубления (?межклетники) в экзотесте. Масштабные линейки (мкм): A, B, D – 100, C – 200, E, F – 50, G – 5.
Fig. 1. SEM micrographs of seed and seed surface in Atocion armeria.
A – seed in lateral view, B – seed in dorsal view (the bark), C – seed in ventral view, D, F – exotesta cells in surface view on the lateral seed side, E – exotesta cells on the bark, G – outer tangential cell wall and cell boundaries in exotesta; cr – wax crystal, h – hilum, hr – hilar region, lf – lateral seed coat folds near hilum, p papilla on the outer periclinal wall of exotesta cell; pores in the exotesta (?intercellular spaces) are marked by asterisk. Scale bars, µm: A, B, D – 100, C – 200, E, F – 50, G – 5.

Рис. 2.
Внешний вид и фрагменты поверхности семени у Atocion lithuanicum.
A – семя с латеральной стороны, B – с дорсальной стороны, C – с вентральной стороны, D, F – клетки экзотесты (вид сверху) на латеральной стороне семени при разном увеличении, E – клетки экзотесты на спинке семени. Обозначения те же, что на рис. 1. Масштабные линейки (мкм): A–C – 100, D–F – 50.
Fig. 2. SEM micrographs of seed and seed surface in Atocion lithuanicum.
A – seed in lateral view, B – seed in dorsal view (the bark), C – seed in ventral view, D, F – exotesta cells in surface view on the lateral seed side, E – exotesta cells on the bark. For abbreviations, see Fig. 1. Scale bars, µm: A–C – 100, D–F – 50.
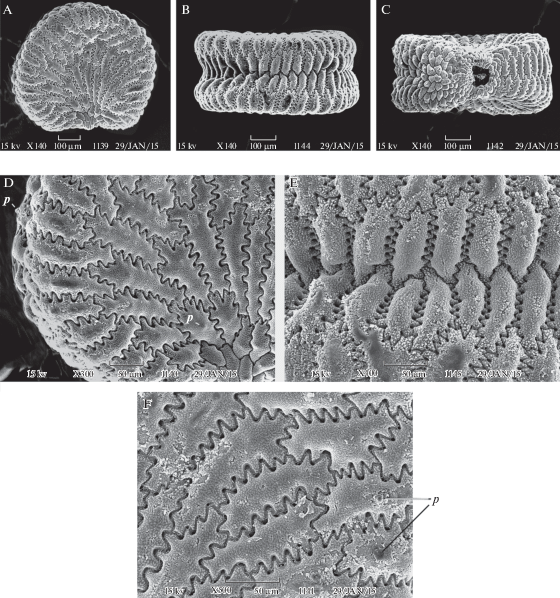
Рис. 3.
Внешний вид и фрагменты поверхности семени у Minjaevia rupestris.
A – семя с латеральной стороны, B – с дорсальной стороны, C – с вентральной стороны, D–G – клетки экзотесты (вид сверху) при разном увеличении: D, E, G – в середине латеральной стороны семени, F – на границе латеральной и дорсальной сторон. Масштабные линейки (мкм): A–C – 100, D, F – 50, E – 20, G – 10.
Fig. 3. SEM micrographs of seed and seed surface in Minjaevia rupestris.
A – seed in lateral view, B – seed in dorsal view, C – seed in ventral view, D–G – exotesta cells in surface view: D, E, G – in the middle of lateral sides, F – on the border of the lateral and dorsal sides. Scale bars, µm: A–C – 100, D, F – 50, E – 20, G – 10.

Рис. 4.
Внешний вид и фрагменты поверхности семени у Viscaria alpina.
A – семя с латеральной стороны, B – с дорсальной стороны, C – с вентральной стороны, D, F – клетки экзотесты (вид сверху) на латеральной стороне семени при разном увеличении, E – клетки экзотесты на дорсальной стороне, G – поверхность наружной тангентальной стенки и граница смежных клеток экзотесты, звездочкой обозначены углубления в экзотесте. Масштабные линейки (мкм): A–C– 100, D, E – 50, F – 10, G – 5.
Fig. 4. SEM micrographs of seed and seed surface in Viscaria alpina.
A – seed in lateral view, B – seed in dorsal view, C – seed in ventral view, D, F – exotesta cells in surface view on the lateral seed side, E – exotesta cells on the bark, G – outer tangential cell wall and cell boundaries in exotesta; pores in the exotesta are marked by asterisk. Scale bars, µm: A–C – 100, D, E – 50, F – 10, G – 5.

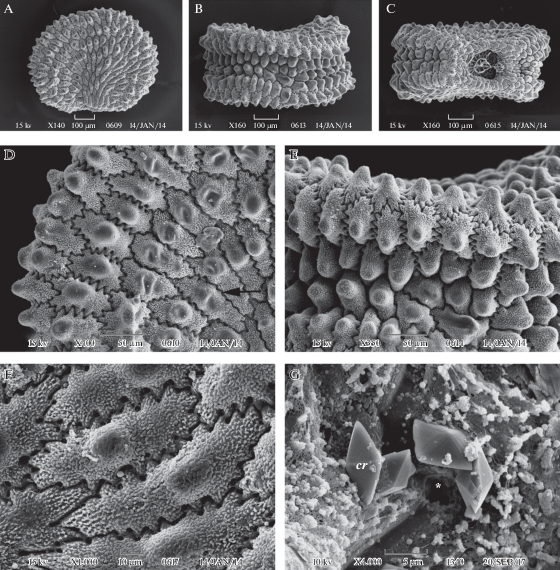
Рис. 5.
Внешний вид и фрагменты поверхности семени у Viscaria viscosa
A – семя с латеральной стороны, B – с дорсальной стороны, C – с вентральной стороны, D, F – клетки экзотесты (вид сверху) на латеральной стороне семени при разном увеличении, E – клетки экзотесты на дорсальной стороне, G – поверхность наружной тангентальной стенки и граница смежных клеток экзотесты; cr – кристалл воска, звездочкой обозначены углубления в экзотесте, стрелкой – слабоволнистые антиклинальные клеточные стенки. Масштабные линейки (мкм): A–C – 100, D, E – 50, F – 10, G – 5.
Fig. 5. SEM micrographs of seed and seed surface in Viscaria viscosa.
A – seed in lateral view, B – seed in dorsal view, C – seed in ventral view, D, F – exotesta cells in surface view on the lateral seed side, E – exotesta cells on the bark, G – outer tangential cell wall and cell boundaries in exotesta; cr – wax crystal, pores in the exotesta are marked by asterisk, and weakly waved anticlinal cell walls – by arrow. Scale bars, µm: A–C – 100, D, E – 50, F – 10, G – 5.
Семенная кожура (табл. 2; рис. 1–5) экзотестальная, со склерифицированной экзотестой. Ее первичная скульптура на латеральных сторонах семени радиально прерывисто-струйчатая (рис. 1A, 2A, 3A), бугорчато радиально-струйчатая (рис. 5A) или сходная с коликулярной (colliculate) (рис. 4A). Клетки экзотесты на латеральных сторонах семени большей частью продольно удлиненные и расположены 2–5 концентрическими рядами (у Atocion lithuanicum ряды иногда местами сбиты), у Viscaria alpina продольная ориентация клеток и концентрические ряды отсутствуют. Клетки различной формы: звездчатые (рис. 7A), удлиненно-звездчатые (рис. 7B), сердцевидные (рис. 7C), продолговатые (рис. 7D), линейные (= гусеницевидные) (рис. 7E), стреловидные (рис. 7F), достигают наибольшей длины у Atocion armeria (220 мкм), наиболее короткие (до 105 мкм) у Viscaria alpina. Их наружная периклинальная стенка выпуклая до полусферической, иногда с центральным закругленно-коническим или мамиллятно-коническим бугорком, либо сосочком, тонко cкульптурированная: вторичная скульптура зернистая (гранулированная) или сглажено-зернистая. На поверхности наружных стенок у большинства видов найдены структуры воска в форме кристаллов и друз (рис. 1G, 5G). Антиклинальные стенки этих клеток извилистые с 11‒25 зубцами: S-волнистые (рис. 2F), V-волнистые или зубчатые (рис. 1D), слабо волнистые (рис. 5D, обозначены стрелкой), волнообразные (широко-волнистые – рис. 7E). В одной клетке могут сочетаться S- и V- типы волнистости, а на одном семени иногда встречается большее число типов.
Таблица 2.
Микроморфология семенной кожуры у видов Viscaria group (Sileneae) Table 2. Seed coat micromorphology in members of Viscaria group (Sileneae)
| Вид species |
Первичная скульптура Primary sculpture |
Клетки экзотесты на латеральных сторонах Exotesta cells on lateral sides |
Клетки экзотесты на дорсальной стороне Exotesta cells on dorsal side |
Клетки экзотесты на границе дорсальной и латеральной сторон Exotesta cells on the border of the dorsal and lateral sides | Структуры воска Wax structures |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Число рядов Number of cell rows |
Основная форма Main shape |
Максимальная длина (мкм)/ max length (µm) | Наружная пери-
клинальная стенка Outer periclinal wall |
Вторичная скульптура Secondary sculpture |
Антиклиналь-
ные стенки Anticlinal walls |
Число зубцов Number of teeth |
Число рядов Number of cell rows |
Форма Shape |
Ориентация Orientation |
Наружная перикли-
нальная стенка Outer periclinal wall |
||||
| Atocion armeria | радиально
прерывисто- струйчатая radially discon- tinuous-striate |
3(4) | Ob, L, Sa | 220 | C/H ин. с сосочком C/H smt. with papilla |
Gr | S, V | 16–25 | 3–5 | Ov to Ob | поперек спинки transverse to back |
C/H ин. с сосочком C/H smt. with papilla |
1 согнутая 1 bent |
cr, d |
| Atocion lithuanicum | неясно радиально струйчатая indistinctly radially discon- tinuous-striate |
3(2) | Ob, L, Sa | 190 | C/H ин. с сосочком C/H smt. with papilla |
Gr/ smGr | S, V | 19–24 | 3–5 | Ob | поперек спинки transverse to back |
C/H ин. с сосочком C/H smt. with papilla |
1 согнутая 1 bent |
– |
| Minjaevia rupestris | радиально
струйчатая radially discon- tinuous striate |
2–3 | Ob to L | 160 | C | Gr | S, WS, реже V | 17–21 | 4 | Ob | поперек спинки transverse to back |
C/H | 1 согнутая 1 bent |
s.cr |
| Viscaria alpina | сходная с коликулярной similar to colliculate |
– | St | 105 | H | Gr | S, реже V и WS S, less often V and WS | 11–17 | – | St | – | H | 2 разно-
направленные 2 divergent |
– |
| Viscaria viscosa | бугорчато-
радиально-
струйчатая tuberculate radially-striate |
4–5 | Ob, ESt | 115 | C с закругленно-коническим бугорком C with rounded- conical tubercle |
Gr | V, WU, WS | 19–25 | 5–6 | 2 типов:
1. Ob с ц. бугорком, 2. IP клетка-бугорок 2 types: 1. Ob with c. tubercle, 2. cell-tubercle |
– | выпуклая с ц. мамиллятным бугорком или вся вытянута в такой бугорок C with c. mamillate tubercle/all cell stretched out in such a tubercle |
2 разно-
направленные 2 divergent |
cr |
| Ixoca arcana | пазловидная jigsaw puzzle-like |
1–3 | A | 190 | C | Gl | L | 9–17 | 3 | плоские волоски flat hairs |
– | вытянута
в волосок stretched into a hair |
обычные
или с бугорком usual/with tubercle |
s.cr |
| Ixoca carpatica | пазловидная jigsaw puzzle-like |
1–3 | A | 180 | C | Gl | L | 13–17 | 3 | плоские волоски flat hairs |
– | вытянута
в волосок stretched into a hair |
обычные или с бугорком usual/with tubercle | s.cr |
Примечания. Основная форма (очертания): A – амебовидная, E St – удлиненно-звездчатая, IP – неправильно-полигональная, L – линейная, Ob – продолговатая, Ov – овальная, Sa – стреловидная; St – звездчатая; наружная периклинальная стенка: С – выпуклая, H – полусферическая; вторичная скульптура: Gr – зернистая, smGr – сглаженно-зернистая, Gl – комковатая; антиклинальные клеточные стенки: L – лопастные, S – S-волнистые, V – V-волнистые (зубчатые), WS – широко S-волнистые, WU – слабоволнистые; структуры воска: cr – кристаллы, d – друзы, s.cr – единичные кристаллы; ин. – иногда, ц. – центральный.
Notes. Main cell shape (outlines): A – amoeboid, E St – elongated stellate, IP – irregularly polygonal, L – linear, Ob – oblong, Ov – oval, Sa – sagittate, St – stellate; оuter periclinal wall: C – convex, H – hemispherical; secondary sculpture: Gr – granulate, smGr – smooth-granulate, Gl –glebulate; anticlinal cell walls: L – lobate, S – S-undulate, V – V-undulate (dentate), WS – widely S-undulate, WU – weakly undulate; wax structures: cr – crystals, d – druses, s.cr – solitary crystals; c. – central, smt. – sometimes.
Рис. 6.
Внешний вид и фрагменты поверхности семени у Ixoca arcana (A–D, F) и I. carpatica (E, G).
A – семя с латеральной стороны, B – с дорсальной стороны (волоски срезаны), C – с вентральной стороны, D–F – клетки экзотесты (вид сверху) на латеральной стороне семени при разном увеличении, G – поверхность наружной тангентальной стенки и граница смежных клеток экзотесты; c коронка из волосков, h rim – околорубчиковый валик. Масштабные линейки (мкм): A – 200, C – 100, B, D – 50, E, F – 20, G – 10.
Fig. 6. SEM micrographs of seed and seed surface in Ixoca arcana (A–D, F) и I. carpatica (E, G).
A – seed in lateral view, B – seed in dorsal view (crown hairs are cut off), C – fragment of seed in ventral view, D–F – exotesta cells in surface view on the lateral seed side, G – outer tangential cell wall and cell boundaries in exotesta; c crown of hairs, h rim – hilar rim. Scale bars, µm: A – 200, C – 100, B, D – 50, E, F – 20, G – 10.
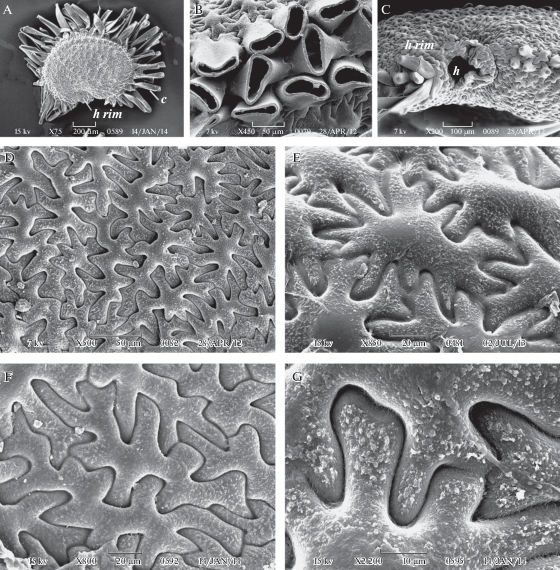
Рис. 7.
Клетки экзотесты на латеральных сторонах семени у некоторых видов трибы Sileneae (Viscaria group).
A – звездчатая (Viscaria alpina), B – удлиненно-звездчатая (V. alpina), C – сердцевидная (V. alpina), D – продолговатая (Minjaevia rupestris), E – линейная или гусеницевидная (Atocion lithuanicum), F – стреловидная (A. lithuanicum), G – звездчато-амебовидная (Ixoca arcana), H – удлиненно-амебовидная (I. arcana). Масштабная линейка 50 мкм.
Fig. 7. Exotesta cells on lateral seed side of certain Sileneae species (clade Viscaria group).
A – stellate (Viscaria alpina), B – elongated-stellate (V. alpina), C – cordate (V. alpina), D – oblong (Minjaevia rupestris), E – linear or larva-shaped (Atocion lithuanicum), F – sagittate (A. lithuanicum), G – stellate-ameboid (Ixoca arcana), H – elongated-ameboid (I. arcana). Scale bar 50 µm.

На дорсальной стороне семени клетки экзотесты от овальных до продолговатых, редко звездчатые, расположены 3–5 рядами (кроме V. alpina: у этого вида четкие ряды клеток отсутствуют). У Atocion и Minjaevia клетки ориентированы поперек спинки, тогда как у видов рода Viscaria они не имеют определенной ориентации. Наружная периклинальная стенка более выпуклая, чем на латеральных сторонах семени, полусферическая или гребневидная (Atocion, Minjaevia), у Viscaria viscosa с крупным центральным мамиллятным бугорком или вся вытянута в широкий мамиллятный бугорок (рис. 5F).
Вдоль более или менее отчетливых ребер на границе латеральных и дорсальной сторон семени проходит либо один ряд длинных согнутых экзотестальных клеток (рис. 1E, 3F), либо два ряда коротких разнонаправленных клеток (рис. 5E). У V. alpina ребра семени неотчетливые (рис. 4B, D).
Семена Ixoca благодаря морфологическим особенностям иногда выделяют в особый тип (Ghazanfar, 1983). У изученных нами видов (рис. 6A–F) они резко отличаются от семян в Viscaria group по нескольким признакам: 1) наличие крупных лучеобразных волосков на дорсальной стороне семени (рис. 6A), за что этот род получил название Heliosperma (Reichenb.) Reichenb. – солнцесемянник; плоские жесткие волоски развиваются из ориентированных различным образом клеток экзотесты, расположенных на спинке семени в 3 ряда; 2) выступающий в основании семени и ограничивающий рубчик валик (по-видимому, производное фуникулуса); 3) часто асимметричное положение рубчика; 4) необычная амебовидная форма клеток экзотесты, с лопастными антиклинальными стенками (рис. 7G, H; рис. 6D–E); они расположены в основном неупорядоченно, иногда сильно вытянуты вдоль семени, и лишь по периферии латеральных сторон семени находятся в 1–3 концентрических рядах, более отчетливых и широких у некоторых семян I. carpatica; 4) комковатая (glebulate), а не зернистая, вторичная скульптура наружной периклинальной стенки этих клеток – из мелких, в основном редко расположенных комочков, местами слившихся и уплощенных (рис. 6G). Нужно отметить, что клетки экзотесты такой формы, расположенные хаотично, не встречены нами более ни у одного изученного таксона из трибы Sileneae, однако имеются у представителей трибы Alsineae Lam. et DC.: некоторых видов родов Mesostemma Vved. (Arabi et al., 2017), Stellaria L. (Mahdavi et al., 2012; Bozchaloyi, Keshavarzi, 2014), Moehringia L. (Minuto et al., 2006), а также у видов рода Spergularia (Pers.) J. Presl et C. Presl из трибы Paronychieae Dumort. (Abdel-Maksoud, Fawzi, 2016). Полученные нами результаты соответствуют, таким образом, результатам молекулярно-филогенетических исследований, показавших обособленность рода Ixoca в трибе Sileneae. Этот вывод подтверждают также данные по анатомии плодов (Weberbauer, 1898; Kravtsova, Bolotova, 2019). Кроме того, обращает на себя внимание точное повторение характерного признака семян Ixoca – коронки из нескольких рядов плоских волосков по спинке семени у некоторых видов рода Minuartia L. (Alsineae) – M. inamoena (C.A. Mey.) Woronow, M. rhodocalyx (Albov) Woronow (Zaychenko, Zernov, 2017) и M. macrocarpa var. koreana (Nakai) H. Hara (Song et al., 2015).
Полученные результаты показывают, что для Atocion armeria и A. lithuanicum характерны округло-почковидные, значительно уплощенные семена, отличительными признаками которых являются желобок на дорсальной стороне, крупные ребра по периферии латеральных сторон, экзотеста из сильно удлиненных клеток без бугорков (характерных также для многих видов рода Silene), иногда с центральным мелким сосочком. Семенная кожура на границе дорсальной и латеральных сторон образована 1 рядом согнутых клеток. Различия между этими видами рода Atocion по величине семян незначительные: семена у A. armeria несколько шире (до 700 мкм), чем семена у A. lithuanicum (до 600 мкм), но менее длинные (высокие). Эти результаты расходятся с данными V. Martynyuk et al. (2015), согласно которым семена A. lithuanicum мельче, чем у A. armeria по обоим параметрам. Нет соответствия и в описании формы семян: так, мы не нашли у A. armeria треугольно-почковидных семян, указываемых этими авторами. Совпадают наблюдения, касающиеся межвидовых различий по величине клеток экзотесты: у A. armeria они длиннее, чем у A. lithuanicum на латеральных сторонах семени. Различия между видами по величине папилл на наружной периклинальной стенке этих клеток на нашем материале не выявлены, возможно, в связи с отсутствием достаточного количества материала. Нужно отметить, что проведенное в цитируемой работе разграничение видов рода Atocion по микроморфологии семян основано большей частью на количественных признаках, которые значительно перекрываются у A. armeria и A. lithuanicum.
Особенностями семян Minjaevia rupestris, очень сходных по микроморфологии семенной кожуры с видами рода Atocion, являются их значительная толщина, отсутствие желобка на спинке и крупных ребер по периферии латеральных сторон. Мы считаем, что в иерархии признаков семени признак формы клеток экзотесты занимает первое место, а признаки толщины семени, наличия или отсутствия желобка по спинке имеют в данном случае менее важное таксономическое значение, позволяя диагностировать виды.
Семена изученных видов рода Viscaria значительно различаются по микроморфологии. Можно выделить 2 их типа: в типе V. alpina семена более крупные, чем у V. viscosa, обычно менее округлые в очертании, несколько асимметричные, с широкой, ровной или слегка выпуклой спинкой без ложбинки, ребра не выражены; поверхность по всему семени сглажено-бугорчатая, сходная с коликулярной (colliculate). Семенная кожура имеет примитивные черты: она слабо дифференцирована, клетки экзотесты одинаковой формы на латеральных и дорсальной сторонах, расположены хаотично, не собраны в ряды.
В типе V. viscosa семена мельче, почти округлые в очертании, более симметричные и плоские, чем у V. alpina, иногда с желобком на дорсальной стороне; поверхность бугорчатая с правильными рядами бугорков – концентрическими на латеральных сторонах и продольными по спинке. Клетки экзотесты различаются по форме не только на разных сторонах семени, но и в разных рядах на дорсальной стороне: на латеральных сторонах семени они продолговатые с закругленно-коническим бугорком, в крайних рядах на спинке такие же с мамиллятно-коническим бугорком, в двух средних рядах спинки представляют собой такой бугорок на звездчатом основании, вся их наружная стенка сильно вытянута.
Выявление двух типов семени в роде Viscaria согласуется с его делением на 2 секции (Ikonnikov, 1987): V. alpina была выделена в отдельную секцию Liponeurum (Schott, Nym. et Kotschy) Ikonn.
Разнотипные семена обнаружены также у близких видов многих секций рода Silene s.l. (Candáu, Talavera, 1978; Ghazanfar, 1983; Ocaña et al., 2011; Hoseini et al., 2017). При этом различия между разными типами семян по их микроморфологии более или менее выражены. Резко разнотипные семена найдены в секции Erectofractae Chowdhuri (Candáu, Talavera, 1978; Ocaña et al., 2011), у некоторых видов они аномальные: экзотеста образована не характерными для трибы Sileneae мелкими, неправильно-полигональными, хаотично расположенными клетками.
Семена Viscaria обоих типов заметно отличаются от семян Atocion и Minjaevia по микроморфологии семенной кожуры: клетки экзотесты у Viscaria более короткие, нередко звездчатые, с коническим бугорком или сильно выпуклой (полусферической) наружной периклинальной стенкой; семенная кожура на ребре (если имеется) из 2 рядов разнонаправленных клеток. Сходные очертания клеток экзотесты, наличие у них конического бугорка, присутствие на спинке семени “клеток-бугорков”, вся наружная стенка которых вытянута в крупный бугорок, сближает род Viscaria с родом Lychnis. Ранее сходство скульптурных элементов семенной кожуры у Viscaria и Lychnis отметил Кожанчиков (Kozhanchikov, 1967). Немногими общими признаками семени у родов Viscaria group являются очень небольшой размер семян (не превышает 0.7 мм в длину, 0.8 мм в ширину); в основном открытый рубчик без хорошо развитых околорубчиковых латеральных складок и выступов (остатков фуникулуса по предположению Романовой и Кравцовой (Romanova, Kravtsova, 2016); заметные “поры” на семенной кожуре, представляющие собой, по-видимому, межклетники в экзотесте; наличие структурированных эпикутикулярных восковых отложений на поверхности клеток экзотесты в виде кристаллов.
ЗАКЛЮЧЕНИЕ
Полученные результаты свидетельствуют, на наш взгляд, о том, что микроморфология семени в исследуемой группе имеет таксономическое и диагностическое значение. Наиболее таксономически важными признаками являются форма и ориентация клеток экзотесты, наличие или отсутствие, а также форма бугорка на наружной периклинальной стенке.
Проведенное исследование подтвердило обособленное положение рода Ixoca в трибе Sileneae. Кроме коронки из волосков семена их видов отличаются от семян в Viscaria group необычной амебовидной формой в основном неупорядоченно расположенных клеток экзотесты, другой скульптурой их наружной периклинальной стенки, наличием валика вокруг рубчика, обычно асимметрично расположенного.
Близкое родство Atocion и Minjaevia, выявленное молекулярными методами, подтверждается сравнительно-морфологическим изучением семян.
Результаты расходятся с молекулярными данными о сестринских отношениях родов Atocion и Viscaria. Выявленные у Viscaria 2 типа семени (V. alpina и V. viscosa) значительно различаются по микроморфологии; оба типа заметно отличаются от семян Atocion и Minjaevia. Таким образом, полученные нами результаты лишь частично соответствуют филогении рассматриваемой группы (Viscaria group), основанной на молекулярных данных. Они расходятся с этими данными в основном вопросе – родственных отношениях Atocion и Viscaria. Мы предполагаем, что расхождение морфологических и молекулярных исследований может быть связано с выявленным своеобразием рода Viscaria, которое проявляется в разнотипности семян у изученных видов, а также в особенном вскрывании плодов (primarily loculicidal, по Friman et al., 2013), не встречающемся в других родах трибы.
Список литературы
Abdel-Maksoud H.S., Fawzi N.M. 2016. Seed morphology of some taxa of Caryophyllaceae. – Bull. Fac. Agric., Cairo Univ. 67 (3): 213–227.
Aeschimann D. 1984. Étude biosystematique du Silene vulgaris s. l. (Caryophyllaceae) dans de domaine alpin. Morphologie de la graine. – Candollea. 39 (1): 135–149.
Arabi Z., Ghahremaninejad F., Rabeler R.K., Heubl G., Zarre Sh. 2017. Seed micromorphology and its systematic significance in tribe Alsineae (Caryophyllaceae). – Flora. 234: 41–59. https://doi.org/10.1016/j.flora.2017.07.004
Arman M., Gholipour A. 2013. Seed morphology diversity in some Iranian endemic Silene (Caryophyllaceae) species and their taxonomic significance. – Acta Biol. Szeged. 57 (1): 31–37.
Atazadech N., Keshavarzi M., Sheidai M., Gholipour A. 2017. Seed morphology of Silene commelinifolia Boiss. complex (Caryophyllaceae Juss.). – Modern Phytomorphology. 11: 5–13. https://doi.org/10.5281/zenodo.34558
Barthlott W. 1981. Epidermal and seed surface characters of plants: systematic applicability and some evolutionary aspects. – Nord. J. Bot. 1 (3): 345–355.
Barthlott W., Mail M., Bhushan B., Koch K. 2017. Plant Surfaces: Structures and Functions for Biomimetic Innovations. – Nano-Micro Lett. 9: 23. https://doi.org/10.1007/s40820-016-0125-1
Boissier E. 1867. Flora Orientalis. Geneve. Vol. 1. 1017 p.
Bojňanský V., Fargašová A. 2007. Atlas of Seeds and Fruits of Central and East-European Flora: the Carpathian Mountains Region. Dordrecht: Springer Netherlands. 1046 p.
Bozchaloyi S.E., Keshavarzi M. 2014. Micro- and macromorphological study of Stellaria (Caryophyllaceae) and its closest relatives in Iran. – Phytologia Balcanica. 20 (2): 179–197.
Camelia I. 2011. Aspects regulating seeds morphology and germination peculiarities at some taxa from Silene L. genera. – J. Plant Develop. 18: 5–10.
Candáu P., Talavera S. 1978. Polen y semillas de las especies de Silene, sect. Erectofractae Chowdhuri. – Lagascalia 8 (2): 127–133.
Chowdhuri P.K. 1957. Studies in the genus Silene – Notes Roy. Bot. Gard. Edinburg. 22: 221–278.
Dadandi M.Y., Yildiz K. 2015. Seed morphology of some Silene L. (Caryophyllaceae) species collected from Turkey. – Turk. J. Bot. 39: 280–297. https://doi.org/10.3906/bot-1307-35
Erixon P., Oxelman B. 2008. Reticulate or tree-like chloroplast DNA evolution in Sileneae (Caryophyllaceae)? – Mol. Phylogen. Evol. 48 (1): 313–325. https://doi.org/10.1016/j.ympev.2008.04.015
Fawzi N., Fawzy A., Mohamed A. 2010. Seed morpho-logical studies on some species of Silene L. (Caryophyllaceae). – Int. J. Bot. 6 (3): 287–292. https://doi.org/10.3923/ijb.2010.287.292
Frajman B., Oxelman B. 2007. Reticulate phylogenetics and phytogeographical structure of Heliosperma (Sileneae, Caryophyllaceae) inferred from chloroplast and nuclear DNA sequences. – Mol. Phylogen. Evol. 43 (1): 140–155. https://doi.org/10.1016/j.ympev.2006.11.003
Frajman B., Eggens F., Oxelman B. 2009a. Hybrid origins and homoploid reticulate evolution within Heliosperma (Sileneae, Caryophyllaceae) – a multigene phylogenetic approach with relative dating. – Syst. Biol. 58 (3): 328–345. https://doi.org/10.1093/sysbio/syp030
Frajman B., Heidary N., Oxelman B. 2009b. Phylogenetic relationships of Atocion and Viscaria (Sileneae, Caryophyllaceae) inferred from chloroplast, nuclear ribosomal and low-copy gene DNA sequences. – Taxon. 58 (3): 811–824. https://doi.org/10.1002/tax.583010
Frajman B., Thollesson M., et Oxelman B. 2013. Taxonomic revision of Atocion and Viscaria (Sileneae, Caryophyllaceae). – Bot. J. Linn. Soc. 173 (2): 194–210. https://doi.org/10.1111/boj.12090
Ghazanfar S.A. 1983. Seed characters as diagnostic in the perennial sections of the genus Silene L. (Family Caryophyllaceae) 1. – Pak. J. Bot. 15 (1): 7–12.
Gholipour A., Kuhdar F. 2014. Seed morphology diversity in the certain Silene species (Caryophyllaceae) from Iran and its taxonomic significance. – Taxonomy and Biosystematics. 6 (18): 107–118.
Greuter W. 1995. Silene (Caryophyllaceae) in Greece: a subgeneric and sectional classification. – Taxon. 44 (4): 543–581. https://doi.org/10.2307/1223499
Hong S.P., Han M.J., Kim K.J. 1999. Systematic significance of seed coat morphology in Silene L. s. str. (Sileneae – Caryophyllaceae) from Korea. – J. Plant Biol. (Korea). 42: 146–150. https://doi.org/10.1007/BF03031023
Hoseini E., Ghahremaninejad F., Assadi M., Edalatiyan M.N. 2017. Seed micromorphology and its implication in subgeneric classification of Silene (Caryophyllaceae, Sileneae). – Flora. 228: 31–38. https://doi.org/10.1016/j.flora.2017.01.006
[Ikonnikov] Иконников С.С. 1984. Заметки о гвоздичных (Caryophyllaceae). 7. – Новости сист. высш. раст. 21: 61–67.
[Ikonnikov] Иконников С.С. 1987. Заметки о гвоздичных (Caryophyllaceae). 8. – Новости сист. высш. раст. 24: 79–85.
Jafari F., Zare S., Gholipour A., Eggens F., Rabeler R.K., Oxelman B. 2020. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). – Taxon. 69 (2): 337–368. https://doi.org/10.1002/tax.12230
Jeanmonod D. 1985. Révision de la section Siphonomorpha Otth du genre Silene L. (Caryophyllaceae) en Mediterranée occidentale V: Synthèse. – Candollea. 40 (1): 35–56.
Keshavarzi M., Mahdavinejad M., Sheidai M., Gholipour A. 2015. Seed and pollen morphology of some Silene species (Caryophyllaceae) in Iran. – Phytol. Balc. 21 (1): 7–12.
[Kovtonjuk] Ковтонюк Н.К. 1995. Структура поверхности семян сибирских видов рода Gastrolychnis (Caryophyllaceae) в связи с систематикой. – Бот. журн. 80 (9): 98–101.
[Kozhanchikov] Кожанчиков В.И. 1967. Морфологические признаки семян семейства Caryophyllaceae и возможные пути их эволюции. – Бот. журн. 52 (9): 1277–1286.
[Kozhanchikov] Кожанчиков В.И. 1969. Морфолого-географическое исследование семян представителей сем. Caryophyllaceae Juss. европейской части СССР: Дис. … канд. биол. наук. Л. 293 с.
[Kozhanchikov] Кожанчиков В.И. 1975. Изменчивость морфологических признаков семян представителей сем. Caryophyllaceae Juss. – В кн.: Вопросы сравнительной морфологии семенных растений. Л. С. 108–138.
Kravtsova T.I., Bolotova Ya.V. 2019. Pericarp structure in some species in the tribe Sileneae DC. (Caryophyllaceae, Viscaria group). – Botanica Pacifica. 8 (2): 25–34. https://doi.org/10.17581/bp.2019.08211
Kuh M., Yildiz K., Minareci E. 2017. A taxonomic study of the Silene sections Behenantha and Dichotomae (Caryophyllaceae) in Turkey based on the micromorphology of their seed and pollen. – Turk. J. Bot. 41 (5): 493–504. https://doi.org/10.3906/bot-1610-23
Mahdavi M., Assadi M., Fallahian F., Nejadsattari T. 2012. The systematic significance of seed micromorphology in Stellaria L. (Caryophyllaceae) and its closest relatives in Iran. – Iran J. Bot. 18 (2): 302– 310.
[Martynyuk] Мартинюк В.О., Карпенко Н.І., Царенко О.М. 2015. Деякі мікроморфологічні особливості Atocion lithuanicum (Zapał.) Tzvel. та A. armeria (L.) Raf. Флори України. – Биологический вестник Мелитопольского гос. педагогического ун-та им. Богдана Хмельницкого. 5 (1): 8–23. https://doi.org/10.7905/bbmspu.v5i1.901
Melzheimer V. 1977. Biosystematische revision einiger Silene-Arten (Caryophyllaceae) der Balkanhalbinsel (Griechenland). – Bot. Jahrb. Syst. 98 (1): 1–92.
Melzheimer V. 1987. On the taxonomic position of Silene thebana (Caryophyllaceae). – J. Plant Syst. Evol. 155 (1/4): 251–256. https://doi.org/10.1007/BF00936302
Melzheimer V. 1988. Caryophyllaceae. – In: Flora Iranica. 163: 353–508.
[Mikhailova] Михайлова Ю.В. 2016. Исследование ДНК для решения вопросов систематики Sileneae DC. (Caryophyllaceae Juss.). – В кн.: Проблемы ботаники Южной Сибири и Монголии: сборник научных статей по материалам XV международной научно-практической конференции (23–26 мая 2016 г., Барнаул). Барнаул. С. 153–156.
Minuto L., Fior S., Roccotiello E., Casazza G. 2006. Seed morphology in Moehringia L. and its taxonomic significance in comparative studies within the Caryophyllaceae. – Plant Syst. Evol. 262 (3–4): 189–208. https://doi.org/10.1007/s00606-006-0458-1
Murley M.R. 1951. Seeds of the Cruciferae of Northeastern North America. – Am. Midl. Nat. 46: 1–81. https://doi.org/10.2307/2421948
[Nersesian] Нерсесян А.А. 1990. К изучению поверхности семян некоторых видов рода Silene L. (Caryophyllaceae) флоры Армении. – В кн.: Труды Ленингр. молодёжной конференции ботаников. Л. Т. 1. С. 85–94.
Ocaña M.E., Juan R., Fernández I., Pastor J. 2011. Estudio morfológico de semillas de Silene (Caryophyllaceae) del suroeste de España. – Lagacalia. 31: 21–45.
Oxelman B., Lidén M. 1995. Generic boundaries in the tribe Sileneae (Caryophyllaceae) as inferred from nuclear rDNA sequences. – Taxon. 44 (4): 525–542.
Oxelman B., Lidén M., Berglund D. 1997. Chloroplast rps16 intron phylogeny of the tribe Sileneae (Caryophyllaceae). – Plant Syst. Evol. 206 (1–4): 393–410.
Oxelman B., Lidén M., Rabeler R.K., et Popp M. 2001. A revised generic classification of the tribe Sileneae (Caryophyllaceae). – Nord. J. Bot. 20 (6): 743–748. https://doi.org/10.1111/j.1756-1051.2000.tb00760.x
Perveen A. 2009. Seed morphology of the genus Silene: Caryophyllaceae from Pakistan and Kashmir. – Int. J. Biol. Biotech. 6 (4): 219–227.
[Romanova, Kravtsova] Романова В.О., Кравцова Т.И. 2016. Морфологические особенности области рубчика у семян представителей трибы Sileneae (Caryophyllaceae). – Бот. журн. 101 (2): 189–205. https://doi.org/10.1134/S0006813616020058
[Romanova, Kravtsova] Романова В.О., Кравцова Т.И. 2019. Морфология эпикутикулярных отложений воска на поверхности семенной кожуры в трибе Sileneae (Caryophyllaceae). – Turczaninowia. 22 (1): 164–184. https://doi.org/10.14258/turczaninowia.22.1.16
Rorhbach P. 1868. Monographie der Gattung Silene. Leipzig. 249 p.
Song J.-H., Kim J.-M., Ok M.-K., Hong S.-P. 2015. Comparative seed morphology of the tribe Alsineae (Caryophyllaceae) in Korea and its taxonomic implications. – Korean J. Plant Taxon. 45 (4): 369–379. https://doi.org/10.11110/kjpt.2015.45.4.369
Stern W.T. 2004. Botanical Latin. 4th ed. Portland. 546 p.
Tabaripour R., Koohdar F., Sheidai M., Gholipour A. 2013. Intra-specific variations in Silene: Morphometry and micromorphometry analyses. – Afr. J. Biotechnol. 12 (33): 5208. https://doi.org/10.5897/AJB12.188
[Tzvelev] Цвелев Н.Н. 2001. О родах трибы смолевковых (Sileneae DC., Caryophyllaceae) в Восточной Европе. – Новости сист. высш. раст. 33: 90–113.
Weberbauer A. 1898. Beiträge zur Anatomie der Kapselfrüchte. – Botanisches Centralbatt. 73: 54–59, 97–105, 135– 142, 161–168, 193–202, 250–257, 296–302.
Yildiz K. 2002. Seed morphology of Caryophyllaceae species from Turkey (North Anatolia). – Pak. J. Bot. 34 (2): 161–171.
Yildiz K., Cirpici A. 1998. Seed morphological studies of Silene L. from Turkey. – Pak. J. Bot. 30 (2): 173–188.
Zareh M.M. 2005. Seed diversity among certain species of Caryophyllaceae in Egypt. – Pak. J. Biol. Sci. 8 (5): 714–720. https://doi.org/10.3923/pjbs.2005.714.720
Zaychenko S.G., Zernov A.S. 2017. Structural features of the seed coat in Caucasian representatives of Minuartia (Caryophyllaceae). – Wulfenia. 24: 205–220.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал