

Ботанический журнал, 2022, T. 107, № 12, стр. 1166-1192
ОСОБЕННОСТИ СТРОЕНИЯ СЕМЯНОК ПРЕДСТАВИТЕЛЕЙ ПОДСЕКЦИИ NORVEGICAE СЕКЦИИ ABROTANUM РОДА ARTEMISIA (ASTERACEAE)
О. В. Яковлева 1, *, А. А. Коробков 1, А. Н. Иванова 1
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
* E-mail: yakovleva@binran.ru
Поступила в редакцию 12.09.2022
После доработки 31.10.2022
Принята к публикации 01.11.2022
- EDN: WEZGNE
- DOI: 10.31857/S0006813622120122
Аннотация
Исследованы семянки 7 видов Artemisia, относящихся к подсекции Norvegicae секции Abrotanum, включая 4 подвида A. arctica. Методы исследования: световая, сканирующая и просвечивающая электронная микроскопия. У семянок представителей подсекции Norvegicae отсутствует паппус, на площадке прикрепления венчика имеются устьица, вероятно, относяциеся к нектарникам, карпоподиум хорошо выражен. В центре карпоподиума часто обнаруживаются выступающие проводящие пучки. Ребра на поверхности перикарпия выражены слабо. Их количество колеблется от (4) 5 до 7 (8). По данному признаку виды и подвиды мало различаются между собой. Ребра состоят из крупных секреторных каналов, тонкостенных паренхимных клеток и проводящих пучков. В ребрах отсутствуют гидроциты. Механические клетки в небольшом количестве встречаются в проводящих пучках. В экзокарпии, наряду с основными клетками, находятся удлиненные сегментированные клетки, чаще называемые комплексами слизесодержащих клеток. Сегментированные клетки немногочисленные, разной длины и бывают заполнены не только слизью, но и маслянистым секретом. Виды и подвиды различаются по наличию комплексов слизесодержащих клеток и способности их наружных стенок к “ослизнению”. В слизи клеток слизесодержащих комплексов имеются стопки фибрилл, но они не образуют подковообразные структуры, характерные для представителей из других секций рода Artemisia. Перикарпий в промежутках между ребрами двухслойный. Его клетки в разной степени облитерированы, чаще облитерируются клетки второго внутреннего слоя. Спермодерма экзотестальная, хорошо сохранена. Эндосперм двухслойный.
Род Artemisia является самым крупным и одним из трудных в систематическом отношении родов трибы Anthemideae. По мнению И.М. Крашенинникова (Krasheninnikov, 1946), “самой характерной чертой рода Artemisia является огромный внутривидовой полиморфизм, та неустойчивость, изменчивость морфологических признаков, которая и обусловливает разноречивость в понимании объема многих видов у различных авторов, занимающихся вопросами систематики рода. Вероятно, это же было одной из причин, почему за последнее столетие не появлялось полной монографии рода в целом, подводящей итоги всего накопленного, очень обширного фактического материала” (Krasheninnikov, 1946: 87).
Морфологическое и анатомическое строение семянок изучалось лишь у немногих видов полыней (Korobkov, 1973, 1981; Ouyahya, Viano, 1990; Ouyahya, 1995; Huang et al., 2000; Yakovleva et al., 2002; Amelchenko, 2006; Kreitschitz, 2012; Boyko, 2013). В большинстве работ рассматривается только поверхность перикарпия, и эти сведения распределяются по секциям рода. Одни авторы считают, что анатомическое строение плодовой и семенной оболочек могут использоваться в качестве таксономических отличий секций, подсекций и отдельных видов (Amelchenko, 2006), другие – что ни один из типов строения поверхности семянок не является характерным для какой-либо отдельной секции (Boyko, 2013). В настоящем исследовании используются представители более мелкой таксономической группировки: подсекции Norvegicae (Rydb.) Korobkov секции Abrotanum (Korobkov, 1981). До настоящего времени нет единого мнения об объеме подсекции в целом и о ранге и систематическом положении некоторых таксонов в подсекции. И.М. Крашенинников (Krasheninnikov, 1946), рассматривал комплекс видов подсекции Norvegicae как отдельный цикл “Polyanthae”, состоящий из двух серий, а именно: серии “Subarctica”, в которую он включал A. norvegica, A. subarctica, A. disjuncta и A. punctigera, и серии “Pacificae”, состоящей из A. arctica. Э. Хультен (Hulten, 1954) считал, что комплекс A. norvegica Fries – A. arctica Less. очень слабо дифференцирован на виды и таксоны внутривидового ранга. Строение сложнорассеченных листьев, характерное для некоторых таксонов этого комплекса, варьирует в удивительно широких пределах и встречается в различных таксономических группах. Созревшие семена, которые могли бы обладать хорошими характеристиками, редко встречаются в материале. В связи с этим задача разделения комплекса на отдельные группы на основе доступного материала далеко не проста. Э. Хультен (Hulten, 1954) предложил для решения таксономических проблем в группе A. norvegica – A. arctica использовать морфологические особенности зрелых семянок наибольшего числа видов, собранных в различных пунктах ареала.
В подсекции Norvegicae насчитывается около 15 таксонов видового и внутривидового рангов, которые представляют морфологически слабо дифференцированную группу травянистых многолетников с обширным раздробленным аркто-альпийским ареалом (Hulten, 1954; Ling, 1992, 1995). В горах Центральной и Южной Норвегии, на севере Шотландии, а также на Полярном и Среднем Урале встречается A. norvegica Fries (Hulten, 1954). Наибольшее габитуальное сходство с A. norvegica имеет A. subarctica Krasch., которая распространена в арктических и горно-таежных районах Якутии (Korobkov, 1981). Специализированным скальным видом подсекции Norvegicae является A. disjuncta Krasch., встречающаяся в высокогорьях Восточного Тянь-Шаня Китая и в Монголии, на песчаных дюнах острова Сахалин произрастает A. punctigera Krasch. ex Poljak. (Krascheninnikov, 1946). В пределах обширного азиатско-американского ареала A. arctica Less. s.l. дифференцирована на географические расы и формы, различающиеся типом опушения или количеством железистых структур. И.М. Крашенинников (Krasheninnikov, 1946) в свое время отмечал, что A. arctica является сборным видом, разбивающимся на несколько мелких видов, каждый со своим обособленным ареалом. К подсекции Norvegicae (серия Pacificae) следует относить также два небольших горных вида, распространенных в Северной Америке в Скалистых горах, а именно: густо опушенную A. saxicola Rydb. и лишенную опушения A. parryi A. Gray. На Аляске на хребте Брукса растет A. comata Rydb. (Rydberg, 1916). В Японии на островах Хоккайдо и Хонсю произрастает в альпийских условиях A. sinanensis Yabe, которая принадлежит также к группе родства Pacificae (Ehrendorfer, 1964). По основным морфологическим признакам все указанные выше таксоны включаются в подсекцию Norvegicae.
В задачи настоящего исследования входило выявление основных признаков строения перикарпия и спермодермы у видов подсекции Norvegicae секции Abrotanum рода Artemisia, собранных в различных пунктах ареала.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для исследования послужили семянки, выделенные А.А. Коробковым из образцов, хранящихся в Гербарии БИН РАН (LE). Всего изучены семянки 7 видов и 4 подвида A. arctica подсекции Norvegicae секции Abrotanum. Для выявления изменчивости морфологических признаков у A. norvegica и A. subarctica и 3 подвидов A. arctica взяты семянки растений, произрастающих в разных точках ареала. Семянки нескольких редких таксонов подсекции Norvegicae локальных популяций из высокогорий Северной Америки и Японии не исследованы.
Artemisia arctica Less. ssp. arctica – 9358-457: Чукотский автономный округ, залив Лаврентия, о. Беннета (Россия), А.А. Коробков, 1969; на восточном побережье, разнотравная луговина на морской террасе.
A. arctica Less. ssp. beringensis (Hultén) Hultén – 9358-528: Берингово море: Накнек, Аляска (США), I.L. Norberg, 1946; на морской террасе.
– 9358-530: Остров Уналяшка (США), Г. Кастельский, 1826–1829; на морской террасе.
A. arctica Less. ssp. ehrendorferi Korobkov – 9358-333: Магаданская обл., Ольский р-н (Россия), В.Н. Васильев, 1938; разнотравный луг на морском берегу.
– П-549: Магаданская обл., окрестности г. Магадана, северный склон Марчеканской сопки (Россия), А.А. Коробков, В.В. Петровский, 1972; на северном склоне, каменистые россыпи на склоне сопки.
– П-546: Западное побережье Охотского моря, бухта Нагаева, окрестности г. Магадана (Россия), А.A. Коробков, В.В. Петровский, 1972; на западном побережье, луговина по берегу ручья.
– 9358-524: Хабаровский край, Верхне-Буреинский р-н, истоки р. Правая Бурея, хребет Дуссе-Алинь (Россия), А.А. Нечаев, 1978; на днище кара в осевой части хребта, 1750 м над ур. м., луговина.
A. arctica Less. ssp. psilosantha Hultén – П-334: Сахалинская обл., о. Парамушир (Россия), Е.С. Короткевич, 1954; на песчаной морской террасе.
– П-334а: Северо-западная оконечность о-ва Кунашир (Россия), В.В. Якубов, 2008; на каменистом склоне и скалах у вершины вулкана.
– 9358-335: Северный Сихотэ-Алинь, долина р. Ботчи, верховья р. Левой Ботчи (Россия), И.К. Шишкин, 1924; на поляне среди кустов кедрового стланика, голец, 1800 м над ур. м.
– 9358-525: Хабаровский край, бассейн р. Тумнин, хребет Большой Ян, гора Командная (Россия), И.Б. Вышин, 1983; на каменистых россыпях подгольцового пояса.
– 9358-531: о. Хоккайдо (Япония), К. Кондо, 1926; в альпийском поясе.
A. comata Rydb. – 9358-519: Plants of the vicinity of Point Lay, Alaska (USA), J.H. Thom, 1950; на краю песчаной речной террасы.
A. disjuncta Krasch. – 2016-03: Баян-Хонгорский аймак, Баян-Гоби сомон, северный макросклон хребта Их-Богдын-Нуру, средняя часть ущелья Их-Пишуг (Монголия), В.И. Дорофеев, А.А. Коробков, Э. Ганболд, Д. Энхтувшин, Б. Гантуяа, 2015; на северном макросклоне хребта в трещинах скал.
A. norvegica Fries. – 9358-523: Plantae Scandinaviae N. Dovre (Norvegieqe), J.H. Hagelin, 1889; на скалах и каменистых россыпях.
– 9358-512: Архангельская обл., восточная часть Большеземельской тундры (Россия), А.М. Верличенко, 1938; в восточной части тундры, каменистый склон хребта.
– 9358-520: Северный Урал. Хребет Уральский. Восточный склон (Россия), Б.А. Тихомиров, 1940; на восточном склоне, сухая кустарничковая пятнистая тундра на склоне хребта.
– 9358-521: Южный Урал. Пермская губерния (Россия), П.Н. Крылов, 1846; в каменистой горной тундре, каменистый склон хребта.
A. punctigera Krasch. ex Poljak. – 9358-337: Сахалинская обл., о. Сахалин, селение Виахту (Россия), П.П. Глен, 1861; на песчаной морской террасе.
A. saxicola Rydb. – 9358-527: шт. Юта (США), E. Neese, 1983; на лугах субальпийской зоны.
A. subarctica Krasch. – 9358-522: Республика (Саха) Якутия, бассейн р. Индигирки, цепь гор Улахан-Эрги-Сис, гора Салыр-Тас (Россия), В.А. Шелудякова, 1936; в горной щебнистой тундре.
– 9358-526: Республика (Саха) Якутия, Томпонский р-н, Верхоянский хребет, западный склон на левом берегу р. Хунхады (Россия), В.П. Самарин, 1954; на западном склоне, лиственничное горное редколесье, 1800 м над ур. м.
– сг-74-67: Якутская АССР, Кондаковские горы, хребет Улахан-Тас, верховья р. Тугучан (Россия), А.А. Коробков, 1974; в каменистой лишайниковой тундре на вершине хребта.
Для исследования с каждого образца брали по 5 семянок и более. Анатомия семянок изучена у всех видов и подвидов. Поверхность семянок изучали в тех случаях, когда позволяло количество материала.
Морфологию и ультраскульптуру поверхности семянок изучали при помощи стереомикроскопа Stemi 2000-C (Carl Zeiss, Germany) и сканирующих электронных микроскопов (СЭМ) JSM 6390 (Jeol, Япония) и MIRA3 (Tescan, Чехия), анатомию – с помощью светового микроскопа (СМ) Axio Lab.A1 (Zeiss, Германия), оснащенного цифровой видеокамерой AxioCam MRc5 с программным обеспечением Zen 2011 и просвечивающего электронного микроскопа (ТЭМ) Libra 120 (Zeiss, Германия).
Для исследования поверхности использовали сухие или размоченные в смеси равных частей воды, глицерина и этилового спирта в течение 5–7 дней семянки. Для СЭМ их фиксировали 2.5% раствором глутарового альдегида на 0.1 М фосфатном буфере pH 7.2–7.4, постфиксировали 1% раствором осмиевой кислоты в течение двух часов при комнатной температуре, обезвоживали в серии спиртов, сушили при критической точке СО2 (CPD300, Leica, Австрия) и напыляли золото (Leica EM SCD500, Leica, Австрия).
Для анатомических исследований семянки фиксировали по общепринятой методике: 3% глутаровым альдегидом и 2% параформальдегидом на 0.1 М фосфатном буфере pH 7.2–7.4 в течение нескольких дней и постфиксировали 2% раствором осмиевой кислоты в течение ночи при +4°С. Материал обезвоживали в серии ацетонов возрастающих концентраций (от 30% до 100%) и заключали в смесь эпона и аралдита.
Для изучения морфологии секрета в секреторных каналах семянки замачивали в спиртовом растворе глицерина на 5–7 дней и затем фиксировали раствором глутарового альдегида и раствором осмиевой кислоты, как указано выше, обезвоживали в серии спиртов и заключили в смолу Spurr.
Полутонкие (толщиной 1–2 мкм) и ультратонкие (толщиной 60 нм) срезы получали с помощью ультрамикротома Ultracut E (Reichert-Jung, Австрия). Полутонкие срезы окрашивали 0.5% раствором толуидинового синего на 1% тетраборате натрия, ультратонкие срезы контрастировали цитратом свинца.
При действии на срезы семянок раствором толуидинового синего происходит окрашивание клеточных стенок в синий, кутикулы в голубой цвета, в результате метахромазии слизистое вещество окрашивается в розовый или фиолетовый цвета (O’Brien et al., 1964; Lillie, 1969). Содержимое секреторных каналов и маслянистый секрет (масло, смола) в клеточных стенках и на поверхности семянок, как правило, не поддаются действию толуидинового синего и сохраняют свой исходный цвет (желтый или коричневый). Специальной окраски на смолы (масло) мы не проводили и определяли наличие секрета в клеточных стенках и полостях клеток экзокарпия, а также на поверхности перикарпия по их цвету.
По возможности в работе проводили совмещение данных, полученных с помощью СЭМ, СМ и ТЭМ.
РЕЗУЛЬТАТЫ
Поверхность перикарпия. С поверхности перикарпий зрелых сухих семянок складчато-бороздчатый (рис. 1A, C), у размоченных семянок поверхность более гладкая (рис. 1B, D). Клетки перикарпия в основном ориентированы вдоль длинной оси семянки (рис. 1E, F). На поверхности отдельных клеток имеется слабо выраженная (рис. 1G) или более крупная ребристость (рис. 1H), ориентированная вдоль (рис. 1G, H) или поперек оси клеток (рис. 1F) (табл. 1). У представителей одного и того же таксона из разных мест произрастания отмечается разное расположение ребристости на поверхности клеток, у некоторых таксонов на разных клетках семянки встречаются одновременно оба варианта расположения ребристости. Например, семянки A. arctica ssp. ehrendorferi из Магаданской области (9358-333) и бухты Нагаева окрестности Магадана (П-546) имели только продольную морщинистость, а из Хабаровского края (9358-524) – и продольную и поперечную морщинистость. У A. norvegica продольная и поперечная морщинистость отмечена у экземпляров из Северного и Южного Урала, а у экземпляров из Норвегии и Архангельской области – только продольная (табл. 1).
Рис. 1.
Общий вид и фрагменты поверхности семянок представителей подсекции Norvegicae.
Общий вид сухих (A, C) и размоченных (B, D) семянок A. norvegica 9358-512 (A, B) и A. subarctica 9358-522 (C, D); фрагменты поверхности перикарпия: клетки ориентированы вдоль длинной оси семянки A. arctica ssp. ehrendorferi П-546 (E) и A. arctica ssp. psilosantha П-334 с ребристостью, ориентированной поперек оси клеток (F); слабо выраженная ребристость A. subarctica 9358-522 (G) и более крупная ребристость A. arctica ssp. psilosantha 9358-525 (H), ориентированные вдоль оси семянок; удлиненные слизесодержащие комплексы A. arctica ssp. psilosantha 9358-525, состоящие из мелких сегментов (I); общий вид карпоподиумов A. subarctica 9358-526 (J) и A. arctica ssp. psilosantha-9358-335 (K). 1 – ребристость, 2 – слизесодержащие комплексы, 3 – карпоподиум.
Масштабные линейки, мкм: B – 500; A, C-E – 200; J, K – 50; I – 20; F–H – 10.
Fig. 1. Achenes and pericarp surface in the subsection Norvegicae members.
Dry (A, C) and soaked in water/alcohol solution (B, D) achenes of A. norvegica 9358-512 (A, B) and A. subarctica 9358-522 (C, D); pericarp surface: cells outstretched along achene axis, A. arctica ssp. ehrendorferi П-546 (E), and cells having ribs perpendicular to cell axis, A. arctica ssp. psilosantha П-334 (F); flattened ribs, A. subarctica 9358-522 (G), and pronounced ribs, A. arctica ssp. psilosantha 9358-525 (H), outstretched along achene axis; elongated mucilage-containing cell complexes composed of small modules, A. arctica ssp. psilosantha 9358-525 (I); carpopodium: A. subarctica 9358-526 (J) and A. arctica ssp. psilosantha 9358-335 (K). 1 – ribbing, 2 – mucilage-containing cell complexes, 3 – carpopodium.
Scale bars, μm: B – 500; A, C-E – 200; J, K – 50; I – 20; F–H – 10.
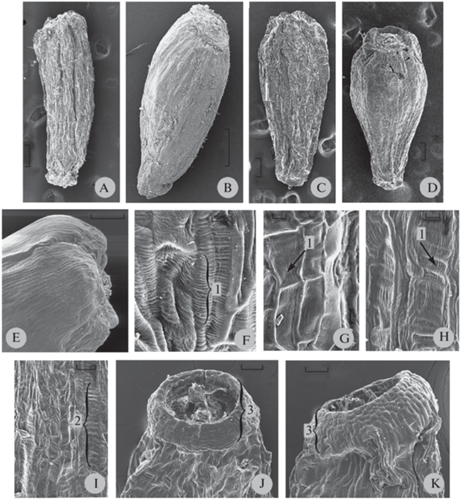
Таблица 1.
Морфологические и анатомические признаки семянок исследованных представителей подсекции Norvegicae секции Abrotanum рода Artemisia (Asteraceae) Table 1. Morphological and anatomical characters of achenes in studied members of the genus Artemisia section Abrotanum subsection Norvegicae (Asteraceae)
| Таксон и его номер
в коллекции LE Taxon and its number in LE collection |
Количество рядов клеток карпоподиума/наличие устьиц на площадке прикрепления венчика Number of cell rows in carpopodium/stomata on corolla attachment site |
Ребристость повер-хности/наличие Сг Ribbing of surface/Sg |
Секреторные структуры Secretory structures |
Количество ребер/Cх
в ребре, цвет секрета Number of ribs/SC per ribs, colour of secret |
Наличие и лока-лизация секрета
в основных клетках экзокарпия/экзотесты Secretion localization in the main cells of exocarp/exotesta |
|---|---|---|---|---|---|
| A. arctica Less. ssp. arctica – 9358-457 | н.и. n.s. |
н.и. n.s. |
Сх +, ск –, нар.кл.ст. утолщена, нар.ч. пектин SC +, mc –, outer cw thickened, outer part pectic |
6/одиночные, голубовато-зеленоватый 6/solitary, bluish-green |
–/– |
| A. arctica Less. ssp. beringensis (Hultén) Hultén – 9358-528 | н.и. n.s. |
Поперечная/н.и. Transversal/n.s. |
Сх недоразвиты, ск –, нар.кл.ст.тонкие SC obsolete, mc –, outer cw thin |
5–6/1–2, | полости клеток/полости клеток cell insides/cell insides |
| A. arctica Less. ssp. beringensis (Hultén) Hultén – 9358-530 | 5–7/+ | Поперечная/н.и. Transversal/n.s. |
Сх недоразвиты, ск –, нар.кл.ст. немного утолщена SC obsolete, mc –, outer cw slightly thickened |
5/одиночные, желтоватый и зеленоватый 5/solitary, yellowish and greenish |
–/клеточные стенки, полости клеток –/cell walls, cell insides |
| A. arctica Less. ssp. ehrendorferi Korobkov – П-549 | н.и. n.s. |
н.и. n.s. |
Сх –, железки +, ск – слизь в полости клетки, нар.кл.ст. ослизняются SC –, glands +, mc contain mucilage inside, outer cw sliming |
4/–, пучки мелкие ассоциированы с железками 4/–, bundles small, beneath the glands |
–/клеточные стенки или полости клеток –/cell walls or cell insides |
| A. arctica Less. ssp. ehrendorferi Korobkov – 9358-333 | 4–5/+ | Продольная/Сг, корот-кие, разной длины Longitudinal/Sg, short, of various length |
Сх +, железки +, ск + SC+, glands +, mc + |
5/одиночные, коричневато-зеленоватый 5/solitary, brownish light green |
клеточные стенки, полости клеток/клеточные стенки cell walls, cell insides/ cell walls |
| A. arctica Less. ssp. ehrendorferi Korobkov – П-546 | 6–8/н.и. 6–8/n.s. |
Продольная/редкие Сг Longitudinal/few Sg |
Сх +, у клеток экзокарпия над Сх масло в кл. ст. и полостях, ск –, нар.кл.ст. ослизняются SC +, exocarp cells next to SC contain oil in cell walls and inside, mc –, outer cw sliming |
5/одиночные, коричневато-зеленоватый 5/solitary, brownish light green |
клеточные стенки, полости клеток/клеточные стенки, полости клеток cell walls, cell insides/ cell walls, cell insides |
| A. arctica Less. ssp. ehrendorferi Korobkov – 9358-524 | 6–8/н.и. 6–8/n.s. |
Продольная, поперечная/Сг, короткие и длинные Longitudinal, transversal/ Sg, short and long |
Сх +, масло в полостях экзокарпия, , железки, ск –, нар.кл.ст. немного утолщена SC +, exocarp cells next to SC contain oil inside, mc –, outer cw slightly thickened |
5/1–2, голубовато-зеленоватый 5/1–2, bluish light green |
полости клеток/клеточные стенки, полости клеток cell insides/cell walls, cell insides |
| A. arctica Less. ssp. psilosantha Hultén – П-334 | 5–6/– | Продольная, поперечная/Сг Longitudinal, transversal/ Sg |
Сх +, масло в кл. ст. неравномерно и полостях экзокарпия, ск –, нар.кл.ст. немного
утолщена SC +, exocarp cells next to SC contain oil inside and irregularly in cell walls, mc –, outer cw slightly thickened |
7/1–2, сероватый 7/1–2, greyish |
клеточные стенки/ клеточные стенки, полости клеток cell walls/cell walls, call cavities |
| A. arctica Less. ssp. psilosantha Hultén – П-334а | 3–5/+ | Продольная, поперечная/Сг Longitudinal, transversal/ Sg |
Сх +, железки, ск + (редкие), нар.кл.ст. ослизняются SC+, mc + (few), glands +, outer cw sliming |
6/одиночные, голубовато-зеленоватый 6/solitary, bluish light green |
клеточные стенки/ клеточные стенки cell walls/cell walls |
| A. arctica Less. ssp. psilosantha Hultén – 9358-335 | 3–4/+ | Продольная, поперечная/Сг Longitudinal, transversal/ Sg, long and short |
Сх +, ск + (редкие), нар.кл.ст. ослизняются SC+, mc + (few), outer cw sliming |
6/1–2, желтый и голубой 6/1–2, yellow and blue |
–/клеточные стенки –/cell walls |
| A. arctica Less. ssp. psilosantha Hultén – 9358-525 | 5–7/– | Продольная, поперечная/Сг, длинные и ко-роткие, со смолой и слизью Longitudinal, transversal/ Sg, long and short, contain resin and mucilage |
Сх +, ск – слизь в полости клетки, нар.кл.ст. ослизняются SC +, mc – contain mucilage inside, outer cw sliming |
6/1–2, голубой 6/1–2, blue |
–/клеточные стенки –/cell walls |
| A. arctica Less. ssp. psilosantha Hultén – 9358-531 | 4–5 (8)/н.и. 4–5 (8)/n.s. |
Продольная, поперечная/Сг Longitudinal, transversal/ Sg |
Сх +, железки, ск + (редкие), нар.кл.ст. ослизняются SC +, glands +, mc + (few), outer cw sliming |
5/1–2, голубовато-желтоватый 5/1–2, bluish yellow |
–/клеточные стенки –/cell walls |
| A. comata Rydb. – 9358-519 | 3–4–5/– | Продольная/Сг, длиной во всю семянку Longitudinal/Sg, as long as achene |
Сх +, ск +, с фибриллами SC +, mc +, contain fibrils |
8/одиночные, желтоватый 8/solitary, yellowish |
–/клеточные стенки, полости клеток –/cell walls, cell insides |
| A. disjuncta Krasch. – 2016-03 | н.и. n.s. |
н.и. n.s. |
Сх +, масло в полости клеток, ск – SC+, oil inside cells, mc – |
6/одиночные, коричневатый 6/solitary, brownish |
–/клеточные стенки, полости клеток –/cell walls, cell insides |
| A. norvegica Fries – 9358-523 | 6–8/н.и. 6–8/n.s. |
Продольная/Сг, длинные и короткие Longitudinal/Sg, long and short |
Сх +, железки, ск +, с фибриллами, нар.кл.ст. ослизняются SC +, glands +, mc +, contain fibrils, outer cw are sliming |
6/1–2, голубовато-коричневатый 6/1–2, bluish light brown |
–/– |
| A. norvegica Fries – 9358-512 | 4–7/+ | Продольная, поперечная/Сг, средней длины, заходят на карпоподиум Longitudinal/Sg, short, entering carpopodium |
Сх +, железки, ск +, с фибриллами SC +, mc +, contain fibrils |
7–10/1–2, коричневатый 7–10/1–2, brownish |
–/– |
| A. norvegica Fries – 9358-520 | 3–7/н.и. 3–7/n.s. |
Продольная, поперечная/Сг, разной длины, заходят на карпоподиум Longitudinal, transversal/ Sg, of different lengths, entering carpopodium |
Сх +, железки, ск +, с фибриллами SC +, mc +, contain fibrils |
7/1–2, коричневато-голубоватый 7/1–2, brownish light blue |
–/клеточные стенки –/cell walls |
| A. norvegica Fries – 9358-521 | 7–11 / н.и. 7–11 / n.s. |
Продольная, поперечная/Сг, разной длины, заходят на карпоподиум Longitudinal, transversal/ Sg, of different length, entering carpopodium |
Сх +, железки, ск +, с фибриллами SC +, mc +, contain fibrils |
8/1–2, коричневатый и голубоватый 8/1–2, brownish and bluish |
–/клеточные стенки –/cell walls |
| A. punctigera Krasch. ex Poljak. – 9358-337 | н.и. n.s. |
н.и. n.s. |
Сх +, железки, ск +, с фибриллами, нар.кл.ст. ослизняются SC+, mc +, contain fibrils, outer cw are sliming |
6–7/1–3, желтоватый 6–7/1–3, yellowish |
клеточные стенки/ клеточные стенки, полости клеток cell walls/cell walls, cell insides |
| A. saxicola Rydb. – 9358-527 | 5–7/н.и. 5–7/n.s. |
продольная / Сг Longitudinal / Sg |
Сх +, железки, ск +, с фибриллами, нар.кл.ст. ослизняются SC +, mc +, contain fibrils, outer cw are sliming |
6/1–2, зеленовато-желтоватый, слизь-розоватый 6/1–2, greenish light yellow, mucilage pinkish |
–/– |
| A. subarctica Krasch. – 9358-522 | 8–10/+ | Продольная, поперечная/– Longitudinal, transver-sal/ – |
Сх +, ск –, нар.кл.ст. ослизняются SC +, mc –, outer cw are sliming |
7/1–4, голубовато-желтоватый 7/1–4, bluish light yellow |
–/клеточные стенки –/cell walls |
| A. subarctica Krasch. – 9358-526 | 4–5/н.и. 4–5/n.s. |
Продольная, поперечная/Сг, длинные и короткие, со смолой Longitudinal, transver-sal/ Sg, long and short, contain resin |
Сх +, ск –, нар.кл.ст. ослизняются SC +, mc –, outer cw are sliming |
6/1–2, голубовато-коричневатый 6/1–2, bluish light browh |
–/клеточные стенки –/cell walls |
| A. subarctica Krasch. – сг-74-67 | 6–7–9/н.и. 6–7–9/n.s. |
Продольная, поперечная/Сг? Longitudinal, transver-sal/ Sg? |
Сх +, ск –, нар.кл.ст. ослизняются SC +, mc –, outer cw are sliming |
6/1–2, желтовато-голубоватый, слизь-розоватый 6/1–2, yellowish light blue, mucilage pinkish |
полости клеток/клеточные стенки, полости клеток cell insides/cell walls, cell insides |
Примечание. Сг – сегментированные клетки, нар.кл.ст. – наружная клеточная стенка перикарпия, нар.ч. – наружная часть клеточной стенки перикарпия, н.и. – не изучено; ск – слизепроизводящие клетки, Сх – секреторный ход, +/– – наличие отсутствие структуры, ? – возможно структура имеется;
Note. Sg – segmented cells; mc – mucilage-producing cells, n.s. – not studied; outer cw – outer cell wall of pericarp cell; outer part – outer part of pericarp cell outer wall; SC – secretory canal, +/– – presence /absence of a structure; ? – probable presence of a structure.
В перикарпии сухих семянок некоторых видов отмечены “комплексы слизесодержащих клеток” (Yakovleva et al., 2002), представляющие собой удлиненные клетки, состоящие из множества мелких сегментов (рис. 1I) (табл. 1). Они находятся на одном уровне с основными более короткими клетками перикарпия.
Карпоподиум у большинства видов кольцеобразный, симметричный (рис. 1J, K) и немного более узкий, чем сама семянка. Карпоподиумы состоят из нескольких рядов выпуклых прямоугольных или слегка вытянутых в продольном или поперечном направлении клеток (табл. 1). Клетки последнего ряда карпоподиума часто удлинены в направлении длинной оси семянки. У одних видов карпоподиум четко выражен, как, например, у A. arctica ssp. ehrendorferi 9358-333 или A. comata, у других – он выражен не четко, например, у A. arctica ssp. psilosantha П-334 и П-334а. В центре площадки карпоподиума у некоторых таксонов (A. subarctica сг-74-67, 9358-526, 9358-522, A. norvegica 9358-520, 9358-521, A. arctica ssp. psilosantha 9358-531, A. arctica ssp. beringensis 9358-530, A. saxicola) находится выступающий проводящий пучок (рис. 2A, B).
Рис. 2.
Общий вид и фрагменты поверхности семянок представителей подсекции Norvegicae.
Карпоподиум A. arctica ssp. psilosantha 9358-531 с выступающим проводящим пучком и клетками перикарпия со сморщенной поверхностью (A); проводящий пучок в карпоподиуме семянки A. subarctica 9358-522 (B); апикальная часть семянки A. arctica ssp. ehrendorferi 9358-333 с площадкой прикрепления венчика (C); фрагмент площадки прикрепления венчика A. arctica ssp. psilosantha 9358-335 с одним рядом устьиц (D); комплексы слизесодержащих клеток на поверхности семянок A. norvegica 9358-520 и 9358-521 (E, F); содержимое слизесодержащих комплексов после размачивания семянок A. norvegica 9358-521 в виде сплошной массы (G) и A. norvegica 9358-512 в виде мелких образований (H); общий вид семянок A. comata со слизесодержащими комплексами после размачивания (I) и A. arctica ssp. psilosantha П-334 с пузырями на поверхности (J); фрагменты перикарпия A. arctica ssp. psilosantha П-334 с пузырями разных размеров (K) и A. arctica ssp. psilosantha 9358-335 с клеточными стенками, имеющих вздутия (L); железки на поверхности перикарпия A. arctica ssp. psilosantha 9358-531 (M), карпоподиум A. norvegica 9358-512 с железками (N); клетки перикарпия Artemisia sp. со сморщенной поверхностью (O). 4 – сморщенная поверхность клетки, 5 – проводящий пучок, 6 – площадка прикрепления венчика, 7 – устьице, 8 – пузырь, 9 – вздутие на поверхности клетки, 10 – железка. Остальные обозначения как на рис. 1.
Масштабные линейки, мкм: F, J – 500; I – 200; C, E, H, N – 100; A, K – 50; D, G, L, M, O – 20; B – 10.
Fig. 2. Achenes and pericarp surface of the subsection Norvegicae members.
Carpopodium with exserted vascular bundle and pericarp cells with wrinkled surface, A. arctica ssp. psilosantha 9358-531 (A); vascular bundle in carpopodium, A. subarctica 9358-522 (B); apical part of the achene with corolla attachment area, A. arctica ssp. ehrendorferi 9358-333 (C); fragment of corolla attachment area with one row of stomata, A. arctica ssp. psilosantha 9358-335 (D); complexes of mucilage-containing cells on the achene surface, A. norvegica 9358-520 and 9358-521 (E, F); contents of mucilage-containing complexes in soaked achenes, in the form of a solid mass, A. norvegica 9358-521 (G), and in the form of small formations, A. norvegica 9358-512 (H); soaked achenes with mucilage-containing complexes, A. comata (I), and with bubbles on the surface, A. arctica ssp. psilosantha П-334 (J); pericarp with bubbles of various size, A. arctica ssp. psilosantha П-334 (K), and with cell walls having bulgings, A. arctica ssp. psilosantha 9358-335 (L); glands on the achene, A. arctica ssp. psilosantha 9358-531 (M), carpopodium with glands, A. norvegica 9358-512 (N); pericarp cells with wrinkled surface, Artemisia sp. (O). 4 – wrinkled surface of the cell, 5 – vascular bundle, 6 – corolla attachment area, 7 – stoma, 8 – bubble, 9 – bulging of the cell surface, 10 – gland. For the other notations, see Fig. 1.
Scale bars, μm: F, J – 500; I – 200; C, E, H, N – 100; A, K – 50; D, G, L, M, O – 20; B – 10.

Апикальная часть семянки имеет неглубокое чашеобразное углубление с закругленными краями, в центре которого располагается диск – площадка прикрепления венчика (рис. 2C). У некоторых видов и географических рас A. arctica на площадке прикрепления венчика имеется кольцо с одним рядом устьиц (рис. 2D) (табл. 1). Паппус у всех изученных представителей подсекции Norvegicae отсутствует.
При размачивании семянок в водно-спиртовом растворе глицерина перикарпий расправляется, делается прозрачным, и становятся видны некоторые детали строения, неразличимые при изучении сухих семян. Так у A. norvegica 9358-523 и A. subarctica 9358-522 при рассматривании с помощью стереомикроскопа в прозрачном перикарпии видны реброподобные уплотнения, тянущиеся вдоль семянки (при сканировании поверхности уплотнения не наблюдаются). В апикальной части семянки уплотнения изгибаются, образуя краевой валик. При исследовании с помощью СЭМ становятся более заметными комплексы слизесодержащих клеток (рис. 2E, F, I). Их содержимое набухает (рис. 2E) и комплексы приподнимаются над поверхностью перикарпия. Содержимое комплексов, разрывает наружную клеточную стенку и становится четче видно сегментированность. Содержимое сегментов в одних случаях представляет собой сплошную массу (рис. 2G), в других – в разрывах клеточных стенок мы видим множество более мелких образований (рис. 2H). Эти комплексы у разных видов различаются по длине: у одних видов имеются только короткие (рис. 2F), у других – длинные и короткие, и только у одного вида (A. comata) они длиной во всю семянку (рис. 2I). Содержимое других клеток, сходных с комплексами слизепроизводящих клеток, т.е. также сегментированных, не разбухает, и эти комплексы остаются плоскими и плохо видны (табл. 1). На поверхности семянок многих видов обнаруживаются пузыри разных размеров (рис. 2J, K). Клеточные стенки отдельных клеток образуют вздутия (рис. 2L) (табл. 2). У некоторых видов на поверхности семянок отмечаются небольшие площадки со сморщенной поверхностью (рис. 2A, O).
Таблица 2.
Сопоставление особенностей строения поверхности семянок и условий мест произрастания представителей подсекции Norvegicae с различными типами секреторных структур и секрета. Table 2. Comparison of structural features of the achene surface and growing conditions of subsection Norvegicae representatives having different secretory structures and secretion
| Таксон и его номер
в коллекции LE Taxon name and its number in LE collection |
Характеристика условий произ-растания Growing conditions |
Наружная клеточная стенка экзокарпия: утолщение/ дополнительный слой на внутренней
поверхности Outer cell wall of exocarp: thickening/exessove layer on the inner surface |
Изменение наружной клеточной стенки экзокарпия при контакте с водой Changing mod of outer exocarp cell wall in the contact with water |
Наличие комплексов слизесодержащих клеток/ локализация слизи Presence of mucilage-containing cell complexes/ localization of mucilage |
Выход секрета на поверхность семянки (количество, форма, локализация) Rise of secretion to achene surface (volume,. form and localization) |
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 |
| Таксоны с секреторными хо-дами и слизевыми клетками Taxa with secretory canals and mucilage cells |
Сухие места обитания Dry habitats |
||||
| A. arctica Less. ssp. ehrendorferi Korobkov П-549 | Ветер, холодно Wind, cold | Утолщена немного/– Slightly thickened/– |
1 | – | Незначительно In small extent |
| A. arctica Less. ssp. psilosantha Hultén 9358-525 | Ветер, холодно Wind, cold | Утолщена/+ Thickened/+ |
1 | Длинные и короткие/ слизь в вакуолях клеток комплекса Long and short/mucilage is in vacuoles of cells in mucilage-containing cell complex |
Пузыри, незначительно, масло на поверхности Bubbles, in small ex-tent, oil on the surface |
| A. arctica Less. ssp. psilosantha Hultén П-334а | Ветер, холодно Wind, cold | Утолщена/+, тонкий Thickened/+, thin |
2 | + (редко) + (rare) |
Bubbles |
| A. arctica Less. ssp. psilosantha Hultén 9358-335 | Ветер, холодно Wind, cold | Утолщена/+ Thickened/+ |
2 | + (редко) + (rare) |
n.s. |
| A. subarctica Krasch. сг-74-67 | Ветер, холодно Wind, cold | Утолщена немного/+, тонкий Slightly thickened/+, thin |
2 | –/Слизь в секреторном ходе –/mucilage within secretory canal | Bubbles |
| A. subarctica Krasch. 9358-526 | Холодно Cold |
Утолщена/+ Thickened/+ |
1 | + | Незначительно In small extent |
| A. subarctica Krasch. 9358-522 | Ветер, холодно Wind, cold | Утолщена/+ Thickened/+ |
1 | – | n.s. |
| A. norvegica Fries. 9358-523 | Ветер, холодно Wind, cold | Утолщена немного/+, тонкий Slightly thickened/+, thin |
1 | + | Bubbles |
| A. norvegica Fries. 9358-512 | Ветер, холодно Wind, cold | Утолщена немного/+, тонкий Slightly thickened/+, thin |
3 | + | n.s. |
| A. norvegica Fries. 9358-520 | Ветер, холодно Wind, cold | Утолщена немного/– Slightly thickened/– |
3 | + | Незначительно In small extent |
| A. norvegica Fries. 9358-521 | Ветер, холодно Wind, cold | Утолщена немного/– Slightly thickened/– |
3 | + | Незначительно In small extent |
| A. punctigera Krasch. ex Poljak. 9358-337 | Холодно Cold |
Утолщена немного/– Slightly thickened/– |
2 | +, короткие +, Short |
Незначительно, мас-ло на поверхности In small extent. oil on the surface |
| A. comata Rydb. 9358-519 | Ветер, холодно Wind, cold | Утолщена немного/– Slightly thickened/– |
3 | + | Пузыри, незначительно Bubbles, in small extent |
| Влажные места обитания Wet habitats |
|||||
| A. arctica Less. ssp. ehrendorferi Korobkov 9358-333 | Холодно Cold |
Утолщена немного/– Slightly thickened/– |
3 | + | Пузыри и вздутия клеточных. стенок Bubbles и bulging of the cell surface |
| A. arctica Less. ssp. ehrendorferi Korobkov П-546 | Холодно Cold |
Утолщена немного/+, тонкий Slightly thickened/+, thin |
1 | + (редко) + (rare) |
0 |
| A. saxicola Rydb. 9358-527 | Холодно Cold |
Утолщена/+ Thickened/+ |
1 | +/Слизь в секреторном ходе +/mucilage within secretory canal |
0, незначительно 0, in small extent |
| A. arctica Less. ssp. arctica 9358-457 | Ветер, холодно Wind, cold | Утолщена/+ Thickened/+ |
1 | Да сильно Abundant |
|
| A. arctica Less. ssp. psilosantha Hultén 9358-531 | Ветер, холодно Wind, cold | Утолщена немного/+, тонкий Slightly thickened/+, thin |
1 | + (редко) + (rare) |
n.s. |
| Таксоны только с секреторными ходами Taxa with secretory canals only |
|||||
| A. disjuncta Krasch. 2016-03 | Ветер, холодно, сухо Wind, cold, dry |
Утолщена немного/+ Slightly thickened/+ |
3 | – | Незначительно In small extent |
| A. arctica Less. ssp. psilosantha Hultén П-334 | Ветер, холодно, сухо Wind, cold, dry |
Утолщена немного/– Slightly thickened/– |
3 | – | Пузыри, незначительно, масло на поверхнисти Bubbles, in small ex-tent, oil on the surface |
| A. arctica Less. ssp. beringensis (Hultén) Hultén. 9358-528 | Ветер, холодно, влажно Wind, cold, wet |
Утолщена немного/– Slightly thickened/– |
3 | – | n.s. |
| A. arctica Less. ssp. beringensis (Hultén) Hultén 9358-530 | Ветер, холодно, влажно Wind, cold, wet |
Утолщена немного/– Slightly thickened/– |
3 | – | Масло на поверхности Oil on the surface |
| A. arctica Less. ssp. ehrendorferi Korobkov 9358-524 | Холодно, влажно Cold, wet |
Утолщена немного/– Slightly thickened/– |
3 | – | n.s. |
Примечания. +/– – наличие/отсутствие сруктуры; n.s. – не исследовано; в столбце 4: 1 – наружная часть наружной клеточной стенки экзокарпия ослизняется, 2 – целиком разбухающая наружная клеточная стенка, 3 – изменения клеточной стенки отсутствуют; в столбце 6 – 0 отмечены таксоны со слабо развитыми секреторными каналами.
Notes. +/–, presence/absence of a structure; n.s., not studied. In column 4: 1, sliming of outer part of exocarp cell wall; 2, outer part of exocarp cell wall swells out; 3, no changes. In column 6: 0, marks taxa with underdeveloped secretory canals.
На поверхности семянок у ряда видов имеются железки, чаще двухрядные (рис. 2M). Железки, как правило, немногочисленные, распределены по всей поверхности семянки. Их много только у A. arctica ssp. ehrendorferi П-549. У A. arctica ssp. psilosantha 9358-531 железки сосредоточены в области прикрепления венчика, а у экземпляров A. norvegica из разных мест произрастания железки располагаются на верхушке семянки или на карпоподиуме (рис. 2N) (табл. 1). Волосков на перикарпии мы не встретили ни у одного таксона.
ПОПЕРЕЧНЫЕ И ПРОДОЛЬНЫЕ СРЕЗЫ
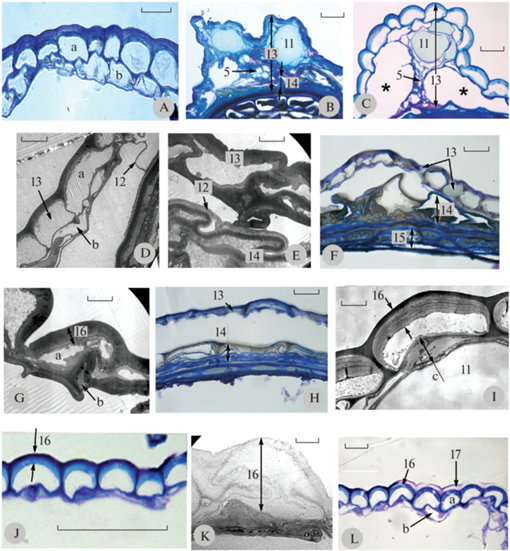
На поперечных срезах видно, что в экзокарпии представителей подсекции Norvegicae встречается как минимум 2 типа клеток: основные клетки эпидермы и клетки, производящие слизь, с разной структурой наружных клеточных стенок. Перикарпий у изученных видов в основном состоит из двух слоев клеток (рис. 3A). У отдельных видов отмечаются участки с 3 слоями клеток (например, у A. punctigera). В области ребер перикарпий многослойный (рис. 3B, C) и, как правило, плотно прилегает к спермодерме. У нескольких представителей (A. arctica ssp. arctica, A. arctica ssp. ehrendorferi П-549, A. arctica ssp. psilosantha 9358-531, A. disjuncta) с боков ребер находятся крупные полости (рис. 3C). С помощью ТЭМ в этих местах обнаруживается тонкая клеточная стенка, тянущаяся от перикарпия к спермодерме, и простирающаяся вдоль наружных клеточных стенок экзотесты (рис. 3D, E) (вероятно, это тонкостенные сильно вытянутые клетки, остатки молодого перикарпия). Клетки перикарпия разных видов бывают в разной степени облитерированы. В большей степени облитерируются клетки второго внутреннего слоя (рис. 3D, F, G, H).
Рис. 3.
Фрагменты поперечных срезов семянок представителей подсекции Norvegicae.
Двухслойный перикарпий A. arctica ssp. ehrendorferi П-546 (A); многослойные участки перикарпия A. subarctica 9358-526 (B) и A. arctica ssp. arctica (C) в районе ребер; клеточная стенка, соединяющая перикарпий и тесту у A. disjuncta (D) и A. arctica ssp. ehrendorferi П-549 (E); фрагменты перикарпия с облитерированными клетками второго слоя A. arctica ssp. psilosantha П-334 (F) и A. arctica ssp. ehrendorferi П-549 (G); фрагменты перикарпия A. arctica ssp. ehrendorferi 9358-333 с облитерированными клетками первого и второго слоя (H); многослойная наружная клеточная стенка перикарпия A. arctica ssp. ehrendorferi П-549 (I); фрагмент перикарпия A. saxicola с многослойной наружной клеточной стенкой (J); электронно-прозрачная наружная клеточная стенка перикарпия A. arctica ssp. psilosantha 9358-335 (K); фрагмент перикарпия A. saxicola с разбухающей наружной частью наружной клеточной стенки (L). 11 – секреторный ход, 12 – клеточная стенка, соединяющая перикарпий и тесту, 13 – перикарпий, 14 – теста, 15 – эндосперм, 16 – клеточная стенка, 17 – разбухающая (ослизняющаяся) наружная часть клеточной стенки, a – первый ряд клеток, b – второй ряд клеток, c – эпителиальные клетки, * – полость. Остальные обозначения как на рис. 1.
Масштабные линейки, мкм: J – 50; A-C, F, H, L – 20; D – 10; E, G, I, K – 5.
Fig. 3. Transverse sections of the achenes in subsection Norvegicae.
Transverse section of double-layered pericarp, A. arctica ssp. ehrendorferi П-546 (A); multilayered parts of pericarps of A. subarctica 9358-526 (B) and A. arctica ssp. arctica (C) in the region of ribs; cell wall connecting the pericarp and the testa in A. disjuncta (D) and A. arctica ssp. ehrendorferi П-549 (E); obliterated cells in the second layer of pericarp, A. arctica ssp. psilosantha П-334 (F), and A. arctica ssp. ehrendorferi П-549 (G); fragment of pericarp with obliterated cells in the first and second layers, A. arctica ssp. ehrendorferi 9358-333 (H); multi-layered outer cell wall of pericarp, A. arctica ssp. ehrendorferi П-549 (I); fragment of pericarp with multi-layered outer cell wall, A. saxicola (J); electronically translucent outer cell wall of pericarp, Artemisia ssp. psilosantha 9358-335 (K); fragment of pericarp with sliming outer part of outer cell wall, A. saxicola (L). 11 – secretory canal, 12 – cell wall connecting the pericarp and the testa, 13 – pericarp, 14 – testa, 15 – endosperm, 16 – cell wall, 17 – sliming outer part of the cell wall, a – the first cell layer, b – the second cell layer, c – epithelial cells, * – cavity. For the other notations, see Fig. 1.
Scale bars, μm: J – 50; A-C, F, H, L – 20; D – 10; E, G, I, K – 5.
Клеточные стенки перикарпия. Основные клетки экзокарпия слизи не содержат, а их наружная клеточная стенка в разной степени утолщена, плотная (рис. 3G, I), иногда состоит из нескольких слоев (рис. 3I, J). У ряда видов имеются клетки с сильно разбухающими наружными клеточными стенками. Эти клеточные стенки слабее окрашиваются толуидиновым синим по сравнению с клеточными стенками основных клеток эпидермы, а в ТЭМ они выглядят электронно-прозрачными (рис. 3K). Очевидно, стенки состоят практически только из углеводов и не содержат белков или фенольных соединений, которые становятся электронно-плотными при подготовке материала для ТЭМ. Разбухающие клеточные стенки отмечаются и у клеток слизепроизводящих комплексов. У изученных представителей имеются клетки, у которых разбухает только наружная часть наружной клеточной стенки перикарпия (рис. 3L). В этом случае разбухшая масса окрашивается толуидиновым синим в фиолетовый цвет, что свидетельствует о наличии пектинов (O’Brien et al., 1964; Lillie, 1969). Такие клеточные стенки отмечаются у подавляющего числа видов (табл. 2). У ряда видов наружные клеточные стенки изнутри имеют дополнительный, более светлый, слой (вероятно, вторичное утолщение). Этот слой толуидиновым синим окрашивается в голубой цвет и бывает утолщенным или очень тонким. На радиальные и внутренние тангентальные клеточные стенки дополнительный слой практически не распространяется. Замечено, что у таксонов, произрастающих на лугу или морской террасе, дополнительный слой у клеточных стенок не образуется (табл. 2).
Слизесодержащие структуры. В перикарпии они, как правило, находятся в экзокарпии (рис. 4A–D) и имеются не у всех представителей подсекции Norvegicae (табл. 1). У ряда видов слизь, вероятно, имеется и во втором слое перикарпия, клетки которого большей частью облитерированы (A. norvegica, A. saxicola). Иногда внутренние стенки клеток второго слоя перикарпия разрываются, и слизь заполняет пространство между перикарпием и спермодермой. У A. arctica ssp. arctica слизесодержащих клеток в перикарпии не встречено, но слизистый слой отмечен в тесте (рис. 4E). В тесте слизистый слой, вероятно, имеется и у A. norvegica 9358-520, A. arctica ssp. ehrendorferi 9358-524, A. subarctica 9358-522.
Рис. 4.
Фрагменты поперечных и продольных срезов семянок представителей подсекции Norvegicae.
Слизесодержащие комплексы: в экзокарпии A. norvegica 9358-520 (A), однорядные и многорядные A. norvegica 9358-521 (B), многоярусные и многорядно-многоярусные A. norvegica 9358-512 (C, D); клетка со слизью в тесте A. norvegica 9358-520 (E); продольный срез слизесодержащего комплекса в перикарпии A. norvegica 9358-521 (F); клетки перикарпия A. arctica ssp. psilosantha 9358-525 со слизью в вакуолях (G); фрагмент перикарпия A. saxicola с секреторным ходом и слизесодержащей клеткой (H); поперечные срезы семянок: A. norvegica 9358-521 с равномерным расположением секреторных ходов (I), A. norvegica 9358-523 с секреторными ходами на противоположных сторонах семянки (J), A. subarctica 9358-522 с односторонним расположением секреторных ходов (K); фрагменты поперечных срезов перикарпия: ребро A. disjuncta с одиночным секреторным ходом (L); ребро A. norvegica 9358-523 с секреторным ходом и слизесодержащими клетками в экзокарпии (M). 18 – однорядные слизесодержащие комплексы, 19 – многорядные слизесодержащие комплексы, 20 – многоярусные слизесодержащие комплексы, 21 – слизесодержащая клетка, 22 – слизь, d – фибриллярное вещество. Остальные обозначения как на рис. 1, 2, 3.
Масштабные линейки, мкм: I – 200; J, K – 100; E, H, L – 50; A-F – 20; M – 2.5; G – 2.
Fig. 4. Transverse and longitudinal sections of achenes in the subsection Norvegicae members.
Mucilage-containing complexes in exocarp, A. norvegica 9358-520 (A), one- and multi-row ones, A. norvegica 9358-521 (B), multi-tiered and multi-row multi-tiered, A. norvegica 9358-512 (C, D); cell of testa containing mucilage, A. norvegica 9358-520 (E); longitudinal section of mucilage-containing complex in pericarp, A. norvegica 9358-521 (F); pericarp cells containing mucilage in vacuoles, A. arctica ssp. psilosantha 9358-525 (G); fragment of pericarp with secretory canal and mucilage-containing cell, A. saxicola (H); transverse sections of achenes: with evenly spaced secretory canals, A. norvegica 9358-521 (I), with secretory canals at the opposite sides of the achene, A. norvegica 9358-523 (J), with secretory canals on one side of the achene, A. subarctica 9358-522 (K); transverse sections of pericarp: rib with solitary secretory canal, A. disjuncta (L); rib with secretory canal and mucilage-containing cells in exocarp, A. norvegica 9358-523 (M). 18 – one-row mucilage-containing complexes, 19 – multi-row mucilage-containing complexes, 20 – multi-tiered mucilage-containing complexes, 21 – mucilage-containing cell, 22 – mucilage, d – fibrillar substance. For the other notations, see Figs. 1–3.
Scale bars, μm: I – 200; J, K – 100; E, H, L – 50; A-F – 20; M – 2.5; G – 2.

На поперечных срезах слизесодержащие структуры в разной степени возвышаются над основными, не содержащими слизи, клетками и представляют собой слизепроизводящие комплексы (рис. 2E–I). Между собой сегменты, слагающие слизевой комплекс, разделены тонкими клеточными стенками (рис. 4F). У представителей подсекции Norvegicae встречаются слизепроизводящие комплексы, сегменты которых содержат только слизь (рис. 4A, B) и сегменты с фибриллярным веществом, погруженным в слизь (рис. 4B–D, F). У изученных представителей подсекции Norvegicae на поперечных срезах видно, что в слизевых комплексах может образовываться несколько рядов слизи, разделенных тончайшими клеточными стенками. В одних случаях на поперечных срезах комплекс состоит из 3–5 узких сегментов (рис. 4B), т. е. можно сказать, что они многорядные. В этом случае они практически не отличаются от продольных срезов комплексов слизепроизводящих клеток (рис. 4F), они различаются только количеством входящих в их состав сегментов. В других случаях ряды слизи с содержащимся в них фибриллярным веществом располагаются друг над другом (рис. 4C, D) и так же разделены клеточными стенками. Это уже не многорядные, а многоярусные слизепроизводящие комплексы. Встречаются и комбинированные слизесодержащие комплексы: многорядно-многоярусные (рис. 4D). В то же время, у отдельных представителей подсекции отмечаются клетки, у которых слизь, вероятно, отложена не в периплазмтическое пространство, а в вакуоль (рис. 4G) (A. arctica ssp. psilosantha 9358-525, A. arctica ssp. ehrendorferi П-549). При намачивании эти клетки не разбухают и не увеличиваются в размерах.
Слизь также встречается в отдельных секреторных ходах у A. saxicola и A. subarctica сг-74-67. У A. saxicola в секреторном ходе находятся слизь и капли масла, а в экзокарпии этому ходу соответствует довольно крупная слизесодержащая клетка (рис. 4H).
На продольных срезах A. arctica ssp. psilosantha 9358-525 и A. subarctica 9358-526 встречаются комплексы клеток, объединенные единой клеточной стенкой, но заполненных не слизью, а маслянистым секретом.
Секреторные структуры перикарпия. Почти у всех изученных видов имеются секреторные каналы (табл. 1), только у одного образца A. arctica ssp. ehrendorferi П-549 секреторные каналы отсутствовали, а на поверхности перикарпия имелись многочисленные железки (табл. 1).
Секреторные каналы у изученных образцов находятся в ребрах под клетками экзокарпия. Клетки экзокарпия, находящиеся над каналами, облитерированы (рис. 3B; рис. 4M) или не облитерированы (рис. 3C; рис. 4H, L). У A. norvegica и A. comata над каналами имеются слизесодержащие комплексы (рис. 4J, M). Виды различаются по количеству и распределению ребер с секреторными каналами у семянок (табл. 1). Ребра равномерно (A. norvegica 9358-512, 9358-520, A. arctica, A. disjuncta) или неравномерно (A. norvegica 9358-523, A. subarctica 9358-522) распределены по периферии семянки (рис. 4I-K). На поперечных срезах видно, что перикарпий в области ребер плотно прилегает к семенной кожуре (рис. 3B, C; рис. 4L), в межреберных участках у большинства видов перикарпий примыкает к семенной кожуре не плотно (рис. 3C, F, H). Клетки перикарпия, входящие в состав ребра, немногочисленные, тонкостенные и, как правило, облитерированы. Секреторные каналы занимают большую часть поперечного среза (или объема) ребра (рис. 3B, C; рис. 4H–L). С поверхности перикарпия и на продольных срезах видно, что вещество, находящееся в секреторных каналах, распределено по их длине неравномерно. Вероятно, в связи с этим не во всех каналах на поперечных срезах мы обнаруживали содержимое. Каналы в ребрах одиночные (рис. 3C; рис. 4H-J, M) или собраны по два–три (рис. 3B; рис. 4K) (табл. 1). У некоторых видов на месте секреторных каналов отмечаются группы клеток, заполненных секретом, а полость отсутствует (A. arctica ssp. ehrendorferi, A. punctigera, A. subarctica). Ходы изнутри выстланы уплощенными секреторными (эпителиальными) клетками (рис. 3I). Содержимое, находящееся в эпителиальных клетках секреторного канала, выделяется в полость канала через клеточные стенки в результате их разрыхления.
Секрет в каналах бывает однородным (рис. 3C) (A. disjuncta, A. punctigera, A. arctica 9358-457, 9358-524, П-546, 9358-333, 9358-531, 9358-525; A. norvegica 9358-523, 9358-521; A. subarctica 9358-522, 9358-526) и/или состоящим из пластинок (A. arctica 9358-333, П-334; A. subarctica сг-74-67; A. norvegica 9358-512, 9358-523), или пористым (рис. 5A) (A. norvegica 9358-520, 9358-521; A. saxicola). Цвет секрета у видов также различается: он может быть желтым или коричневым, или при окрашивании толуидиновым синим полутонких срезов становится голубоватым, или зеленоватым (табл. 1), что, вероятно, свидетельствует об их разном химическом составе. Различается цвет секрета и у представителей одного и того же таксона, собранных в разных точках ареала. Секрет содержится не только в полости, но и в эпителиальных клетках канала, а также паренхимных клетках перикарпия, окружающих секреторный канал.
Рис. 5.
Поперечные срезы семянок представителей подсекции Norvegicae.
Фрагмент поперечного среза семянки A. norvegica 9358-520 с пористым секретом в канале (A); проводящие пучки перикарпия: A. arctica ssp. ehrendorferi П-549 с двумя трахеальными элементами (B); A. disjuncta с сильно утолщенным вторичным утолщением трахеального элемента (C); A. norvegica 9358-521 (D) и A. comata (E) (пучок не связан с секреторным ходом) с механическими клетками; кутикула на поверхности семенной кожуры A. arctica ssp. psilosantha 9358-335 (F); многокомпонентные проводящие пучки семенной кожуры A. subarctica 9358-526 (G) и A. norvegica 9358-521 (H); фрагменты семенной кожуры и эндосперма A. arctica ssp. ehrendorferi 9358-524 (I) и A. arctica ssp. psilosantha П-334 (J); фрагмент перикарпия Dendranthema naktogense с ослизняющейся наружной частью утолщенной наружной клеточной стенки (K). 23 – трахеальный элемент проводящего пучка, 24 – механическая клетка, 25 – кутикула. Остальные обозначения как на рис. 1, 2, 3.
Масштабные линейки, мкм: A, K – 50; E, G–J – 20; B–D, F – 5.
Fig. 5. Transverse sections of achenes in the subsection Norvegicae members.
Part of achene transverse section with porous secretion within the canal, A. norvegica 9358-520 (A); vascular bundles of pericarp: containing two tracheary elements, A. arctica ssp. ehrendorferi П-549 (B), massive secondary thickening of the tracheary element, A. disjuncta (C), containing mechanical cells, A. norvegica 9358-521 (D) and A. comata (E) (vascular bund not related to secretory canal); cuticle on the surface of the seedcoat, A. arctica ssp. psilosantha 9358-335 (F); multicomponent vascular bundles of seedcoat, A. subarctica 9358-526 (G) and A. norvegica 9358-521 (H); seedcoat and endosperm, A. arctica ssp. ehrendorferi 9358-524 (I) and A. arctica ssp. psilosantha П-334 (J); swelling outer part of thickened outer cell wall in pericarp, Dendranthema naktogense (K). 23 – tracheary element of vascular bundle, 24 – mechanical cell, 25 – cuticle. For the other notations, see Figs. 1–3.
Scale bars, μm: A, K – 50; E, G–J – 20; B–D, F – 5.

Помимо секрета в секреторных каналах и окружающих клетках, в перикарпии отмечено накопление маслоподобного содержимого в клеточных стенках и полости клеток экзокарпия. В тесте оно обнаружено в экзотесте: в полости клеток и/или в наружных клеточных стенках (не включая кутикулы) (табл. 1). Содержимое полостей клеток экзокарпия и экзотесты и клеточных стенок не всегда совпадает по цвету с секретом секреторных каналов (табл. 1).
Каналы всегда соседствуют с проводящими пучками. Количество каналов (или групп каналов, если они собраны по 2–3) равно количеству проводящих пучков в перикарпии.
Проводящие пучки и механические клетки. Число проводящих пучков различно у представителей разных таксонов (у A. arctica ssp. arctica, A. arctica ssp. psilosantha, A. disjuncta, A. punctigera, A. saxicola, A. subarctica (кроме образца, собранного в бассейне р. Индигирки) оно равно 6, у A. arctica ssp. ehrendorferi и A. arctica ssp. beringensis – 5, у A. comata – 8, и колеблется у A. norvegica из разных мест произрастания от 6 до 8). Проводящие пучки располагаются среди паренхимных клеток в ребрах непосредственно под одиночными секреторными каналами или между сдвоенными каналами (рис. 3B, C; 4M; 5A) и содержат у разных видов от 1 до 8 трахеальных элементов (рис. 5B, C). В единичных случаях встречаются проводящие пучки, не связанные с секреторными каналами (A. norvegica 9358-523 и A. comata). У трахеальных элементов отмечается сильное развитие вторичных утолщений (рис. 5B, C). Механическая ткань выражена слабо. Отдельные ее элементы сосредоточены в проводящих пучках (рис. 5D). У A. comata и A. subarctica сг-74-67 отмечены крупные относительно тонкостенные механические клетки. Только у A. comata они располагаются с двух сторон от проводящего пучка, не связанного с секреторными ходами (рис. 5E), а у A. subarctica сг-74-67 они рядом с секреторными образованиями (ходами и группами клеток, заполненных секретом). Следует отметить, что на поперечных срезах довольно трудно отличить трахеальные элементы от механических клеток, так как форма и толщина клеточных оболочек механических клеток и трахеальных элементов сходны.
Семенная кожура. Спермодерма имеется у всех исследованных представителей подсекции Norvegicae. Клетки экзотесты в разной степени облитерированы, внутренние клетки спермодермы облитерированы полностью (рис. 3F, H; 4L; 5A, F, H, I, J). У двух видов на поверхности клеток экзотесты обнаруживается заметной толщины кутикула (рис. 5F) (A. arctica ssp. psilosantha 9358-335, 9358-525, A. norvegica 9358-523). Относительно часто на поперечных срезах встречается проводящий пучок, состоящий из трахеальных элементов числом от 4–5 до 17–20. В этом месте спермодерма состоит из большего числа клеток (рис. 5G, H). В клеточных стенках и полостях клеток экзотесты практически у всех видов и их разновидностей обнаруживается коричневатое вещество (табл. 2), которое, очевидно, определяет цвет семянок (при намачивании семянок видно, что клетки перикарпия прозрачные). Отмеченный нами цвет семенной кожуры совпадает с цветом семянок, указанным Коробковым в разделе “Систематический конспект рода Artemisia …” (Korobkov, 1981). Э.В. Бойко (Boyko, 2013) также отмечает, что цвет семянок определяется окраской эпидермы семени, а плодовая оболочка прозрачная.
Эндосперм плотно примыкает к спермодерме, между ними всегда имеется кутикула, сформированная клетками эндосперма. У большинства видов он состоит из двух слоев клеток, в некоторых случаях отмечается три слоя (в частности, у A. norvegica 9358-512). Второй слой клеток, как правило, облитерирован (рис. 3F, 4A; 5A, F, I, J).
ОБСУЖДЕНИЕ
В результате проведенного исследования мы получили новые данные о морфологии и анатомическом строении семянок представителей подсекции Norvegicae секции Abrotanum.
На площадке прикрепления венчика у A. arctica ssp. beringensis, A. arctica ssp. ehrendorferi, A. arctica ssp. psilosantha, A. norvegica, A. subarctica обнаружены устьица, которые, скорее всего, относятся к нектарному кольцу, окружающему основание столбика – стилоподиум. У представителей Artemisia наличие нектарников в известной нам литературе не отмечалось. Однако местоположение нектарников и устьиц с ними связанными известно у представителей сложноцветных (Alexandrov, Savchenko, 1951) и, особенно у видов Anthemideae, где площадка прикрепления венчика может называться нектарником или стилоподиумом (Oberprieler et al., 2009).
Показаны два способа выделения секрета из каналов. Выше мы отмечали, что у ряда видов на поверхности перикарпия встречаются пузыри (табл. 2) (рис. 2J, K). Это сгустки смолы, вышедшей на поверхность путем диффузии, как у других представителей Anthemideae (Savchenko, 1949). Известно, что секрет может продвигаться через клеточные стенки без их изменения (Kolalite, 2001; Kolalite et al., 2003). У изученных представителей подсекции Norvegicae отмечается и другой способ выделения секрета: в результате разрыхления внутренних стенок эпителиальных клеток секреторных каналов. Подобное выведение секрета характерно и описано у растений, выделяющих эфирные масла (Muravnik, 2007). Известно, что в секрете секреторных ходов семянок подсекции Norvegicae содержится эфирное масло, которым богаты представители трибы Anthemideae (Denisova, 1989).
Определены структуры, разбухающие при намокании семени (рис. 3K). На поверхности перикарпия сухих семянок с помощью СЭМ выявлены клетки со сморщенной поверхностью (рис. 2A, O). На поперечных срезах у одиночных клеток экзокарпия и у клеток слизепроизводящих комплексов ряда видов отмечены разбухающие наружные клеточные стенки (табл. 2). Такие клеточные стенки описаны нами ранее у A. subarctica (Yakovleva et al., 2002) и показано, что в них содержится незначительное количество цементирующего их вещества, соединений кремния. Мы полагаем, что сморщенные поверхности сухих семянок соответствуют разбухающим наружным клеточным стенкам, которые при фиксации материала для анатомических исследований разбухают, поглощая влагу из применяемых реагентов.
Ребра у представителей подсекции Norvegicae по своему строению сильно отличаются от ребер, встречающихся у большинства представителей трибы Anthemideae (Alexandrov, Savchenko, 1949, 1951; Melikyan, Muradyan, 1975). Ребра у исследованных представителей подсекции Norvegicae практически не заметны на поверхности перикарпия сухих семянок. У размоченных семянок они проявляются в виде реброподобных тяжей, слегка выступающих над поверхностью. Ребра представителей подсекции Norvegicae состоят из 1–3 секреторных ходов, расположенных под экзокарпием, нескольких более или менее облитерированных тонкостенных паренхимных клеток и слабо развитого проводящего пучка, находящегося под секреторными каналами, в состав которого иногда входят механические клетки. Гидроцитов, толстостенных паренхимных клеток, механических клеток, образующих обкладку проводящего пучка, всех этих элементов, характерных для ребер представителей трибы Anthemideae (Alexandrov, Savchenko, 1949; Melikyan, Muradyan, 1975) мы не встретили ни у одного из представителей подсекции Norvegicae.
Впервые у представителей подсекции Norvegicae в перикарпии обнаружено несколько вариантов слизепроизводящих структур. В первую очередь к ним относятся комплексы слизесодержащих клеток с разрывающимися (A. comata, A. nor-vegica 9358-521) и не разрывающимися (A. arctica ssp. arctica, A. arctica ssp. psilosantha, A. norvegica, A. punctigera, A. subarctica) наружными клеточными стенками. У последних наружные клеточные стенки бывают не разбухающими (A. arctica ssp. arctica, A. norvegica) и разбухающими (A. arctica ssp. psilosantha, A. norvegica, A. punctigera, A. subarctica). Ранее было отмечено (Yakovleva et al., 2002), что разрываются наружные клеточные стенки в результате отсутствия в них цементирующего вещества (например, соединений кремния в аморфной форме). Клеточные стенки, пропитанные соединениями кремния, не разрываются, или разбухают, если кремний в них находится в незначительных количествах. Препятствовать разрыву клеточных стенок может и относительно толстая кутикула (Yakovleva et al., 2002). Среди комплексов слизесодержащих клеток представителей подсекции Norvegicae нами отмечены многорядные и многоярусные. На фотографиях, полученных с помощью СЭМ, у размоченных семянок встречаются комплексы, состоящие из сегментов, содержащих сплошную массу (рис. 2G), и состоящие из множества мелких образований (рис. 2H). Эти же комплексы на поперечных срезах семянок выглядят совершенно по-другому. На рисунке 4B слева находится клетка, описанная как содержащая сплошную массу, а справа – как с множеством мелких образований. Полагаем, что эти клетки правильнее интерпретировать, как однорядную (слева) и пятирядную (справа). Многоярусные слизесодержащие комплексы с помощью СЭМ нам, к сожалению, не удалось увидеть. Вероятно, многорядные и многоярусные слизепроизводящие комплексы образуются и у других представителей рода Artemisia, об этом свидетельствуют рисунки в некоторых работах (например, Yakovleva et al., 2002: рис. 1б; Amelchenko, 2006: рис. 7–10, 14, 17, 20–22, 33, 35 ). У отдельных представителей подсекции Norvegicae отмечаются клетки, у которых слизь отложена не в периплазматическое пространство, а в вакуоль (рис. 4G). Слизь может появляться и в результате ослизнения наружной части наружной клеточной стенки перикарпия (рис. 3L).
Наличие слизесодержащих комплексов с ослизняющимися и неослизняющими наружными клеточными стенками отмечается не только у видов Artemisia (Korobkov, 1973; Yakovleva et al., 2002; Kreitschitz, 2012; Boyko, 2013), но и у многих других видов. Приведем несколько примеров. Разрываются (“ослизняются”) наружные клеточные стенки слизепроизводящих клеток в перикарпии и семенной кожуре у Anastatica hierochuntica L., Anthemis chia L., Arabidopsis thaliana (L.) Heynh., Capsella bursa-pastoris (L.) Medik., Cardaria draba Desv., Carrichtera annua (L.) Prantl, Goodenia ovata Sm., ряда видов Lepidium, Matricaria lamellata Bunge, у видов Plantago, многих видов Stroganowia, Tanacetum umbelliferum Boiss., не разрываются (“не ослизняются”) – у многих видов Stroganowia, Tridactylina kirilovii (Turcz) Sch. Bip., Sisymbriuym subspinescens Bunge, Stubendorffia aptera Lipsky, Urtica dioica L. (Alexandrov, Savchenko, 1949, 1951; Yakovleva, Kravtzova, 1999; Karpova et al., 2010; Karpova, Yakovleva, 2010, 2012; Yakovleva et al., 2010; Kreitschitz, 2009; и др.). Слизь комплексов с разрывающимися (“ослизняющимися”) наружными клеточными стенками образует слизистый чехол на поверхности семянки. A. Kreitschitz (Kreitschitz, Valles, 2007; Kreitschitz, 2009, 2012) так же как и другие авторы (Alexandrov, Savchenko, 1949, 1951; Korobkov, 1981; Huang et al., 2000, 2008; Yang et al., 2011, 2012) считает, что формирование слизистой оболочки является приспособлением для накопления влаги в засушливых условиях произрастания. Относительно “не ослизняющихся” клеток существует мнение, что их появление связано с переходом растений от засушливых мест произрастания в более влажные. Запасенная в “не ослизняющихся” клетках вода может служить для начальных стадий прорастания (Kreitschitz, 2012), или она является необходимой для поддержания покоя или дозревания семян (Karpova, Yakovleva, 2012). К этой же категории, вероятно, можно отнести клетки со слизью в вакуоли (рис. 4G). Слизь этих клеток, возможно, спасает семянки от промерзания, препятствуя образованию льда.
Многоярусные слизесодержащие комплексы сходны со слизепроизводящими клетками листьев. Эти клетки имеют два или более слоя слизи, разделенных клеточной стенкой (Yakovleva, 1988). Мы полагаем, что образование нескольких слоев слизи, разделенных клеточными стенками, связано с ритмами развития растения. У A. norvegica в связи с суровыми условиями произрастания (например, сильное колебание суточных температур), созревание семянок и соответственно секреция слизи происходят в несколько этапов, что и отражается в строении слизепроизводящих клеток. Каждый этап секреции слизи завершается достраиванием клеточной стенки, разделяющей слои слизи (Yakovleva, 1990).
У двух таксонов (A. arctica ssp. psilosantha 9358-525 и A. subarctica 9358-526) в сегментированных клетках, сходных с комплексами слизесодержащих клеток, содержимое имеет другую морфологию. У A. arctica ssp. psilosantha в семянках отмечаются сегментированные клетки, как содержащие слизь, так и содержащие масло. Имеется еще несколько таксонов, у которых на поверхности отмечаются сегментированные клетки, а на поперечных срезах слизь не обнаруживается. К ним относятся – A. arctica ssp. psilosantha П-334, A. subarctica сг-74-67, A. arctica ssp. ehrendorferi П-546 и 9358-524 (табл. 1). Возможно, что и у них вместо слизи в сегментированных клетках находится масло. То, что структура слизесодержащего комплекса на поверхности семянок может сохраняться, но слизеобразование в клетках не наблюдается, отмечает и Э.В. Бойко (Boyko, 2011). В связи с приведенными выше примерами, полагаем, что следует с осторожностью применять термин “комплексы слизесодержащих клеток” при описании поверхности семянок не только представителей рода Artemisia, но и всей трибы Anthemideae. Предлагаем заменить термин “комплексы слизесодержащих клеток” на более нейтральный термин “сегментированные клетки” (табл. 1), и применять этот термин при описании клеток поверхности перикарпия без проверки наличия слизи в клетках на поперечных или продольных срезах. При определении же наличия того или иного вещества, писать “сегментированные слизесодержащие клетки” или “сегментированные маслосодержащие клетки”. Относительно многорядных и многоярусных комплексов слизесодержащих клеток, будет проще сказать: однорядные сегментированные слизесодержащие клетки (рис. 2G, рис. 4A, B) или многорядные (рис. 2H, рис. 4B) и/или многоярусные (рис. 4C, D) сегментированные слизесодержащие клетки.
Относительно исследованной в настоящей статье группы Norvegicae, мы полагаем, что незначительное развитие слизепроизводящих клеток совместно с выступающей на поверхность перикарпия смолой, скорее всего, служат защитой семянки от пересыхания при сильном ветре и сухости условий произрастания (табл. 2), а также для прикрепления к субстрату (табл. 2). При смачивании семянки перед прорастанием содержимое секреторных каналов, имеющихся практически у всех исследованных видов, выходит в районе площадки прикрепления венчика и прикрепляет семянку к субстрату (Яковлева, собственные наблюдения). Для поглощения воды при прорастании у этой группы растений, вероятно, выработался свой механизм: в центре карпоподиума, через который появляется корешок, у них имеется проводящий пучок с сосудами. Возможно, через эти сосуды и происходит поступление воды в семянку, необходимой для прорастания. Ослизняющиеся наружные части наружных клеточных стенок перикарпия также могут участвовать в поддержании определенной влажности семянок, в производстве дополнительного тепла при прорастании семянок, так как считается, что процесс набухания слизи связан с тепловыми изменениями в ней. A. Kreitschitz (Kreitschitz, 2009) сообщает, что во время гидратации пектина и связывания частиц воды, производится определенное количество тепла и это может увеличить температуру и влиять на развитие растений, в том числе и на прорастание (Kreitschitz, 2009).
ОБЩАЯ ХАРАКТЕРИСТИКА СЕМЯНОК ПРЕДСТАВИТЕЛЕЙ ПОДСЕКЦИИ NORVEGICAE
На основании полученных данных была разработана общая характеристика семянок подсекции Norvegicae и выделены признаки, отличающие эту подсекцию от других таксонов рода Artemisia. Постоянным для всех исследованных видов подсекции Norvegicae является: отсутствие волосков на перикарпии, у всех исследованных видов отмечен хорошо выраженный карпоподиум, в центре которого находится выступающий проводящий пучок, состоящий из трахеальных элементов; у основания стилоподиума имеются устьица, вероятно, принадлежащие нектарникам; сегментированные слизесодержащие клетки имеются не у всех представителей секции, они ориентированы вдоль длинной оси семянки, расположены не регулярно и относительно короткие; на поверхности перикарпия кутикула тончайшая или она отсутствует; ребра на поверхности сухих семянок практически не заметны; у размоченных семянок они проявляются в виде реброподобных тяжей, слегка выступающих над поверхностью, и по строению отличаются от ребер других представителей трибы Anthemideae. В ребрах представителей подсекции Norvegicae находятся эфиромасличные ходы в количестве 1–3, проводящие пучки и тонкостенные паренхимные клетки. Гидроциты отсутствуют. Механические клетки в небольшом количестве встречаются в проводящих пучках. Перикарпий между ребрами двухслойный, при этом клетки второго слоя полностью облитерированы. Сегментированные клетки могут быть со слизью и/или маслом. Сегментированные слизесодержащие клетки бывают однорядные и/или многорядные и многоярусные. В семянках отдельных представителей подсекции в слизи сегментированных слизесодержащих клеток обнаруживаются стопки фибрилл, но подковообразные структуры они не образуют, как это было показано для представителей других секций рода Artemisia (Yakovleva et al., 2002). Слизь образуется еще и в результате “ослизнения” наружной части наружной клеточной стенки перикарпия. Спермодерма хорошо выражена, многослойная, экзотестальная. В клеточных стенках и полостях клеток экзотесты содержатся маслянистые вещества. Можно сказать, что у исследованных представителей подсекции Norvegicae большую значимость в жизнеобеспечении семянок имеют секреторные образования (секреторные ходы и сегментированные клетки, содержащие масло), а слизесодержащие сегментированные клетки занимают второстепенное значение в отличие от других представителей рода Artemisia (Boyko, 2013). По числу ребер таксоны подсекции Norvegicae различаются незначительно (табл. 1). У A. norvegica из разных мест произрастания число ребер колеблется от 6 (у норвежской формы) до 8 (у экземпляров с Южного Урала). Обнаружено, что у подвидов A. arctica (ssp. beringensis, ssp. ehrendorferi, ssp. psilosantha), экземпляры которых произрастали во влажных условиях (табл. 2), количество ребер в семянках постоянно и равно 5 (табл. 1). У экземпляров, произраставших в сухих условиях (табл. 2), подвидов A. arctica (ssp. beringensis, ssp. psilosantha), а также у видов A. norvegica и A. subarctica количество ребер больше и колеблется от 6 до 7 (табл. 1). У A. arctica ssp. ehrendorferi, произраставшей в сухих условиях на северном склоне Марчеканской сопки, самое низкое количество ребер (4), этот же образец отличается от остальных экземпляров рядом признаков строения семянки (см. ниже). У A. norvegica с Южного Урала (9358-521) наоборот самое большое количество ребер (8) у семянки в отличие от экземпляров этого же вида, произраставших в сухих условиях. A. norvegica с Южного Урала, так же как и предыдущий образец A. arctica ssp. ehrendorferi, отличается от остальных экземпляров этого же вида рядом признаков (см. ниже). Наши данные совпадают с данными В.Г. Александрова и М.И. Савченко (Alexandrov, Savchenko, 1949), о том, что количество пучков в перикарпии может быть непостоянным у видов одного рода, зависит от условий произрастания и не может быть использовано как систематический признак. Основываясь на цвете секрета и полагая, что цвет секрета определяется его химическим составом, можно сказать, что состав секрета у представителей разных видов подсекции Norvegicae не одинаков. Окраской секрета могут различаться представители одного вида и подвида из разных мест произрастания, в частности, A. arctica ssp. ehrendorferi, A. arctica ssp. psilosantha, A. norvegica, A. subarctica. То, что химический состав секрета в разных местах обитания может меняться, отмечается для вида A. annua L. (Zhigzhitzhapova et al., 2014). У большинства таксонов подсекции Norvegicae во всех секреторных ходах одной семянки встречается одинаково окрашенный секрет, однако у некоторых представителей (A. arctica ssp. beringensis – остров Уналяшка, A. arctica ssp. psilosantha – Северный Сихотэ-Алинь, A. norvegica – Южный Урал) в разных секреторных ходах одной семянки содержится секрет с разной окраской. Вероятно, это свидетельствует о высокой специализации видов, позволяющей им приспосабливаться к широкому спектру неблагоприятных условий.
Морщинистость поверхности перикарпия у одних видов подсекции Norvegicae является постоянным признаком, а у других видов этот признак варьирует. Так, экземпляры A. arctica ssp. beringensis, A. arctica ssp. psilosantha и A. subarctica из разных мест сбора не различались по морщинистости, а виды A. arctica ssp. ehrendorferi и A. norvegica, собранные в разных точках ареала, различались направлением морщинистости (продольная или продольная и поперечная на одном образце) (см. табл. 1). Вероятно, исходная форма морщинистости – продольная, а появление других видов морщинистости является более поздним приобретением. Про экземпляры с двумя видами морщинистости, можно предположить об их гибридном происхождении.
Согласно определению примитивности и продвинутости типов строения семянок сложноцветных (Melikyan, Muradyan, 1975) исследованные таксоны подсекции Norvegicae являются высоко специализированными и приспособленными к суровым условиям произрастания. Критериям высокоспециализированного вида соответствуют следующие признаки: перикарпий редуцирован (состоит из 2 слоев клеток), имеются эфиромасляные каналы, слизепроизводящие клетки, волоски (только на околоцветнике). Спермодерма многослойная, хорошо сохраняется и согласно А.П. Меликян и Л.Г. Мурадян (Melikyan, Muradyan, 1975) в связи с редукцией слоев перикарпия несет функцию механической защиты. В связи с содержанием маслянистых веществ в стенках и полостях клеток экзотесты, спермодерма может служить еще для защиты зародышей от холода.
Среди изученных представителей подсекции Norvegicae по строению перикарпия выделяются американские виды A. arctica ssp. beringensis, A. comata и A. saxicola. Так A. saxicola стоит согласно полученным данным ближе к предковым формам рода Artemisia, таким как Dendranthema (Tzvelev, 1961). У A. saxicola хорошо развито ослизнение наружных клеточных стенок основных клеток экзокарпия (рис. 3L), что, вероятно, является дериватом предковых форм (рис. 5K). Только у A. saxicola отмечается связь слизесодержащей клетки с ребром, в котором находится секреторный ход со слизью и маслом (рис. 4H). Связь “ослизняющихся” (слизепроизводящих) клеток с ребрами, проводящими пучками и, вероятно, секреторными ходами описана у представителей группы Chrysantheminae, к которой относится Dendranthema (Alexandrov, Savchenko, 1949; Melikyan, Muradyan, 1975). Наиболее близко к A. saxicola стоят виды, также имеющие ослизняющиеся наружные клеточные стенки экзокарпия (A. arctica ssp. arctica, A. arctica ssp. ehrendorferi, A. arctica ssp. psilosantha, A. subarctica, A. norvegica). A. comata, выделяется длинными (длиной во всю семянку) “ослизняющимися” содержащими слизь сегментированными клетками, проводящими пучками, не связанными с секреторными ходами, и широкопросветными механическими клетками (рис. 5E). У A. arctica ssp. beringensis мы не обнаружили полностью зрелых семянок, несмотря на то, что сборы проводились в конце вегетационного сезона. Вероятно, A. arctica ssp. beringensis в связи с суровыми условиями произрастания перешел на вегетативную форму размножения.
Проведенные исследования растений, собранных в различных точках ареала, показали в большинстве случаев, что различия в анатомическом строении семянок незначительные. Вместе с тем обнаружено, что семянки некоторых растений, собранных в разных пунктах, различаются по своему строению. Например, среди образцов A. arctica ssp. ehrendorferi выделяется П-549, собранный на северном склоне Марчеканской сопки. У него отсутствуют секреторные ходы, на поверхности перикарпия имеется множество железок, проводящие пучки мелкие. Слизь у этого экземпляра находится в полости клетки, предположительно в вакуоле, а не в периплазматическом пространстве, как у большинства изученных видов. Среди экземпляров A. arctica ssp. psilosantha выделяется образец П-334 с острова Парамушир. У него нет слизевых клеток, наружные клеточные стенки не ослизняются. У этого образца отмечено и самое большое количество ребер – 7 (табл. 1). Сегментированные клетки на поверхности перикарпия имеются, но в полостях клеток находится масло. В клетках экзотесты маслянистое вещество отмечено, как в клеточных стенках, так и в их полостях. У других образцов A. arctica ssp. psilosantha маслянистое вещество найдено только в клеточных стенках экзотесты. У этого же экземпляра П-334 отсутствует проводящий пучок, связанный с карпоподиумом. Аналогичная картина имеет место и в случае A. norvegica с Южного Урала (9358-521). Образцы семянок этого вида отличаются от остальных наличием смешанной морщинистости на перикарпии (продольная и поперечная), большим количеством ребер (8) в перикарпии, образованием разноокрашенного секрета в разных секреторных ходах, сегментированными слизесодержащими клетками с разрывающимися наружными клеточными стенками, и самым многоклеточным карпоподиумом (7–11 рядов клеток).
Таким образом, проведенное исследование выявило ряд морфолого-анатомических особенностей строения семянок у представителей подсекции Norvegicae секции Abrotanum рода Artemisia. Эти особенности могут представлять интерес как для понимания путей структурной адаптации полыней к различным условиям жизни, так и для решения некоторых вопросов систематики и эволюции в роде Artemisia. В то же время следует отметить, что к использованию структурных особенностей семянок следует относиться с осторожностью. Семянки некоторых растений могут существенно отклоняться по своему строению от семянок остальных представителей этого же вида или подвида. Возможно, это свидетельствует о высокой пластичности представителей рассматриваемой группы растений, которая позволяет им заселять самые разные условия обитания.
Список литературы
[Alexandrov, Savchenko] Александров В.Г., Савченко М.И. 1949. Морфолого-анатомические особенности семянок сложноцветных из трибы Anthemideae как показатель условий их произрастания и местообитания. – Бот. журн. 34 (2): 129–147.
[Alexandrov, Savchenko] Александров В.Г., Савченко М.И. 1951. Об особенностях истории развития плода и семени в семействе сложноцветных. – Тр. БИН им. В.Л. Комарова АН СССР. 7 (2): 5–98.
[Amelchenko] Амельченко В.П. 2006. Биосистематика полыней Сибири. Кемерово. 237 с.
[Boyko] Бойко Э.В. 2011. Таксономия и ресурсы дальневосточных видов семейства Asteraceae: Автореф. дис. …. докт. биол. наук. Владивосток. 42 с.
[Boyko] Бойко Э.В. 2013. Микроморфология семянок видов рода Artemisia (Anthemideae, Asteraceae). – Turczaninowia. 16 (2): 91–105.
[Denisova] Денисова Г.А. 1989. Терпеноидсодержащие структуры растений. Л. 141 с.
Ehendorfer F. 1964. Notizen zur Cytotaxonomie und Evolution der Gattung Artemisia. – Öster. Bot. Zeitsch. 111 (1): 84–142.
Huang Z., Boubriak I., Osborne D.J., Dong M., Gutterman V. 2008. Possible role of pectin-containing mucilage and dew in repairing embryo DNA of seeds adapted to desert conditions. – Ann. Bot. 101 (2): 277–283.
Huang Zh., Gutterman Y., Hu Zh. 2000. Structure and function of mucilaginous achenes of Artemisia monosperma in habiting the Negev desert of Israel. – Israel Journal of Plant Sci. 48: 255–266.
Hulten E. 1954. Artemisia norvegica Fr. and its allies. – Nytt Magasin for Botanikk. Oslo. 3: 63–82.
[Karpova, Yakovleva] Карпова А.П., Яковлева О.В. 2010. Строение семенной кожуры представителей рода Stubendorffia (Cruciferae). – Бот. журн. 95 (5): 37–42.
[Karpova et al.] Карпова А.П., Яковлева О.В., Веселова П.В. 2010. Особенности строения перикарпия и семенной кожуры Sisymbrium subspinescens и S. polymorphum (Cruciferae). – Бот. журн. 95 (12): 1700–1707.
[Karpova, Yakovleva] Карпова А.П., Яковлева О.В. 2012. Сравнительный анализ видов рода Stroganowia (Cruciferae). – Вестн. СПбГУ. 3 (1): 14–37.
[Kolalite] Колалите М.Р. 2001. Ультраструктура смоляных ходов некоторых видов сем. Araliaceae. – Растительные ресурсы. 37 (4): 56–62.
Kolalite M.R., Oskolski A.A., Richter H., Schmitt G.U. 2003. Bark anatomy and intercellular canals in the stem of Delarbrea paradoxa (Araliaceae). – IAWA J. 24 (2): 139–154.
[Korobkov] Коробков А.А. 1973. Морфолого-анатомические особенности семянок полыней (Artemisia ssp.) северо-востока СССР. – Бот. журн. 58 (9): 1302–1315.
[Korobkov] Коробков А.А. 1981. Полыни Северо-Востока СССР. Л. 120 с.
[Krasheninnikov] Крашенинников И.М. 1946. Опыт филогенетического анализа некоторых евразиатских групп рода Artemisia L. в связи с особенностями палеогеографии Евразии. – Матер. по истории флоры и растительности СССР. 2: 87–195.
Kreitschitz A. 2009. Biological Properties of Fruit and Seed Slime Envelope: How to Live, Fly, and Not Die. – In: Functional Surfaces in Biology. Little Structures with Big Effects. 1: 11–30. https://doi.org/10.1007/978-1-4020-6697-92
Kreitschitz A. 2012. Mucilage formation in selected taxa of the genus Artemisia L. (Asteraceae, Anthemideae). – Seed Science Research. 22: 177–189. https://doi.org/10.1017/S0960258512000098
Kreitschitz A., Valles J. 2007. Achene morphology and slime structure in some taxa of Artemisia L. and Neopallasia L. (Asteraceae) – Flora. 202: 570–580. https://doi.org/10.1016/j.flora.2006.12.003
[Lillie] Лилли Р.Д. 1969. Патогистологическая техника и практическая гистохимия (перевод с англ.). М. 645 с.
Ling Y.R. 1992. The old word Artemisia Linn. (Compositae). – Bulletin Botanica research. 12 (1): 1–108.
Ling Y.R. 1995. The new world Artemisia L. – In: Advances in Compositae Systematics. Royal Botanical Gardens, Kew. P. 255–281.
[Melikyan, Muradyan] Меликян А.П., Мурадян Л.Г. 1975. Основные направления эволюции перикарпия и спермодермы в подтрибе Chrysantheminae (Asteraceae). – Бот. журн. 60 (8): 1123–1133.
[Muradyan] Мурадян Л.Г. 1968. Анатомия плода представителей родов Carcrinia, Dendranthema, Spathipappus, Trichanthemis в связи с их систематикой. – Акад. наук Арм. ССР. Биологический журнал Армении. 21 (10): 63–67.
[Muravnik] Муравник Л.Е. 2007. Морфология и ультраструктура трихом перикарпия видов Juglans (Juglandaceae) в связи с синтезом вторичных метаболитов. – Бот. журн. 92 (11): 1720–1732.
Oberprieler C., Himmelreich S., Källersjö M., Vallès J., Watson L.E., Vogt R. 2009. Anthemideae. – In: Systematics, evolution, and biogeography of Compositae. Vienna, Austria. P. 631–666.
O’Brien T.P., Feder N., McCully M.E. 1964. Polyehromatic Staining of Plant Cell Walls by Toluidine Blue O. – Protoplasma. 59: 368–373. https://doi.org/10.1007/bf01248568
Ouyahya A. 1995. Etude anatomique preliminare des akenes de quelques Artemisia dans le basin mediterraneen occidental. – Comp. Newsl. 26: 40–48.
Ouyahya A., Viano J. 1990. Etude au meb du test des akenes du genera Artemisia. – Bol. Soc. Bot. 63: 99–113.
Rydberg A. 1916. Anthemideae. – In: North American Flora. 34 (3): 244–285.
[Savchenko] Савченко М.И. 1949. Применение анатомо-морфологического анализа к выяснению положения рода Brachanthemum DC. в сем. Compositae. – Бот. матер. Гербария Бот. инст. АН СССР. 11: 202–207.
[Tzvelev] Цвелев Н.Н. 1961. Род Ajania Poljakov. – В кн.: Флора СССР. М. Т. 26. С. 398–411.
[Yakovleva et al.] Яковлева О.В., Коробков А.А., Бойко Э.В. 2002. Строение слизепроизводящих клеток в перикарпии семянок некоторых видов Artemisia (Asteraceae). – Бот. журн. 87 (9): 1–16.
[Yakovleva et al.] Яковлева О.В., Тучина Н.Г., Иванова А.Н. 2010. Семейство Goodeniaceae. – В кн.: Сравнительная анатомия семян. Двудольные. Lamiidae, Asteridae. СПб. Т. 7. С. 402–414.
[Yakovleva, Kravtsova] Яковлева О.В., Кравцова Т.И. 1999. Ультраструктура клеток перикарпия Urtica dioica (Urticaceae). – Бот. журн. 84 (7): 33–41.
[Yakovleva] Яковлева О.В. 1988. Слизевые клетки эпидермы листа двудольных растений (данные электронной микроскопии). – Бот. журн. 73 (7): 977–987.
[Yakovleva] Яковлева О.В. 1990. Формирование слизевых клеток эпидермы листа двудольных растений (данные электронной микроскопии). – Бот. журн. 75 (10): 1400–1408.
Yang X., Baskin C.C., Baskin J.M., Zhang W., Huang Z. 2012. Degradation of seed mucilage by soil microflora promotes early seedling growth of a desert sand dune plant. – Plant Cell Environ. 35 (5): 872–883. https://doi.org/10.1111/j.1365-3040.2011.02459.x
Yang X., Zhang W., Dong M., Boubriak I, Huang Z. 2011. The achene mucilage hydrated in desert dew assists seed cells in maintaining DNA integrity: adaptive strategy of desert plant Artemisia sphaerocephala. – PLoS One. 6 (9): e24346.https://doi.org/10.1371/journal.pone.0024346
[Zhigzhitzhapova et al.] Жигжитжапова С.В., Рандалова Т.Э., Раднаева Л.Д. 2014. Биологически активные вещества Artemisia annua L. – Фундаментальные исследования. 12 (11): 2383–2387.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал