

Ботанический журнал, 2022, T. 107, № 7, стр. 627-651
ОСОБЕННОСТИ КОНСТРУКТИВНОЙ ОРГАНИЗАЦИИ ДЕРЕВЬЕВ УМЕРЕННОГО ПОЯСА, ОБУСЛОВЛЕННЫЕ СТРОЕНИЕМ И РИТМОМ РАЗВИТИЯ ГЕНЕРАТИВНЫХ ПОБЕГОВ
М. В. Костина 1, 2, *, Н. С. Барабанщикова 1, **, О. И. Недосеко 3, ***, О. И. Ясинская 4, ****
1 ФГБОУ ВО “МПГУ”
129164 Москва, ул. Кибальчича, 6, корп. 3, Россия
2 ФГАОУ ВО “Севастопольский государственный университет”
299053 Севастополь,
ул. Университетская, 33, Россия
3 Национальный исследовательский Нижегородский государственный университет им. Н.И. Лобачевского, Арзамасский филиал ННГУ
607220 Арзамас, ул. К. Маркса, 36, Россия
4 Главный ботанический сад им. Н.В. Цицина РАН
127276 Москва,
ул. Ботаническая, 4, Россия
* E-mail: mv.kostina@mpgu.su
** E-mail: ns.barabanshchikova@mpgu.su
*** E-mail: nedoseko@bk.ru
**** E-mail: ksen.yasinka@mail.ru
Поступила в редакцию 28.01.2022
После доработки 10.05.2022
Принята к публикации 12.05.2022
- EDN: NLEBVH
- DOI: 10.31857/S0006813622070067
Аннотация
Цель исследования состояла в выявлении особенностей конструктивной организации деревьев умеренного пояса, обусловленных строением и ритмом развития генеративных побегов, а также степенью их участия в построении скелетных осей. Объектами исследования были 14 видов деревьев, составляющих первые ярусы лесных фитоценозов. 7 из них имеют односезонные неолиственные или слабоолиственные генеративные побеги. У 3 видов формируются генеративные побеги с интеркалярным расположением соцветий, стробилов или шишек и полностью сохраняющейся осью, у 4 видов – генеративные побеги с терминальными соцветиями и сохраняющейся осевой частью вегетативной зоны. Ранние сроки цветения определяются отсутствием или небольшим числом листьев срединной формации, развивающихся под терминальным соцветием, расположением пазушных соцветий в проксимальной части генеративных побегов с интеркалярным цветорасположением и формированием генеративных побегов с терминальными соцветиями, зимующими на стадии предсоцветия. Появление в кроне односезонных генеративных побегов и генеративных побегов с интеркалярным расположением не влияет на способ нарастания скелетных осей. Односезонные генеративные побеги могут образоваться как на основе генеративных побегов с терминальным, так и интеркалярным цветорасположением в результате недоразвития почек возобновления. Появление в кроне генеративных побегов с терминальным цветорасположением и с сохраняющейся осевой частью вегетативной зоны вызывает смену моноподиального нарастания на симподиальное. Генеративные побеги деревьев умеренного пояса, как правило, имеют компактные соцветия, небольшие размеры и приурочены к осям высоких порядков. Они либо не участвуют в образовании мощных скелетных осей, либо участвуют, но только на заключительных этапах их формирования. Особенности строения генеративных побегов, их расположения в кроне и смещение сроков цветения на весну-начало лета способствуют сохранению многолетней осевой системы деревьев в условиях короткого вегетационного периода.
Во многих климатических поясах деревья являются эдификаторами зонального типа растительности. Дерево – жизненная форма растений, характеризуемая наличием ствола, сохраняющимся до конца жизни, и кроной, образованной ветвями или пучком крупных листьев. У кронообразующих видов форма кроны, как и сам процесс ее образования, определяется различиями в интенсивности и направлении роста отдельных побегов и формирующихся на их основе осей. Несмотря на единый принцип структурной организации деревьев, эндогенная ритмика развития побегов и побеговых систем у деревьев умеренного пояса находится под жестким контролем экзогенных факторов, что накладывает отпечаток как на габитус деревьев, так и на способы их адаптации к суровым условиям произрастания (Serebryakov, 1962; Hallé, Oldeman, Tomlinson, 1978; Millet, Bouchard, Еdelin, 1998 а, в, 1999).
В 70−90-х годах прошлого века зарубежные исследователи активно развивали представления о конструктивной организации древесных растений на основе изучения деревьев экваториального и бессезонного тропического климата и разработали концепцию архитектурных моделей (Hallé, Oldeman, 1970; Hallé, Oldeman, Tomlinson. 1978).
При выделении архитектурных моделей обращают внимание на наиболее крупные их элементы – ствол и отходящие от ствола мощные оси.
К признакам, по которым различают архитектурные модели, относятся:
1. Характер нарастания осей (ритмическое или постоянное, детерминированное или недетерминированное, моноподиальное, симподиальное или смешанное);
2. Характер ветвления (терминальное, боковое или без ветвления, акротонное, мезотонное или базитонное, силлептическое, пролептическое или из спящих почек);
3. Морфологическая дифференциация побегов и осей по направлению роста (оси ортотропные, плагиотропные, оси с плагиотропным и ортотропным элементами); размерам (длинные, короткие), выполняемым функциям (ростовые, генеративные, ассимилирующие, подземные и т.д.);
4. Расположение соцветий – терминальное или пазушное.
Каждая архитектурная модель определяется сочетанием перечисленных выше морфологических признаков. Хотя теоретически число таких сочетаний весьма велико, в природе признаки группируются чаще только определенным образом. Такие устойчивые, наиболее часто встречающиеся и наиболее стабильные комбинации признаков соответствуют 24 архитектурным моделям.
Основные архитектурные модели возникли в тропиках. По мере удаления от тропиков к северу число архитектурных моделей сокращается, поскольку не все модели могут реализовываться в условиях сурового климата (Hallé, Oldeman,1970; Hallé, Oldeman, Tomlinson, 1978).
Впоследствии с позиций данной концепции была проанализирована структурная организация многих видов деревьев умеренного климата, произрастающих в Европе и в Северной Америке. Было установлено, что древесные растения умеренного пояса, в отличие от большинства деревьев влажных тропических лесов, интенсивнее ветвятся до 5−8 порядков (Hallé, Oldeman, Tomlinson, 1978). Среди деревьев умеренного пояса нет архитектурных моделей, представляющих собой одноствольные неветвящиеся деревья (модели Chamberlain, Corner, Holtum). Поскольку все процессы, связанные с ростом, цветением и плодоношением, резко ограничены временным фактором и прерываются длительным периодом покоя, то в умеренном климате не представлены модели, характеризующиеся непрерывным нарастанием осей. Было показано, что у деревьев умеренного климата конструктивная организация нередко сочетает признаки нескольких моделей – Troll, Rauh, Koriba, Massart (Millet, Bouchard, Édelin, 1998, а, в, 1999; Millet, Bouchard, 2011).
Ряд признаков, используемых для характеристики архитектурных моделей, касающихся положения соцветий на многолетних скелетных осях, требуют уточнения. В рамках концепции архитектурных моделей этот признак рассматривается с точки зрения влияния соцветий на способ нарастания скелетных осей. В том случае, когда соцветие занимает терминальное положение, моноподиальное нарастание оси сменяется на симподиальное, при пазушном положении соцветия ось сохраняет возможность нарастать моноподиально (Barthelemy, Caraglio, 2007). Вместе с тем соцветие как специализированная система силлептически ветвящихся цветоносных осей (Nukhimovsky, 1986; Kuznetsova, 1992) не является самостоятельным конструктивным элементом побегового тела растения, соцветие встраивается в систему растущего побега и является его частью. Такие побеги, включающие соцветия, W. Troll (1964, 1969) называл Blütentrieb, И.Г. Серебряков (Serebryakov, 1952) – цветоносными побегами. В данной работе применительно к этому конструктивному элементу мы используем термин генеративный побег (Kostina, 2005; Kostina et al., 2018). Способ нарастания скелетных осей определяется строением генеративных побегов и их положением на скелетных осях.
Успешное существование деревьев в условиях континентального климата с длительным зимним периодом зависит не только от конструктивной организации вегетативной побеговой сферы растений, но и от сроков цветения, которые во многом обусловлены строением генеративных побегов, а также способом их встраивания в многолетнюю осевую систему растения. Вопрос о факторах, определяющих сроки цветения растений, рассматривался многими исследователями. Предполагается, что раннее цветение дает видам преимущество в конкуренции за успешное опыление и достаточное время для развития плодов и семян, что особенно важно в климатических поясах с коротким вегетационным периодом (Munguia-Rosas et al., 2011; Ehrlen, 2015; Austen et al., 2017). Одним из факторов, обеспечивающих ранние сроки цветения, считается преформация органов в зимующих почках многолетних растений (Schnablova et al., 2021). На связь строения генеративных побегов со степенью преформации и сроками цветения указывал И.Г. Серебряков (Serebryakov, 1949, 1952) и другие исследователи (Bulygin, 1964; Filatova et al., 1986), которые отмечали, что виды, у которых в состав генеративного побега помимо соцветия входят листья срединной формации, цветут обычно позже, чем виды, у которых на генеративных побегах такие листья отсутствуют. Ряд исследователей отмечали зависимость между сроками цветения, способами нарастания побегов и конструктивной организацией растений (Geber, 1990; Duffy et al., 1999; Kudoh et al., 2002).
Цель настоящей работы заключается в обобщении литературных и наших собственных данных, как полученных ранее, так и совсем новых, касающихся конструктивной организации лесообразующих видов деревьев первой величины, произрастающих в средней полосе европейской части России, обусловленных особенностями строения и ритмом развития генеративных побегов, а также степенью их участия в построении скелетных осей.
Задачи исследования:
– выявить особенности строения генеративных побегов, а также их роль в формировании многолетней осевой системы растений;
– установить особенности структурной организации и ритма развития генеративных побегов, обуславливающие ранние сроки цветения;
– рассмотреть модусы преобразования генеративных побегов, обеспечивающие ранние сроки цветения;
– провести сопоставление конструктивной организации деревьев умеренного пояса и тропического по признакам, связанным с различными вариантами встраивания соцветий в вегетативное тело растения.
МАТЕРИАЛ И МЕТОДЫ
Объектами исследования были Acer platanoides L. (Aceraceae), Alnus glutinosa (L.) Gaertn., Alnus incana (L.) Moench, Betula pendula Ehrh., Betula pubescens Ehrh., (Betulaceae), Fraxinus excelsior L. (Oleaceae), Picea abies (L.) Karst., Pinus sylvestris L. (Pinaceae), Populus tremula L., Salix alba L., Salix pentandra L. (Salicaceae), Quercus robur L. (Fagaceae), Tilia cordata Mill. (Tiliaceae), Ulmus glabra Huds., Ulmus laevis Pall. (Ulmaceae).
Таблица 1.
Признаки генеративных побегов (ГП) аборигенных видов деревьев первой величины, произрастающих в Средней полосе европейской части России Table 1. Features of generative shoots (GSh) of native first size tree species occurring in the Central European part of Russia
| Вид Species |
ГП полностью отмирают после
цветения и плодоношения GSh die completely after flowering and fruiting |
После цветения и плодо-
ношения на ГП остается резид Wood base remains on GSh after flowering and fruiting |
Терминальное расположение
соцветий или шишек Terminal location of inflorescences or cones |
Интеркалярное расположение
соцветий или шишек Intercalary location of inflorescences or cones |
Наличие развитых листьев
в вегетативной зоне ГП Developed leaves present in vegetative zone of GSh |
ГП образуются из
пазушных почек GShs are formed from axillary buds |
ГП образуются из
верхушечных почек GShs are formed from apical buds |
Есть стадия предсоцветия Stage of pre-inflorescence exists |
Ветроопыление Wind pollination |
Влияние ГП на характер
нарастания осей GShs impact on the pattern of axis development |
|---|---|---|---|---|---|---|---|---|---|---|
| Acer platanoides | нет/no | да/yes | да/yes | нет/no | да/yes | да/yes | да/yes | нет/no | нет/no | да/yes |
| Alnus glutinosa | нет/no | да/yes | да/yes | нет/no | да/yes | да/yes | да/yes | да/yes | да/yes | да/yes |
| Betula pendula (мужские/male) | нет/no | да/yes | да/yes | нет/no | да/yes | да/yes | нет/no | да/yes | да/yes | да/yes |
| Betula pendula (женские / female) | нет/no | да/yes | да/yes | нет/no | да/yes | да/yes | да/yes | нет/no | да/yes | да/yes |
| Betula pubescens (мужские / male) | нет/no | да/yes | да/yes | нет/no | да/yes | да/yes | нет/no | да/yes | да/yes | да/yes |
| Betula pubescens (женские / female) | нет/no | да/yes | да/yes | нет/no | да/yes | да/yes | да/yes | нет/no | да/yes | да/yes |
| Fraxinus excelsior | да/yes | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no |
| Picea abies (мужские / male) | да/yes | нет/no | да/yes | нет/no | нет/no | да/yes | редко/seldom | нет/no | да/yes | редко/seldom |
| Picea abies (женские / female) | да/yes | нет/no | да/yes | нет/no | да/yes | да/yes | да/yes | нет/no | да/yes | часто/frequently |
| Pinus sylvestris | нет/no | да/yes | нет/no | да/yes | да/yes | да/yes | нет/no | нет/no | да/yes | нет/no |
| Populus tremula | да/yes | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no |
| Quercus robur (1-й вариант/1st variant) | да/yes | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no |
| Quercus robur (2-й вариант/2nd variant) | нет/no | да/yes | нет/no | да/yes | да/yes | да/yes | да/yes | нет/no | да/yes | нет/no |
| Salix alba | да/yes | нет/no | да/yes | нет/no | да/yes | да/yes | нет/no | нет/no | нет/no | нет/no |
| S. pentandra | да/yes | нет/no | да/yes | нет/no | да/yes | да/yes | нет/no | нет/no | нет/no | нет/no |
| Tilia cordata | нет/noo | да/yes | нет/no | да/yes | нет/no | да/yes | нет/no | нет/no | нет/no | нет/no |
| Ulmus glabra | да/yes | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no |
| Ulmus laevis (1-й вариант/1st variant) | да/yes | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no | нет/no | да/yes | нет/no |
| U. laevis (2-й вариант/2nd variant) | нет/no | да/yes | нет/no | да/yes | да/yes | да/yes | нет/no | нет/no | да/yes | нет/no |
Структурно-ритмологический анализ побеговых систем осуществляли путем выделения в теле растения конструктивных элементов. В качестве таковых использовали вегетативные и генеративные побеги. У всех изученных видов почки закрытые, элементарные побеги были хорошо различимы благодаря наличию в их основании зоны укороченных междоузлий и почечных чешуй.
Определения терминов, используемых для описания побегов и систем побегов
Пролепсис – развитие боковых побегов происходит в том же сезоне, что и образование материнского побега, но после непродолжительного периода покоя. В основании пролептического побега находятся короткие междоузлия и чешуевидные листья (Müller-Doblies, Weberling, 1984).
Силлепсис – развитие боковых побегов происходит одновременно с образованием материнского побега (Späth, 1912; Grudzinskaya, 1960; Müller-Doblies, Weberling, 1984). В основании силлептического побега обычно отсутствуют укороченные междоузлия и чешуевидные листья (Tomlinson. Gill, 1973; Hallé et al., 1978; Müller-Doblies, Weberling, 1984). Синхронность развития материнского и силлептических побегов дает основание рассматривать их не как отдельные самостоятельные элементарные побеги, а как составные элементы единого разветвленного побега, образующегося за один цикл роста (Mikhalevskaya, 2002).
Вегетативный побег – конструктивный элемент, образующийся за один цикл видимого роста из зимующей или спящей почки и включающий стебель, листья и почки. В русскоязычной литературе такие побеги называют элементарными побегами (Grudzinskaya, 1960), а за рубежом единицами роста (англ.: unit groth) или единицами прироста (англ.: unit of extension) (Barthelemy, Caraglio, 2007). В случае силлептического ветвления за один цикл роста формируется разветвленный вегетативный побег.
Генеративный побег – конструктивный элемент, образующийся за один цикл видимого роста из зимующих или спящих почек и включающий, помимо стебля, листьев и почек, еще и цветки (Kostina, 2005; Kostina et al., 2018). Поскольку у растений умеренного пояса соцветие обычно развивается в результате силлептического ветвления (Kusnetzova, 1988; Kusnetzova, 1992; Nukhimovsky, 1986), то генеративный побег – частный случай силлептически разветвленного побега. В морфологии растений нередко используют термины “генеративный побег” и “вегетативно-генеративный побег”. Однако мы считаем, что при описании побегов, в состав которых входят генеративные структуры, наличие или отсутствие листьев срединной формации является важным, но не единственным признаком. Не менее значимые признаки – это положение соцветий на побеге (терминальное или пазушное), характер отмирания и возобновления генеративного побега после цветения и плодоношения, наличие или отсутствие резида, степень специализации побега и т.д. Поэтому понятие “генеративный побег” более универсально, а “вегетативно-генеративный побег” – один из его вариантов.
Соцветие – достаточно целостная и отчетливо отграниченная от вегетативных частей растения группировка цветков, образовавшаяся в результате силлептического ветвления (Nukhimovsky, 1986; Kusnetzova, 1988; Kusnetzova, 1992). Соцветие в системе генеративного побега может занимать терминальное или интеркалярное положение.
Терминальное цветорасположение. Соцветие завершает собой генеративный побег. У голосеменных терминальное положение в системе генеративных побегов могут занимать стробилы или шишки.
Генеративная зона – отрезок главной оси генеративного побега, располагающийся между вегетативными участками, на котором развиваются пазушные цветки, пазушные соцветия, стробилы или шишки.
Интеркалярное цветорасположение – расположение пазушных цветков или соцветий на моноподиально нарастающей оси (Ims., 1964; Weberling, 1981, 1982). Мы рассматривали вариант, при котором генеративная зона располагается на главной оси генеративного побега, т.е. на приросте текущего года между вегетативными участками (Kuznetsova, Timonin, 2017). У голосеменных растений интеркалярное положение на оси генеративного побега могут занимать стробилы или шишки.
Предсоцветие – стадия развития соцветия, на которой завершившее внутрипочечный этап развития соцветие выходит из почки, зимует голым (не защищенным почечными чешуями) и переходит к цветению только весной следующего года (Nukhimovsky, 1997).
Вегетативная зона – часть генеративного побега, включающая обычно листья срединной формации, зимующие и спящие почки, а иногда и силлептические вегетативные побеги.
Резид – многолетняя осевая часть генеративного побега с зимующими и спящими почками (Nukhimovsky, 1997).
В качестве конструктивных элементов использовали также скелетные оси видимого порядка, т.е. части растения, которые имеют по одному основанию и по одной видимой верхушке, служат опорами для других частей, формируются в результате мноподиального или симподиального нарастания побегов и существуют более одного года (Nukhimovsky, 1971; Barthelemy, Caraglio, 2007).
Скелетные оси видимых порядков являются теми реальными структурными элементами, которые можно найти у всех деревьев и которые позволяют осуществлять сравнительно-морфологические манипуляции. Ось I видимого порядка – ствол, от ствола отходят оси II видимого порядка и т.д.
При изучении генеративных побегов выявляли характер цветорасположения (терминальное, пазушное), строение вегетативной зоны, особенности отмирания генеративных побегов после пыления и образования семян.
Учитывали положение почек (верхушечное, пазушное), из которых развиваются генеративные побеги, а также характер материнских побегов (вегетативные, генеративные), на которых эти почки закладываются.
Определяли степень участия генеративных побегов в построении скелетных осей I и мощных скелетных осей II порядка.
Составляли схемы строения генеративных побегов, а также схемы строения двухлетних побеговых систем, в образовании которых принимали участие генеративные побеги.
РЕЗУЛЬТАТЫ
У всех изученных нами видов деревьев зимующие почки как вегетативные, так и генеративные закрытые, т.е. защищены почечными чешуями. В течение вегетационного периода обычно наблюдается один цикл ростовых процессов, и границы годичных вегетативных побегов совпадают с границами элементарных побегов. Исключение представляют Quercus robur и Populus tremula, у которых можно наблюдать образование двух, реже – более, элементарных побегов. Для большинства видов, за исключением Betula pendula, Alnus glutinosa, Salix alba, не характерно силлептическое ветвление вегетативных побегов. Размеры вегетативных побегов в пределах кроны сильно варьируют в зависимости от их положения в кроне и функций.
Генеративные побеги Quercus robur
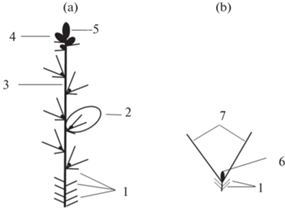
У Quercus robur могут формироваться четыре варианта генеративных побегов.
1. Генеративные побеги длиной 10–30 см образуются из верхушечных и пазушных почек, состоят из стебля, завершающегося верхушечной почкой, листьев срединной формации (5 и более) с пазушными почками регулярного возобновления, мужских и женских сережек, почечных чешуй с расположенными в их пазухах спящими почками. Мужские сережки (открытый брактеозный тирс) находятся в пазухах верхних почечных чешуй и нижних листьев срединной формации, а женские (открытый брактеозный колос) – формируются в дистальной части генеративного побега, в пазухах листьев срединной формации. Такие генеративные побеги могут развиваться как из верхушечных, так и из пазушных почек. Мужские сережки входят в состав генеративной зоны, образующейся в проксимальной части генеративного побега, а женские соцветия – в генеративную зону, расположенную в дистальной части.
Первыми переходят к цветению мужские сережки, позже зацветают женские. После цветения и плодоношения пазушные соцветия отмирают, а вся осевая часть генеративного побега входит в состав многолетней осевой системы растения. Благодаря верхушечной почке, побеги сохраняют способность к дальнейшему моноподиальному нарастанию (рис. 1а).
Рис. 1.
Генеративные побеги Quercus robur.
а – Генеративные побеги с мужскими и женскими соцветиями.
b – Генеративные побеги с женскими соцветиями.
с – Генеративные побеги с мужскими соцветиями.
d – Генеративные односезонные побеги с мужскими соцветиями.
1 – Почечные чешуи; 2 – спящие почки; 3 – мужское соцветие; 4 – почка регулярного возобновления; 5 –лист срединной формации; 6 – женское соцветие; 7 – терминальная почка; 8 – флоральная зона с пазушными мужскими соцветиями; 9 – флоральная зона с пазушными женскими соцветиями; 10 – гибель верхушечной меристемы. Толстой линией обозначены многолетние оси, тонкой – однолетние.
Fig. 1. Generative shoots in Quercus robur.
а – Generative shoots with male and female inflorescences.
b – Generative shoots with female inflorescences.
с – Generative shoots with male inflorescences.
d – Generative single-season shoots with male inflorescences.
1 – Bud scales; 2 – dormant buds; 3 – male inflorescence; 4 – bud of regular renewal; 5 – foliage leaf; 6 – female inflorescence; 7 – terminal bud; 8 – floral zone with male axillary inflorescences; 9 – floral zone with female axillary inflorescences; 10 – apical meristem cell death. Perennial axes are represented with thick lines, annual axes are represented with thin lines.

Более ранние сроки цветения мужских цветков по сравнению с женскими обусловлены тем, что у Quercus robur мужские цветки в почках начинают закладываться в июле, а женские – лишь весной следующего года (Minina, 1954; Stairs, 1964).
2. На генеративных побегах длиной 10–30 см, образующихся из верхушечных и пазушных почек, развиваются только женские сережки (рис. 1b).
3. На генеративных побегах длиной 2–10 см, формирующихся из верхушечных и пазушных почек, развиваются, в отличие от первого варианта, только мужские сережки (рис. 1b).
У побегов 2 и 3 типа, как и у побегов 1 типа, вся осевая часть генеративного побега входит в состав многолетней осевой системы растения.
4. На генеративных побегах длиной 1–2 см образуются почечные чешуи и мужские сережки. Такие генеративные побеги после пыления полностью отмирают, т.е. являются односезонными (рис. 1d). Они обычно развиваются из почек, расположенных в непосредственной близости от верхушечной почки вегетативного или генеративного побега.
Генеративные побеги с женскими сережками принимают участие в построении скелетных осей. Однако мощные скелетные оси у Q. robur сначала формируются на основе вегетативных побегов, а затем в их образовании начинают участвовать генеративные побеги. Появление генеративных побегов, имеющих 1–3-й варианты строения, не влияет на характер нарастания скелетных осей, поскольку эти побеги завершаются вегетативными верхушечными почками (рис. 2a, 2b).
Рис. 2.
Формирование двулетней побеговой системы с мужскими и женскими сережками у Quercus robur.
a – Строение генеративного побега.
b – Строение двулетней побеговой системы.
1 – Почечное кольцо; 2 – мужское соцветие; 3 – лист срединной формации; 4 – вегетативная почка регулярного возобновления; 5 – женское соцветие; 6 – пазушная почка с мужскими соцветиями; 7 – верхушечная почка; 8 – прошлогоднее опавшее мужское соцветие; 9 – прошлогоднее отмершее женское соцветие; 10 – вегетативные побеги; 11 – односезонный побег с мужскими сережками.
Fig. 2. Formation of biennial shoot system with male and female catkins in Quercus robur.
a – Structure of generative shoot.
b – Structure of biennial shoot system.
1 – Bud scar; 2 – male inflorescence; 3 – foliage leaf; 4 – vegetative bud of regular renewal; 5 – female inflorescence; 6 – axillary bud with male inflorescences; 7 – apical bud; 8 – abscissed male inflorescence of last year; 9 – dead female inflorescence of last year; 10 – vegetative shoots; 11 – single-season shoot with male catkins.

Генеративные побеги Pinus sylvestris
Для того, чтобы было понято, как устроены генеративные побеги у Pinus sylvestris, рассмотрим сначала разветвленные вегетативные побеги, формирующиеся из почек регулярного возобновления. В их состав входят стебель, стерильные чешуевидные листья, находящиеся в основании стебля, укороченные силлептические побеги (брахибласты), верхушечная и несколько пазушных почек возобновления (рис. 3а). Брахибласты состоят из стебля, стерильных почечных чешуй, двух листьев срединной формации и верхушечной почки. Брахибласты и почки возобновления находятся в пазухах чешуевидных листьев (рис. 3b).
Рис. 3.
Строение вегетативного побега Pinus sylvestris.
а – Строение брахибласта.
b – Строение вегетативного побега.
1 – Стебель брахибласта; 2 – чешуевидные листья в основании брахибласта; 3 – верхушечная почка брахибласта; 4 – листья срединной формации (хвоинки); 5 – чешуевидные листья в основании вегетативного побега; 6 – стебель ауксибласта; 7 – брахибласт; 8 – верхушечная вегетативная почка; 9 – пазушные вегетативные почки; 10 – спящие почки.
Fig. 3. Structure of vegetative shoot in Pinus sylvestris.
а – Structure of brachyblast.
b – Structure of vegetative shoot.
1 – Stem of brachyblast; 2 – cataphylls at the brachyblast base; 3 – apical bud of brachyblast; 4 – foliage leaves (needles); 5 – cataphylls at the base of vegetative shoot; 6 – stem of auxiblast; 7 – brachyblast; 8 – apical vegetative bud; 9 – axillary vegetative buds; 10 – dormant buds.
У Pinus sylvestris образуется три варианта генеративных побегов с мужскими стробилами и женскими шишками, развивающихся из пазушных и верхушечных почек.
1. Генеративные побеги с мужскими стробилами. Генеративные побеги длиной 3–15 см отличаются от вегетативных побегов тем, что в их проксимальной части формируются мужские стробилы, образующие генеративную зону. После пыления мужские стробилы отмирают (рис. 4а, 4b).
Рис. 4.
Формирование двулетних побеговых систем Pinus sylvestris, в образовании которых принимают участие генеративные побеги.
а – Генеративный побег с мужскими шишками.
b – Двулетняя побеговая система с мужскими шишками.
с – Генеративный побег с женскими шишками.
d – Двулетняя побеговая система с женскими шишками.
1 – Чешуевидные листья; 2 – мужские шишки; 3 – брахибласт; 4 – вегетативная пазушная почка; 5 – верхушечная вегетативная почка; 6 – флоральная зона с мужскими шишками; 7 – опавшие прошлогодние мужские шишки; 8 – вегетативные побеги, образующиеся из пазушных и верхушечной почек; 9 – флоральная зона с женскими шишками; 10 – женская шишка первого года существования; 11 – двулетняя мужская шишка.
Fig. 4. Formation of biennial shoot systems in Pinus sylvestris, involving generative shoots.
а – Generative shoot with male cones.
b – Biennial shoot system with male cones.
с – Generative shoot with female cones.
d – Biennial shoot system with female cones.
1 – Cataphylls; 2 – male cones; 3 – brachyblast; 4 – axillary vegetative bud; 5 – apical vegetative bud; 6 – floral zone with male cones; 7 – abscissed male cones of last year; 8 – vegetative shoots developed from axillary and apical buds; 9 – floral zone with female cone; 10 – annual female cone; 11 – biennial male cone.

2. Генеративные побеги длиной от 15 до 50 см с женскими шишками. Женские шишки формируются под верхушечной почкой, образуя генеративную зону в дистальной части генеративных побегов. Поскольку женские шишки у Pinus sylvestris созревают два года, они сохраняются в системе двулетней побеговой системы, после чего опадают (рис. 4c, 4d).
3. Генеративные побеги с женскими шишками и мужскими стробилами. Такие генеративные побеги у Pinus sylvestris образуются редко.
Мужские стробилы созревают раньше женских шишек. Такая динамика развития связана с тем, что у Pinus sylvestris в конце августа – начале сентября в почках уже имеются зачатки мужских стробилов с примордиями микроспорангиев, в которых находится спорогенная ткань. В отличие от мужских стробилов, примордии женских шишек в год заложения остаются слабо дифференцированными (Mikhalevskaya, 1962; Kozubov, 1974).
В образовании мощных скелетных осей сначала принимают участие вегетативные побеги, а на заключительных этапах развития осей – и генеративные. При этом сохраняется моноподиальный характер нарастания, благодаря формированию на генеративных побегах верхушечной вегетативной почки.
Генеративные побеги Tilia cordata
У Tilia cordata генеративные побеги длиной 2–70 см состоят из стебля, почечных чешуй со спящими почками в их пазухах, листьев срединной формации и пазушных соцветий, представляющих собой дихазии или многочленные дихазии из 3–9 цветков.
На генеративных побегах, так же как и на вегетативных, верхушечная зимующая почка не формируется, но сохраняется способность к акросимподиальному нарастанию, благодаря формированию вегетативного побега замещения. В зимующих почках зачатки пазушных соцветий, как правило, отсутствуют и образуются в год цветения в начале второй декады мая (Bulygin, 1964). Пазушные соцветия появляются в текущем году, в процессе развития материнского побега, в акропетальной последовательности. Генеративная зона, образованная пазушными соцветиями, располагается между нижней и верхней вегетативными зонами (рис. 5а).
Рис. 5.
Генеративные побеги Tilia cordata.
а – Генеративный побег летом в период цветения.
b – Генеративный побег в конце вегетационного периода.
с – соцветие с плодами и α предлистом, отрывающееся от генеративного побега.
1 – Почечная чешуя; 2 – спящая почка; 3 – лист срединной формации; 4 – почка регулярного возобновления; 5 – α‑предлист; 6 – β-предлист; 7 – зона срастания α-предлиста с осью пазушного соцветия; 8 – почка регулярного возобновления; 9 – цветок; 10 – нижняя вегетативная зона; 11 – верхняя вегетативная зона; 12 – флоральная зона; 13 – завершение моноподиального нарастания; 14 – отмерший лист; 15 – соцветие с плодами и α‑предлистом; 16 – резид с почкой регулярного возобновления в пазухе β-предлиста; 17 – соцветие с плодами и α‑предлистом.
Fig. 5. Generative shoots of Tilia cordata
а – Generative shoot in summer, at the flowering stage.
b – Generative shoot at the end of the growing season.
с – Inflorescence with fruits and prophyll α, separating from generative shoot.
1 – Bud scale; 2 – dormant bud; 3 – foliage leaf; 4 – bud of regular renewal; 5 – prophyll α; 6 – prophyll β; 7 – cohesive zone between prophyll α and the axis of axillary inflorescence; 8 – bud of regular renewal; 9 – flower; 10 – lower vegetative zone; 11 – upper vegetative zone; 12 – floral zone; 13 — monopodial growth termination; 14 – dead leaf; 15 – inflorescence with fruits and prophyll α; 16 – basal part of the axis with the bud of regular renewal in the axil of prophyll β; 17 – inflorescence with fruits and prophyll α.

В основании пазушного цветоноса формируются два предлиста. Листовидный α-предлист, срастаясь с осью цветоноса, превращается в крыло (Troll, 1957). В пазухе чешуевидного предлиста закладывается почка регулярного возобновления (рис. 5b). После образования плодов от пазушного цветоноса отделяется соцветие с прицветным крылом (рис. 5с). Остается резид длиной 0.2 см с почкой регулярного возобновления, из которой на следующий год развивается генеративный или вегетативный побег.
Появление генеративных побегов у Tilia cordata не влияет на акросимподиальный характер нарастания осей.
Генеративные побеги Acer platanoides
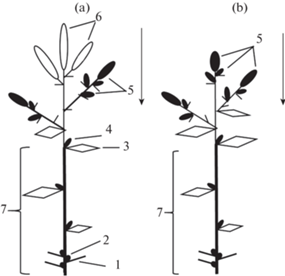
Генеративные побеги длиной 8–19 см имеют стебель, почечные чешуи с пазушными спящими почками, листья срединной формации (две, реже одна пара) и завершаются соцветием метелкой длиной до 12 см. В пазухах верхних листьев обычно формируются силлептические побеги с верхушечной генеративной, реже вегетативной почкой и одной парой листьев срединной формации, в пазухах которых могут закладываться генеративные почки. На мощных генеративных побегах силлептические побеги могут достигать в длину более 20 см, на них развивается 2–4 пары листьев срединной формации. После плодоношения соцветие отмирает, а осевая часть побега с почками возобновления и силлептическими побегами (резид) входит в состав многолетней системы растения (рис. 6а, 6b). Резид – 1–7 см (без учета длины силлептических побегов) (Mikhalevskaya, 2001; Kiseleva, 2003; Kostina, 2008).
Рис. 6.
Формирование двулетней побеговой системы Acer platanoides на основе генеративных побегов.
a – Строение генеративных побегов Acer platanoides.
b – Строение двулетней побеговой системы, формирующийся на основе генеративных побегов.
1 – Почечные чешуи; 2 – спящие почки; 3 – лист срединной формации; 4 – силлептический побег; 5 – генеративная верхушечная почка; 6 – верхушечное соцветие; 7 – вегетативная зона генеративного побега; 8 – отмершее соцветие; Жирной линией показаны многолетние оси, тонкой – отмирающие в конце вегетационного сезона.
Fig. 6. Formation of biennial shoot system in Acer platanoides, involving generative shoots.
a – Structure of generative shoots in Acer platanoides.
b – Structure of biennial shoot system formed of generative shoots.
1 – Bud scales; 2 – dormant buds; 3 – foliage leaf; 4 – sylleptic shoot; 5 – generative apical bud; 6 – terminal inflorescence; 7 – vegetative zone of generative shoot; 8 – dead inflorescence; Perennial axes are represented with thick lines, and those withering away at the end of the growing season are represented with thin lines.

После вступления растения в период цветения и плодоношения генеративные побеги принимают существенное участие в формировании осей III и последующих видимых порядков. В образовании оси I порядка (ствола) и наиболее мощных скелетных осей II порядков растения генеративные побеги принимают участие только на заключительных этапах формирования этих осей. Их появление вызывает смену моноподиального нарастания на симподиальное (Kostina, 2008). Далее на основе генеративных побегов начинают формироваться ложнодихотомические системы, хорошо заметные в кроне генеративных растений (рис. 7).
Рис. 7.
Формирование скелетной оси Acer platanoides, в образовании которой принимают участие генеративные побеги.
1 – Почечное кольцо; 2 – вегетативные побеги; 3 – отмершее соцветие; 4 – соцветие текущего года; 5 – силлептические побеги.
Fig. 7. Skeletal axis formation in Acer platanoides, involving generative shoots.
1 – Bud scar; 2 – vegetative shoots; 3 – dead inflorescence; 4 – inflorescence of current year; 5 – sylleptic shoots.

В зимующих почках у Acer platanoides присутствуют не только зачатки листьев вегетативной зоны, но и зачаточные силлептические побеги, расположенные под зачаточным соцветием (Mikhalevskaya, 2002).
Генеративные побеги Betula pendula и Betula pubescens
Генеративные побеги с мужскими сережками длиной от 5 до 35 см, имеют стебель, почечные чешуи со спящими почками в их пазухах, листья срединной формации с пазушными генеративными и вегетативными почками возобновления и завершаются терминальной и пазушными мужскими сережками (открытые брактеозные тирсы) (рис. 8а). У плакучих форм B. pendula под мужскими сережками может развиваться силлептический побег до 50 см в длину (Jäger, 1980; Kostina, 2001). Сережки начинают формироваться в конце мая. К зиме мужские цветки полностью развиты, но пылят весной следующего года. Таким образом, образовавшиеся на генеративных побегах соцветия с мужскими цветками зимуют на стадии предсоцветия и в зимний период защищены плотно сомкнутыми сросшимися прицветниками и прицветничками.
Рис. 8.
Строение генеративных побегов Betula pendula и Betula pubescens с мужскими сережками (а) и с женскими сережками (b).
1 – Спящие почки; 2 – почечные чешуи; 3 – лист срединной формации; 4 – почка с женской сережкой; 5 – почка, из которой образуется побег с мужскими сережками; 6 – вегетативная почка; 7 – пазушные мужские сережки 8 – терминальная мужская сережка; 9 – терминальная женская сережка, 10 – вегетативная зона генеративного побега с почками регулярного возобновления.
Fig. 8. Structure of generative shoots in Betula pendula and Betula pubescens with male (а) and female catkins (b).
1 – Dormant buds; 2 – bud scales; 3 – foliage leaf; 4 – bud with female catkin; 5 – bud giving rise to the shoot bearing male catkins; 6 – vegetative bud; 7 – male axillary catkins; 8 – male terminal catkin; 9 – female terminal catkin, 10 – vegetative zone of generative shoot with buds of regular renewal.

Генеративные побеги с женскими сережками длиной до 2–4 см, имеют стебель, почечные чешуи со спящими почками в их пазухах, листья срединной формации (2–3) и завершаются терминальной женской сережкой (открытый брактеозный тирс), реже еще и двумя-тремя пазушными сережками (Jäger, 1980; Kostina, 2001; Mikhalevskaya et al., 2006). В вегетативной зоне длиной до 1 см в пазухах листьев обычно закладываются генеративные почки с женскими сережками, реже вегетативные почки (рис. 8b). Женские сережки зимуют в почках, т.е. защищены почечными чешуями. К концу вегетационного периода в год, предшествующий цветению, в женских цветках только начинают развиваться семязачатки (Korchaginа, 1980).
Генеративные почки с женскими соцветиями могут развиваться из пазушных почек, закладывающихся на вегетативных побегах, генеративных побегах с мужскими сережками и на генеративных побегах с женскими сережками. На укороченных вегетативных побегах генеративные побеги с женскими сережками могут формироваться из верхушечных почек (Mikhalevskaya, 2002). На основе генеративных побегов с женскими сережками формируются короткие симподиально нарастающие в течение 2–4 лет веточки, появление которых в кроне практически никак не сказывается на внешнем облике деревьев.
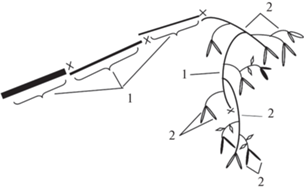
Генеративные побеги с мужскими сережками принимают участие в формировании скелетных осей, в том числе ствола и наиболее мощных осей II порядка. Однако, сначала в образовании таких симподиально нарастающих осей участвуют удлиненные вегетативные побеги, а затем побеги с мужскими сережками, которые, чередуясь с вегетативными, образуют рыхлые повисающие симподиально нарастающие системы (рис. 9). Генеративные и вегетативные побеги в таких системах практически перестают утолщаться (Serebryakov, 1962). Таким образом, образование плакучей части ветвей, определяющих габитус генеративного дерева, связано с регулярным появлением в кроне побегов с мужскими сережками (Kostina et al., 2015; Kostina et al., 2015).
Рис. 9.
Строение симподиально нарастающей скелетной оси Betula pendula, завершающейся системой вегетативных побегов и побегов с мужскими сережками.
1 – Вегетативные побеги; 2 – отмершие мужские сережки; 3 – мужские сережки текущего года.
Fig. 9. Structure of sympodially growing skeletal axis in Betula pendula, terminating in the system of vegetative shoots and shoots bearing male catkins.
1 – Vegetative shoots; 2 – dead male catkins; 3 – male catkins of current year.
У Betula pendula помимо типичного расположения мужских сережек, завершающих собой годичные побеги, существует большое разнообразие реже образующихся вариантов генеративных побегов, возникающее за счет силлептического и пролептического ветвления, благодаря которому увеличивается количество мужских и женских сережек. Так, например, высокая интенсивность пыления этого вида в 2012 г. была ответной реакцией на аномально жаркое и сухое лето 2010 г. (Kostina et al., 2015). Поскольку редкие варианты генеративных побегов у этого вида формируются в экстремальных погодных условиях, в данной статье мы их не рассматриваем.
Генеративные побеги Alnus glutinosa и Alnus incana
Генеративные побеги длиной 5–40 см имеют стебель, почечные чешуи со спящими почками в их пазухах, листья срединной формации (3–10), в пазухах которых закладываются генеративные и вегетативные почки. Завершаются генеративные побеги терминальным мужским соцветием, ниже которого развивается несколько боковых (3–5). Мужские соцветия представляют собой открытые тирсы, которые выглядят как сережки на ножках. Под мужскими сережками в том же вегетационном сезоне развиваются боковые оси с тремя-десятью женскими соцветиями – открытыми сидячими тирсами, которые выглядят как “шишки” (рис. 10а). Таким образом, развитие мужских и женских соцветий на генеративных побегах происходит в базипетальной последовательности: сначала появляются мужские соцветия, а затем женские (Jäger, 1980; Kostina, 2001). И те и другие зимуют на стадии предсоцветия, т.е. в зимний период цветки в таких соцветиях защищены не почечными чешуями, а элементами соцветия – прицветниками и прицветничками, которые срастаются между собой и пропитываются смолами. Мужские соцветия начинают формироваться в начале лета. К зиме мужские цветки полностью развиты и готовы к цветению. В женских соцветиях, образующихся позже мужских, к зиме цветки развиты слабо. Пыление происходит в апреле-начале мая, до распускания листьев. Оплодотворение осуществляется через 1–4 месяца после созревания зародышевого мешка (Korchaginа, 1980). После пыления сразу же опадают мужские соцветия, а женские соцветия после высыпания плодов еще некоторое время висят, а затем опадают. Остается резид длиной до 30 см.
Рис. 10.
Строение генеративных побегов Alnus glutinosa.
а – Строение генеративного побега с мужскими и женскими соцветиями.
b – Строение генеративного побега с женскими соцветиями.
1 – почечные чешуи; 2 – спящие почки; 3 – развитые листья; 4 – почки регулярного возобновления; 5 – женские соцветия; 6 – мужские соцветия; 7 – вегетативная зона генеративного побега. Стрелками показан порядок формирования соцветий.
Fig. 10. Structure of generative shoots in Alnus glutinosa.
а – Structure of generative shoot with male and female inflorescences.
b – Structure of generative shoot with female inflorescences.
1 – Bud scales; 2 – dormant buds; 3 – developed leaves; 4 – buds of regular renewal; 5 – female inflorescences; 6 – male inflorescences; 7 – vegetative zone of generative shoot. Arrows show the order of the inflorescence development.
Систему мужских и женских соцветий, формирующуюся на генеративных побегах, можно описать как соцветие метелка из мужских и женских открытых тирсов.
Реже у Alnus glutinosa формируются генеративные побеги длиной от 5 до 40 см, завершающиеся только системой женских соцветий, состоящей из терминального женского, зацветающего первым, и ниже расположенных боковых осей с женскими сережками, появляющихся и переходящих к цветению в базипетальной последовательности (рис. 10b). Развитие терминальной системы женских сережек начинается одновременно с развитием боковых осей с женскими соцветиями, образующимися на генеративных побегах под мужскими сережками. Таким образом, мужские сережки начинают формироваться раньше женских, вне зависимости от их положения в системе генеративного побега.
Генеративные побеги развиваются из верхушечных и пазушных почек. Появление генеративных побегов из верхушечной почки приводит к смене нарастания скелетных осей с моноподиального на симподиальное и к формированию симподиально нарастающих систем побегов.
Генеративные побеги Fraxinus excelsior
Генеративные побеги длиной 4–8 см (во время цветения) имеют стебель, почечные чешуи и завершаются метелкой. В вегетативной зоне листья срединной формации не образуются. Генеративные побеги появляются только из пазушных почек. Цветение происходит до распускания листьев. Генеративные побеги односезонные (рис. 11а). В зимующих почках генеративный побег сформирован полностью, включая соцветие (Kostina, 2010).
Рис. 11.
Односезонные генеративные побеги.
a – Генеративные побеги Fraxinus excelsior.
b – Генеративные побеги Salix alba, S. pentandra.
c – Генеративные побеги мужских и женских растений Populus tremula.
d – Брактеозная открытая кисть.
1 – Почечные чешуи; 2 – катафиллы; 3 – вегетативная зона; 4 – цветки; 5 – двукилевая почечная чешуя; 6 – развитые листья; 7 – открытая брактеозная кисть.
Fig. 11. Single-season generative shoots.
a – Generative shoots of Fraxinus excelsior.
b – Generative shoots of Salix alba, S. pentandra.
c – Generative shoots in male and female plants of Populus tremula.
d – Open bracteous raceme.
1 – Bud scales; 2 – cataphylls; 3 – vegetative zone; 4 – flowers; 5 – bicarinate bud scale; 6 – developed leaves; 7 – open bracteous raceme.
Скелетные оси у Fraxinus excelsior могут длительное время нарастать моноподиально. Появление односезонных генеративных побегов из закладывающихся на этих осях пазушных почек не влияет на характер их нарастания.
Генеративные побеги Salix alba и S. pentandra
У мужских и женских растений Salix alba генеративные побеги во время цветения длиной 5–8 см имеют стебель, двукилевую почечную чешую в виде колпачка, катафиллы, небольшие листья (2–5) и завершаются соцветием кисть, имеющей облик сережки (рис. 11b, 11d). После цветения генеративные побеги мужских растений полностью отмирают. У женских растений на генеративных побегах после плодоношения отмирает только соцветие, а вегетативная зона сохраняется до конца вегетационного периода. Такие женские генеративные побеги называют двуэтапноопадающими (Skvortsov, 1965; Nedoseko, 2018). Нижняя олиственная часть двуэтапноопадающих женских генеративных побегов фотосинтезирует до осени.
Генеративные побеги S. pentandra имеют в вегетативной зоне, помимо двукилевой почечной чешуи и катафиллов, до 5–8 листьев срединной формации. Мужские генеративные побеги отмирают сразу после цветения, а у женских генеративных побегов вегетативная зона может сохраняться до весны следующего года – условно-неопадающие генеративные побеги регулярного возобновления (Nedoseko, 2018).
Мощные скелетные оси у Salix alba, S. pentandra нарастают акросимподиально. Появление генеративных побегов не сказывается на способе их нарастания.
Генеративные побеги Populus tremula
Генеративные побеги состоят из стебля, имеют почечные чешуи и завершаются сережковидными кистями. Листья срединной формации в вегетативной зоне генеративных побегов не развиваются (рис. 11c, 11d). Генеративные побеги после цветения (мужские) и после плодоношения (женские) полностью опадают.
Генеративные побеги Ulmus laevis и U. glabra
Генеративные побеги Ulmus laevis отличаются разнообразием, и их можно свести к трем основным вариантам (Grudzinskaya, 1966; Valikhodjaeva, 1976; Kostina, 2006).
1. Генеративные побеги длиной 3–7 мм имеют стебель, почечные чешуи и многочленные дихазии, которые образуются в пазухах верхних почечных чешуй. Общее число цветков на генеративном побеге колеблется от 20 до 100 и в среднем равно 30 (рис. 12а). После завершения роста побега его верхушка абортируется, как и во всех последующих вариантах генеративных побегов этого вида. После плодоношения генеративные побеги отмирают полностью. Такие генеративные побеги можно охарактеризовать как односезонные, не имеющие листьев срединной формации и завершающиеся открытым брактеозным тирсом.
Рис. 12.
Генеративные побеги Ulmus laevis (а–с), U. glabra (d).
1 – Почечная чешуя; 2 – спящие почки; 3 – развитый лист; 4 – почка регулярного возобновления; 5 – гибель верхушечной меристемы (завершение моноподиального нарастания); 6 – цветок; 7 – дихазий.
Fig. 12. Generative shoots of Ulmus laevis (а–с), U. glabra (d).
1 – Bud scale; 2 – dormant buds; 3 – developed leaf; 4 – bud of regular renewal; 5 – apical meristem cell death (monopodial growth termination); 6 – flower; 7 – dichasium.

2. Генеративные побеги длиной 7–25 см. В их основании в пазухах нижних почечных чешуй формируются спящие почки, а в пазухах верхних почечных чешуй и нижних листьев срединной формации образуются многочленные дихазии. В пазухах листьев, расположенных выше, формируются почки регулярного возобновления. После плодоношения отмирают только пазушные соцветия, а вся ось генеративного побега входит в состав многолетней осевой системы растения (рис. 12b). Такие генеративные побеги можно охарактеризовать как олиственные, с интеркалярным брактеозным тирсом, расположенным в проксимальной части генеративного побега.
3. Генеративные побеги достигают в длину 5–7 см. Выше зоны пазушных соцветий развиваются несколько небольших листьев срединной формации. После цветения генеративные побеги обычно отмирают полностью (рис. 12с).
Большая часть генеративных побегов, образующаяся у Ulmus laevis, относится к первому варианту. Генеративные почки, из которых формируются такие побеги, закладываются в основании длинных вегетативных побегов, в основании и средней части побегов средней длины или по всей длине коротких. Олиственные генеративные побеги с интеркалярным соцветием приурочены к верхней части кроны, где ростовые процессы протекают наиболее интенсивно.
У U. glabra генеративные побеги имеют стебель, почечные чешуи. В пазухах верхних почечных чешуй формируются цимы, состоящие из 1–3 цветков. Длина осевой части таких побегов не превышает 5 мм. Всю совокупность пазушных цветоносов можно описать как открытый компактный брактеозный тирс, состоящий из 15–30 цветков (рис. 12d).
Генеративные побеги Picea abies
Генеративные почки с мужскими стробилами обычно развиваются из пазушных, реже из верхушечных почек, закладывающихся на вегетативных побегах (рис. 13a). В состав мужских генеративных побегов длиной до 1 см входят стебель, почечные чешуи и терминальный мужской стробил (рис. 13b). Почки с женскими шишками могут образовываться как из пазушных, так и из верхушечных почек (рис. 13c). Как правило, на одном материнском побеге формируются или мужские генеративные почки, или женские. Реже можно наблюдать образование и тех и других. Женские генеративные побеги длиной до 1.5 см с молодыми шишками имеют стебель, почечные чешуи, несколько листьев срединной формации и завершаются терминальной женской шишкой (рис. 13d). Мужские генеративные побеги отмирают после пыления, женские – после высыпания семян.
Рис. 13.
Формирование двулетней побеговой системы Picea abies, в образовании которой принимают участие генеративные побеги.
а – Строение вегетативного побега с вегетативными почками и почками с мужскими шишками.
b – Строение двулетней побеговой системы.
с – Строение вегетативного побега с вегетативными почками и почками с женскими шишками.
d – Строение двулетней побеговой системы.
1 – Почечные чешуи; 2 – спящие почки; 3 – развитый лист (хвоинка); 4 – генеративная почка с мужской шишкой; 5 – вегетативная пазушная почка; 6 – вегетативная верхушечная почка; 7 – генеративный побег с мужской шишкой; 8 – мужской генеративный побег с мужской шишкой; 9 – вегетативный побег из пазушной почки; 10 – вегетативный побег из верхушечной почки; 11 – пазушная почка с женской шишкой; 12 – терминальная почка с женской шишкой; 13 – женская шишка; 14 – генеративный побег с женской шишкой.
Fig. 13. Biennial shoot system formation in Picea abies, involving generative shoots.
а – Structure of vegetative shoot bearing vegetative buds and buds with male cones.
b – Structure of biennial shoot system.
с – Structure of vegetative shoot bearing vegetative buds and buds with female cones.
d – Structure of biennial shoot system.
1 – Bud scales; 2 – dormant buds; 3 – developed leaf (needle); 4 – generative bud with male cone; 5 – vegetative axillary bud; 6 – vegetative apical bud; 7 – generative shoot with male cone; 8 – male generative shoot with male cone; 9 – vegetative shoot developed from the axillary bud; 10 – vegetative shoot developed from the apical bud; 11 – axillary bud with female cone; 12 – terminal bud with female cone; 13 – female cone; 14 – generative shoot with female cone.
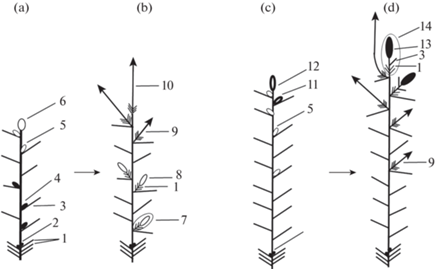
Мощные скелетные оси I и II порядков у P. abies на первых этапах развития формируются на основе вегетативных побегов. Далее в их образовании начинают участвовать вегетативные побеги, на которых помимо вегетативных закладываются генеративные почки с женскими шишками. Последние занимают пазушное положение и развиваются в средней части побега. Такой характер расположения генеративных почек позволяет вегетативным побегам сохранять способность к моноподиальному нарастанию, так как они завершаются верхушечными вегетативными почками. На заключительных этапах формирования скелетных осей в их образовании принимают участие вегетативные побеги с верхушечными генеративными почками. Образование генеративных побегов с женской шишкой из верхушечных почек приводит к прекращению нарастания скелетной оси или к смене способа нарастания оси с моноподиального на симподиальное (рис. 13с, 13d).
Вегетативные побеги с мужскими генеративными почками обычно не принимают участие в образовании мощных скелетных осей I и II порядков. Мужские генеративные почки обычно приурочены к осям III и последующих порядков. Появление из верхушечных почек генеративных побегов с мужскими стробилами обычно ведет к прекращению нарастания скелетных осей.
Расположение мужских и женских генеративных почек в побеговой системе дерева P. abies подчиняется определенным закономерностям (Anikeeva, Minina, 1959): генеративные почки с женскими шишками обычно формируются на ростовых побегах, а генеративные почки с мужскими стробилами – на побегах коротких и средней длины.
ОБСУЖДЕНИЕ
Результаты проведенного исследования позволили выявить следующие основные типы генеративных побегов, характерных для деревьев, произрастающих в средней полосе европейской части России.
1. Односезонные генеративные побеги.
У семи из рассмотренных нами видов (Populus tremula, Salix alba, S. pentandra, Fraxinus excelsior, Ulmus glabra, U. laevis, Picea abies) формируются односезонные генеративные побеги. У первых шести видов генеративные побеги развиваются только из пазушных почек, у Picea abies – как из пазушных, так и из верхушечных.
Генеративные побеги Populus tremula, Fraxinus excelsior, Ulmus glabra, мужские генеративные побеги Picea abies не имеют листьев срединной формации, а у Salix alba и S. pentandra они развиваются. У Quercus robur также формируются мужские односезонные генеративные побеги, обычно не имеющие листьев срединной формации и образующиеся из пазушных почек.
2. Олиственные генеративные побеги с интеркалярным расположением соцветий или мужских стробилов и женских шишек и полностью сохраняющейся осью. Пазушные соцветия или шишки/стробилы образуют генеративные зоны, расположенные между вегетативными зонами генеративного побега.
У Quercus robur и Pinus sylvestris оси генеративных побегов сохраняются, завершаются верхушечными зимующими вегетативными почками, пазушные соцветия и шишки / стробилы полностью отмирают после пыления или образования плодов или семян. У Tilia cordata верхушечная почка в конце вегетации отмирает, и сохраняется вся ось генеративного побега и проксимальные участки осей пазушных соцветий с почками регулярного возобновления.
У Ulmus laevis наряду с односезонными генеративными побегами изредка формируются генеративные побеги с сохраняющейся осью, отмирающей в конце вегетационного периода верхушечной почкой и интеркалярным расположением соцветий.
3. Олиственные генеративные побеги с терминальными соцветиями и сохраняющей осевой частью вегетативной зоны.
У Acer platanoides, Alnus glutinosa и Betula pendula, Betula pubescens соцветия в системе генеративного побега занимают терминальное положение. В вегетативной зоне формируются листья срединной формации, в пазухах которых закладываются вегетативные и генеративные почки или формируются вегетативные силлептические побеги. После цветения и плодоношения отмирает только соцветие, а осевая часть вегетативной зоны с почками регулярного возобновления входит в состав многолетней осевой системы растения.
Участие генеративных побегов в построении многолетней осевой системы растения
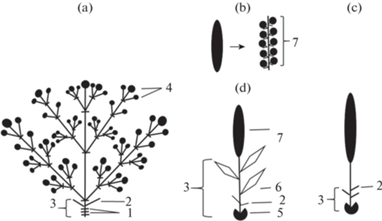
По степени участия генеративных побегов в построении многолетней осевой системы их можно разделить на три группы. К первой группе относятся односезонные генеративные побеги, формирующиеся у Populus tremula, Salix alba, S. pentandra, Fraxinus excelsior, Ulmus glabra, U. laevis (большая часть), Picea abies, Quercus robur (с мужскими сережками), которые не принимают участие в формировании многолетней осевой системы растения и не влияют на характер нарастания скелетных осей (рис. 14a), за исключением Picea abies. У последнего вида генеративные побеги с женскими шишками, появляющиеся на заключительных этапах формирования скелетных осей из верхушечных почек, приводят к смене моноподиального нарастания на симподиальное.
Рис. 14.
Строение трехлетних побеговых систем с разными вариантами генеративных побегов.
a – Трехлетняя побеговая система с односезонными генеративными побегами.
b – Трехлетняя побеговая система с генеративными побегами с пазушными соцветиями или стробилами.
c – Трехлетняя побеговая система с генеративными побегами, имеющими терминальные соцветия и многолетний осевой участок.
1 – Почечное кольцо; 2 – пазушные вегетативные побеги; 3 – отмершее соцветие; 4 – генеративный побег прошлого года; 5 – соцветие текущего года; 6 – генеративный побег текущего года; 7 – вегетативный побег, принимающие участие в формировании скелетной оси; 8 – продолжение моноподиального нарастания.
Fig. 14. Structure of triennial shoot systems with different kinds of generative shoots.
a – Triennial shoot system with single-season generative shoots.
b – Triennial shoot system with generative shoots bearing axillary inflorescences or cones.
c – Triennial shoot system with generative shoots bearing terminal inflorescences and perennial section of the axis.
1 – Bud Scar; 2 – axillary vegetative shoots; 3 – dead inflorescence; 4 – generative shoot of last year; 5 – inflorescence of current year; 6 – generative shoot of current year; 7 – vegetative shoot involved in the skeletal axis formation; 8 – continuing monopodial growth.

У видов, односезонные генеративные побеги которых развиваются только из пазушных почек, существует определенная закономерность в размещении вегетативных и генеративных почек на длинных и коротких материнских вегетативных побегах. На длинных ростовых побегах в верхней их части формируются вегетативные почки, а в средней и нижней частях – генеративные. Такой характер расположения вегетативных и генеративных почек позволяет ростовым вегетативным побегам формировать крону за счет активного нарастания и акротонно ветвиться. На коротких побегах генеративные почки занимают большую часть побега.
У всех остальных рассмотренных нами видов (вторая и третья группы) после цветения и плодоношения остается резид, который встраивается в многолетнюю осевую систему растения.
Во второй группе у видов с пазушными соцветиями (Quercus robur с олиственными побегами (рис. 1а, 1b, 1с), Tilia cordata и Pinus sylvestris) появление генеративных побегов не вызывает перестройку работы верхушечных меристем скелетных осей (рис. 14).
В третьей группе с верхушечными и пазушными соцветиями у Acer platanoides, Alnus glutinosa, A. incana и Betula pendula, B. pubescens образование генеративных побегов, напротив, приводит к появлению побеговых систем, изменяющих габитус дерева. У первых трех видов появление генеративных побегов вызывает смену моноподиального нарастания на симподиальное (рис. 14с). Генеративные побеги этих видов участвуют в формировании скелетных осей I и II порядков, но только на заключительных стадиях их развития. Это обусловлено тем, что на первых этапах формирования ствола и мощных ветвей длина вегетативных побегов значительно превышает длину генеративных побегов. Так, например, у Betula pendula длина вегетативных побегов, из которых строится ствол, колеблется от 60 до 150 см. По мере развития скелетных осей длина вегетативных побегов постепенно уменьшается, и когда размеры этих побегов становятся сопоставимыми с размерами генеративных побегов, последние также начинают участвовать в образовании осей.
Следует подчеркнуть, что в первых двух группах пазушные соцветия и односезонные генеративные побеги не являются гомологичными структурами. Односезонные генеративные побеги функционально ведут себя как пазушные соцветия: и те и другие отмирают после плодоношения (за исключением Tilia cordata). В то же время односезонные генеративные побеги представляют собой самостоятельные конструктивные элементы, имеющие собственные почечные чешуи. Повреждение одной генеративной почки не влияет на состояние других. Пазушные соцветия являются элементами генеративного побега и на внутрипочечном этапе развития защищены общими почечными покровами этого побега. Повреждение почки приводит к отмиранию всех пазушных соцветий.
Особенности строения генеративных побегов, определяющие сроки цветения
Рассмотренные нами виды деревьев, за исключением Tilia cordata, цветут весной до или одновременно с распусканием листьев (Serebryakov, 1949, 1952). Одним из факторов, определяющих сроки цветения, является степень сформированности генеративного побега в почке (Serebryakov, 1949, 1952; Bulygin, 1964; Schnablova et al., 2021).
По степени сформированности генеративного побега будущего года в почках регулярного возобновления в конце вегетационного периода все наши древесные и травянистые растения были подразделены И.Г. Серебряковым (Serebryakov, 1949, 1952) на три группы. К первой группе он отнес растения, у которых в почках регулярного возобновления побег будущего года сформирован полностью, включая соцветия, т.е. генеративный побег полностью преформирован. Ко второй группе были отнесены виды, у которых в почках регулярного возобновления сформирована только вегетативная часть генеративного побега, а соцветие развивается в текущем году, в процессе внепочечного роста генеративного побега. В третью группу объединены растения, у которых в почках регулярного возобновления успевает сформироваться только часть вегетативной сферы генеративного побега. Следует отметить, что при выделении этих групп И.Г. Серебряков не принимал во внимание взаимное расположение соцветий и вегетативной зоны у генеративного побега.
Проведенное нами исследование показало, что ранние сроки цветения обусловлены тремя структурно-ритмологическими вариантами генеративных побегов.
1. К первому варианту относятся семь видов с односезонными генеративными побегами, как с листьями срединной формации, так и без них, и один вид с терминальным соцветием и небольшим числом листьев срединной формации в вегетативной зоне. Генеративные побеги у представителей этого варианта имеют небольшие размеры – от 1 см (у Ulmus glabra) до 19 см (у Acer platanoides) и преформированы в зимующих почках (Serebryakov, 1952; Bulygin, 1964). Все эти виды относятся к первой группе, выделенной И.Г. Серебряковым.
2. Ко второму варианту относятся виды с генеративными побегами, имеющими интеркалярные соцветия, стробилы или шишки – Quercus robur и Pinus sylvestris. Длина генеративных побегов у первого вида может достигать 30 см, а у второго – до 50 см. У представителей этого варианта степень сформированности мужских соцветий или стробилов в зимующих почках намного превосходит степень сформированности женских соцветий или шишек. Это определяется разным положением мужских и женских соцветий, а также стробилов и шишек на оси генеративного побега. Благодаря тому, что мужские соцветия или стробилы находятся в проксимальной части генеративного побега, к началу цветения (пыления) в их цветках или микроспорангиях уже сформирована полноценная пыльца, опыляющая женские цветки или шишки. Женские же цветки или семязчатки, образующиеся в дистальной части генеративного побега, к моменту опыления не способны к оплодотворению. По классификации И.Г. Серебрякова, эти виды попадают в первую выделенную им группу.
Следует отметить, что в отличие от представителей первого варианта, у которых сроки цветения определяются числом листьев срединной формации под терминальным соцветием, сроки цветения генеративных побегов с интеркалярным расположением не зависят от длины генеративных побегов и числа листьев, расположенных выше генеративной зоны.
3. Виды с генеративными побегами, у которых соцветия зимуют на стадии предсоцветия. В зимующих почках Alnus glutinosa сформирована только вегетативная зона, а мужские и женские сережки образуются в начале лета на внепочечной стадии побега. Их дальнейшее развитие тормозится на стадии предсоцветия. У берез такой структурно-ритмологический вариант строения имеют только генеративные побеги с мужскими сережками.
И.Г. Серебряков отнес Alnus glutinosa, Betula pendula, B. pubescens к первой группе. Однако поскольку в почках у этих видов преформирована только вегетативная сфера, а мужские соцветия у берез и мужские и женские соцветия у ольхи развиваются в текущем году, то их следовало бы отнести ко второй группе. Но у представителей второй группы И.Г. Серебрякова соцветия переходят к цветению в год образования, а у Alnus glutinosa, Betula pendula, B. pubescens остаются на стадии предсоцветия и цветут только весной следующего года. Поэтому представителей семейства березовые нельзя отнести ни к одной из групп, выделенных И.Г. Серебряковым.
Как и у Quercus robur, у Alnus glutinosa, A. incana, Betula pendula, B. pubescens цветки в мужских соцветиях в большей степени дифференцированы, чем в женских. Но если у первого вида это достигается различием в положении соцветий на оси генеративного побега, то у Alnus glutinosa и A. incana – более поздней закладкой женских соцветий, а у берез – нахождением женских соцветий на верхушках олиственных побегов в зимующих почках.
Позднецветущий вид Tilia cordata И.Г. Серебряков отнес ко второй группе. Однако, исходя из строения генеративных побегов невозможно объяснить, почему в зимующих почках T. cordata отсутствуют зачатки соцветий, хотя соцветия на генеративных побегах располагаются в пазухах листьев срединной формации, начиная со 2–3-го. По строению вегетативной сферы этот вид должен попасть в первую группу, выделенную И.Г. Серебряковым, потому что во вторую группу отнесены виды, у которых до соцветия развивается более пяти листьев. Поздние сроки цветения T. cordata определяются не особенностями строения вегетативной сферы, а ритмом развития пазушных цветоносов, которое можно охарактеризовать как промежуточное между пролептическим и силлептическим. На пролептический характер ветвления указывает образование укороченного гипоподия, двух чешуевидных листьев и почки регулярного возобновления, а также неполное отмирание пазушного цветоноса после плодоношения. Возможно, что задержка заложения и цветения пазушных соцветий у T. cordata стала результатом расхождения экологических ниш в конкуренции за опылителей с рано цветущими травами широколиственных и смешанных лесов.
Вопрос о том, трактовать ли пазушные цветоносы липы как самостоятельные генеративные побеги, либо как силлептические побеги в составе разветвленного генеративного побега, остается открытым. А.А. Чистякова (Chistyakova, 1979) рассматривает пазушные цветоносы T. cordata как самостоятельные генеративные побеги. Мы относим липу к видам, имеющим интеркалярное цветорасположение.
Анализ строения генеративных побегов и сроков цветения показал, что у большей части деревьев первой величины существуют сходные структурно-ритмологические адаптации, позволяющие им реализовать возможность раннего цветения, а у целого ряда видов – и ветроопыления. У меньшей части видов ранние сроки цветения обусловлены задержкой развития соцветий на стадии предсоцветия. Поздние сроки цветения T. cordata, не вписывающиеся в общую картину феноритмики деревьев умеренной зоны, не влияют на успешность существования этого вида в сезонном климате. Это еще раз подтверждает множественность путей адаптаций растений, принадлежащих одной жизненной форме и произрастающих в одном климате, среди которых, наряду с весьма распространенными, есть и редко встречающиеся, но не менее эффективные.
Генеративные побеги с интеркалярным расположением соцветий
Результаты проведенного исследования показали, что у Ulmus laevis и Quercus robur на одном растении наряду с генеративными побегами, имеющими пазушные соцветия и сохраняющуюся ось генеративного побега, развиваются односезонные генеративные побеги, не имеющие листьев срединной формации. У Q. robur образование таких побегов происходит путем недоразвития вегетативной зоны, расположенной выше генеративной зоны с пазушными мужскими соцветиями. Благодаря появлению односезонных побегов у этого вида значительно увеличивается число мужских цветков, а следовательно, и пыльцы. У U. laevis преобладают односезонные генеративные побеги. Их образование, так же, как и у Q. robur, связано с завершением морфогенеза генеративного побега после формирования зоны с пазушными соцветиями. Расположение соцветий у этих видов в основании генеративных побегов обуславливает ранние сроки цветения.
Генеративные побеги с терминальным расположением соцветий
У видов родов Acer и Fraxinus по характеру и числу листьев, развивающихся в вегетативной зоне, и по числу цветков в соцветии генеративные побеги можно расположить в сравнительно-морфологический ряд. Крайними членами этого ряда, с одной стороны, будут генеративные побеги, у которых в вегетативной зоне развиваются ассимилирующие листья и почки регулярного возобновления. С другого края этого ряда располагаются односезонные, крайне специализированные генеративные побеги, имеющие ранние сроки цветения.
Например, в роде Fraxinus самые ранние сроки цветения в коллекции ГБС имеют виды, у которых формируются крайне специализированные односезонные генеративные побеги. Так, Fraxinus pennsylvanica Marsh. и F. excelsior цветут в начале мая, до развертывания листьев. Позже переходит к цветению F. rhynchophylla Hance, в вегетативной зоне генеративных побегов которого располагаются листья срединной формации (Drevesny’e…, 2005).
В роде Acer ранними сроками цветения также обладают виды с крайне специализированными односезонными генеративными побегами. В коллекции ГБС к таким видам относятся Acer rubra L. и широко распространенный в нашем регионе чужеродный североамериканский двудомный вид A. negundo L. Причем деревья с женскими генеративными побегами, в вегетативной зоне которых развивается пара ассимилирующих листьев, цветут позже мужских растений, для которых характерны крайне специализированные генеративные побеги (Mikhalevskaya, 2001; Kiseleva, Shorinа, 2003; Kostina, 2005, 2009). Возникновение таких специализированных генеративных побегов связано с полной редукцией листьев срединной формации и почек возобновления в вегетативной зоне.
Рассмотренные выше сравнительно-морфологические ряды генеративных побегов можно трактовать как в сторону увеличения, так и в сторону уменьшения количественных параметров (листьев срединной формации, почек). Однако направление преобразования генеративных побегов у деревьев умеренного климата определяется спецификой организации данной жизненной формы, адаптированной к существованию в условиях короткого вегетационного периода и продолжительного зимнего периода покоя. Для деревьев с их медленной сменой основных скелетных осей основной стратегической задачей является сохранение многолетней осевой системы в зимний период. Поэтому смещение всех процессов, связанных с ростом и цветением, на весну или начало лета позволяет представителям данной жизненной формы заранее подготовиться к зимнему периоду. Можно предположить, что морфогенетические перестройки генеративных побегов, направленные в сторону односезонных рано цветущих генеративных побегов, появление которых в побеговом теле растений не отражается на характере нарастания осей, будут преобладающими.
2. Модусы, затрагивающие ритм развития соцветий
Структурно-ритмологические преобразования могут затрагивать не только вегетативную зону генеративного побега, но и изменять ритм развития соцветия. Поздние сроки цветения Tilia cordata, по всей вероятности, определяются некоторым смещением силлептического ветвления в сторону пролептического.
Еще один вариант ритмологических перестроек можно продемонстрировать на примере Alnus glutinosa, A. incana, Betula pendula и B. pubescens. У первых двух видов мужские и женские сережки зимуют на стадии предсоцветия, а у B. pendula и B. pubescens на стадии предсоцветия в зимний период находятся только мужские сережки, а женские сережки зимуют в почках. Образование предсоцветий у Alnus glutinosa, A. incana, Betula pendula и B. pubescens обусловлено замедлением темпов развития соцветий, которые, минуя внутрипочечный этап развития, переходят к бутонизации и цветению только в следующем году рано весной.
Еще более выраженная задержка формирования боковых осей, несущих женские соцветия, у B. pendula и B. pubescens приводит к тому, что в зимний период они находятся в почках и защищены почечными чешуями, приобретая, таким образом, статус самостоятельных генеративных побегов.
Особенности расположения генеративных побегов в кроне деревьев умеренной зоны
У деревьев умеренно-континентального климата рост большинства побегов происходит непродолжительное время (Serebryakov, 1952). Большую часть вегетационного периода растения тратят на подготовку к зимнему покою, поскольку, в отличие от травянистых растений, им необходимо сохранить свою надземную осевую систему и, прежде всего, ствол и отходящие от него крупные ветви. Генеративные побеги с компактными соцветиями имеют, как правило, небольшие размеры и приурочены к осям высоких порядков. Таких побегов в кроне формируется много. Характер их отмирания позволяет полнее сохранить осевую систему растения, увеличивая при этом степень дифференциации ее осей на скелетные и заполняющие.
У многих тропических деревьев формирование генеративных побегов с крупными соцветиями нередко происходит на осях низших порядков. Отмирание таких соцветий вызывает отторжение значительной части побеговой системы растения, а у видов с верхушечными соцветиями изменяет характер дальнейшего нарастания ствола и мощных ветвей.
Особенности строения генеративных побегов, их ритма развития и расположения в кроне, присущие деревьям умеренного климата, позволяют всем процессам, связанным с ростом и цветением, сместиться на весну или начало лета, что оказывается биологически выгодным. Особенно это касается ветроопыляемых видов, которые цветут до распускания листьев.
Для деревьев умеренного пояса авторы концепции архитектурных моделей указывают модели Rauh и Troll, у которых появление компактных пазушных соцветий не приводит к изменению способа нарастания скелетных осей (Hallé, Oldeman, 1970; Hallé, Oldeman, Tomlinson, 1978). В целом правильно понимая этот аспект организации деревьев умеренного климата, зарубежные коллеги не учитывают все разнообразие вариантов “встраивания” соцветий в побеговую систему деревьев умеренного пояса, не рассматривают специфику односезонных генеративных побегов с терминальными и пазушными соцветиями, предсоцветий.
ВЫВОДЫ
1. Для деревьев средней полосы европейской части России характерны односезонные генеративные побеги, генеративные побеги с интеркалярным расположением соцветий, шишек или стробилов и полностью сохраняющейся осью, генеративные побеги с терминальными соцветиями и сохраняющейся осевой частью вегетативной зоны. Преобладают виды с односезонными генеративными побегами.
2. Ранние сроки цветения определяются отсутствием или небольшим числом листьев срединной формации, развивающихся под терминальным соцветием, расположением пазушных соцветий или стробилов в проксимальной части генеративных побегов с интеркалярным цветорасположением и формированием генеративных побегов с терминальными соцветиями, зимующими на стадии предсоцветия.
3. Односезонные генеративные побеги могут образоваться в результате недоразвития в вегетативной зоне почек регулярного возобновления как на основе генеративных побегов с интеркалярным, так и с терминальным цветорасположением. Изменение ритма формирования терминальных или пазушных соцветий может привести к смещению сроков цветения как на раннюю весну, так и на летний период.
4. Появление в кроне односезонных генеративных побегов не влияет на характер нарастания скелетных осей, так же, как и появление генеративных побегов с интеркалярным расположением соцветий. Образование генеративных побегов с терминальными соцветиями и сохраняющейся осевой вегетативной частью приводит к смене моноподиального нарастания на симподиальное. Генеративные побеги этого типа могут участвовать в образовании мощных скелетных осей, но только на заключительных этапах их формирования. Особенности строения генеративных побегов, их расположения в кроне и смещение сроков цветения на весну-начало лета способствует сохранению многолетней осевой системы растения.
5. Использование при анализе побегового тела растений в качестве конструктивных элементов не только вегетативных, но и генеративных побегов позволяет выявить бóльшее разнообразие вариантов цветорасположения, чем предполагалось концепцией архитектурных моделей для деревьев умеренного климата, а также глубже понять специфику конструктивной организации деревьев, дающей возможность растениям соответствовать годовому климатическому ритму.
Список литературы
[Anikeeva, Minina] Аникеева И.Д., Минина И.Д. 1959. О жизнедеятельности конуса нарастания у древесных пород в связи с сексуализацией побегов. – Бот. журн. 44(7): 907–915.
Austen E.J., Rowe L, Stinchcombe J.R., Forrest J.R.K. 2017. Explaining the apparent paradox of persistent selection for early flowering. New Phytologist. 215: 929–934.
Barthelemy D., Caraglio Y. 2007. Plant architecture: A dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. – Ann. Bot. 99: 375–407.
[Bulygin] Булыгин Н.Е. 1964. Периоды образования цветков в почках древесных растений и влияние погоды на обилие их цветения в Ленинграде. – Труды Всесоюзного заочного лесотехнического ин-та. 8: 193–223.
[Chistyakova] Чистякова А.А. 1978. О жизненной форме и вегетативном разрастании липы сердцевидной. – Бюл. МОИП. Отд. биол. 83(2): 129–137.
[Drevesny’e…] Древесные растения главного ботанического сада им. Н.В. Цицина РАН: 60 лет интродукции. 2005. М. 586 с.
Duffy N.M, Bonser S.P., Aarssen L.W. 1999. Patterns of variation in meristem allocation across genotypes and species in monocarpic Brassicaceae. Oikos 84: 284.
Ehrlen J. 2015. Selection on flowering time in a life-cycle context. Oikos 124: 92–101.
[Filatovа et al.] Филатова О.В., Перцев Е.В., Шатровская В.И. 1989. Корреляция развития почек и сроков цветения деревьев и кустарников. – Бюл. Гл. Бот. сада. 153: 9–14.
Geber M.A. 1990. The cost of meristem limitation in Polygonum arenastrum: negative genetic correlations between fecundity and growth. Evolution. 44: 799.
[Grudzinskaya] Грудзинская И.А. 1960. Летнее побегообразование у древесных растений и его классификация. – Бот. журн. 45(7): 968–978.
[Grudzinskaya] Грудзинская И.А. 1966. Соцветия видов Ulmus L. (Формирование, строение и некоторые вопросы эволюции). – Бот. журн. 51(1): 15–28.
Hallé F., Oldeman R.A.A. Essai sur l’architecture et la dynamique de croissance des arbres tropicaux. 1970. Paris. 178 p.
Hallé F., Oldeman R.A.A., Tomlinson P.B. Tropical trees and forests. An architectural analysis. 1978. Berlin, New York. 441 p.
[Ims] Имс. А. Морфология цветковых растений. 1964. М. 497 с.
Jäger E.J. 1980. Progressionen im Synfloreszenzbau und in der Verbreitung bei den Betulaceae. – Flora. 170: 91–113.
[Kiseleva] Киселева Л.Г. 2003. О типах генеративных побегов клена остролистного (Acer platanoides L.). – Научные труды МПГУ. Серия: Естественные науки. С. 312–314.
[Kiseleva, Shorinа] Киселева Л.Г., Шорина Н.И. 2003. Сравнительный анализ биологии цветения трех видов клена. – Доклады ТСХА. 275: 48–53.
[Korchaginа] Корчагина И.А. 1980. Семейство березовые (Betulaceae). – В кн.: Жизнь растений. Т. 5. Ч. 1. М. С. 311–324.
[Kostina] Костина М.В. 2001. Взаимное расположение и ритм развития мужских и женских сережек у представителей семейства Betulaceae. – Бюл. Гл. Бот. сада. 182: 97–111.
[Kostina] Костина М.В. 2005. Строение и ритм развития генеративных побегов древесных растений в связи с продолжительностью цветения. – Бюл. Гл. Бот.сада. 189: 188–208.
[Kostina] Костина М.В. 2006. Строение и динамика развития генеративных побегов в роде Ulmus (Ulmaceae). – Бот. журн. 91(7): 1015–1028.
[Kostina] Костина М.В. 2008. Роль генеративных побегов в построении кроны у представителей рода Acer L. – Бюл. Гл. Бот. сада. 193: 87–106.
[Kostina] Костина М.В. 2009. Проявление полового диморфизма в строении кроны и генеративных побегов Acer negundo L. (Aceraceae) и Fraxinus pennsylvanica Marsh. (Oleaceae). – Вестник Тв. ГУ. Сер. “Биология и экология”. 11: 108–117.
[Kostina] Костина М.В. 2010. Значение признаков генеративных побегов в систематике и филогенетике родов Syringa L. и Fraxinus L. – В кн.: XII Московское совещание по филогении растений, посвященное 250-летию со дня рождения Георга-Франца Гофмана. М. С. 56–60.
[Kostina et al.] Костина М.В., Ясинская О.И., Битюгова Г.В. 2015. Интенсивность пыления березы повислой (Betula pendula Roth.) в зависимости от строения побегов с мужскими сережками. – Бюл. Моск. о-ва испытателей природы. Отд. Биол. 120(4): 79–85.
Kostina M.V., Barabanshikova N.S., Bityugova G.V., Yasinskaya O.I., Dubach A.M. 2015. Structural Modifications of Birch (Betula pendula Roth.) Crown in Relation to Environmental Conditions. – Contemporary Problems of Ecology. 8(5): 584–597.
[Kostina et al.] Костина М.В., Барабанщикова Н.С., Ясинская О.И. 2018. Теоретические и методические основы изучения крон деревьев умеренной зоны. – В кн.: материалы II Межд. конф. (к 90-летию со дня рождения проф. А.Г. Еленевского) “Систематические и флористические исследования Северной Евразии”. Т. 2. М. С. 58–61.
[Kozubov] Козубов Г.М. 1974. Биология плодоношения хвойных на севере. Л. 136 с.
Kudoh H., Kachi N., Kawano S. 2002. Intrinsic cost of delayed flowering in annual plants: negative correlation between flowering time and reproductive effort. – Plant Species Biology. 17: 101–107.
Kusnetzova T.A. 1988. Angiosperm inflorescences and different types of their structural organization. – Flora. 181(1): 1–17.
[Kuznetsova] Кузнецова Т.В. 1992. О комплементарных подходах в морфологии соцветий. – Бот. журн. 77(12): 7–24.
[Kuznecova, Timonin] Кузнецова, Т.В., Тимонин А.К. 2017. Соцветие: морфология, эволюция, таксономическое значение (применение комплементарных подходов. М. 183 с.
[Minina] Минина Е.Г. 1954. Биологические основы цветения и плодоношения дуба. – Тр. Ин-та леса АН СССР. 17: 5–97.
[Mikhalevskaya] Михалевская О.Б. 1962. Развитие почек сосны обыкновенной в условиях Московской области. – Бюл. ГБС. 48: 61–68.
[Mikhalevskaya] Михалевская О.Б. 2001. Структура и регуляция развития побеговых систем и элементарных побегов у некоторых видов Acer (Aceraceae). – Бот. журн. 86(10): 42–52.
[Mikhalevskaya] Михалевская О.Б. 2002. О внутри- и внепочечной фазах развития побеговых систем и элементарных побегов древесных растений. – Онтогенез. 33(4): 258–263.
[Mikhalevskaya et al.] Михалевская О.Б., Костина М.В., Скалепова Л.В. 2006. Структура и динамика развития вегетативных и генеративных побегов у некоторых представителей семейства Betulaceae. – Бюл. Гл. Бот. сада. 191: 140–149.
Millet J., Bouchard B. 2011. Architecture of silver maple and its response to pruning near the power distribution network. – Can. J. Bot. 33: 726–739.
Millet J., Bouchard B., Edelin C. 1998a. Plant succession and tree architecture: an attempt at reconciling two scales of analysis of vegetation dynamics. – Acta Biotheor. 46: 1–22.
Millet J., Bouchard B., Edelin C. 1998в. Plagiotropic architectural development of four tree species of the temperate forest. – Can. J. Bot. 76: 2100–2118.
Millet J., Bouchard B., Edelin C. 1999. Relationship between architecture and successional status of trees in the temperate deciduous forest. – Ecoscience. 6: 187–203.
Munguia-Rosas M.A., Ollerton J., Parra-Tabla V., De-Nova J.A. 2011. Metaanalysis of phenotypic selection on flowering phenology suggests that early flowering plants are favoured. – Ecology Letters. 14: 511–521.
[Nedoseko] Недосеко О.И. 2018. Становление жизненных форм и архитектоники крон бореальных видов ив подродов Salix и Vetrix Dumort. в онтогенезе: Автореф. дис. … докт. биол. наук. М. 43 с.
[Nukhimovsky] Нухимовский Е.Л. 1971. Осевая и побеговая система семенных растений. – Изв. ТСХА. 1: 54–66.
[Nukhimovsky] Нухимовский Е.Л. 1986. Особенности фенетической организации биоморф семенных растений. – Успехи современной биологии. 102.2(5): 289–306.
[Nukhimovsky] Нухимовский Е.Л. 1997. Основы биоморфологии семенных растений: Т. 1. Теория организации биоморф. М. 630 с.
[Paderovskaya] Падеровская М.И. 1953. Биологические особенности почек дуба. – Уч. зап. Московского гос. пед. ин-та им. В.И. Ленина. 73(2): 73–130.
Schnablova R., Huang L., Klimesova J., Smarda P., Herben T. 2021. Inflorescence preformation prior to winter: a surprisingly widespread strategy that drives phenology of temperate perennial herbs. – New Phytologist. 229: 620–630.
[Serebryakov] Серебряков И.Г. 1949. Структура и ритм в жизни цветковых растений. – Бюл. МОИП. Отд. биол. 54(1): 47–62.
[Serebryakov] Серебряков И.Г. 1952. Морфология вегетативных органов высших растений. М. 391 с.
[Serebryakov] Серебряков И.Г. 1962. Экологическая морфология растений. М. 378 с.
[Skvortsov] Скворцов А.К. 1965. Ивы средней России в зимнем состоянии. – Бюл. Моск. общ. испыт. природы. Отд. биол. 60(3): 115–127.
Späth H.L. 1912. Der Johannistrieb. Berlin. 91 s.
Stairs C.R. 1964. Microsporogenesis and embryogenesis in Quercus. – Bot. Gaz. 125(2): 115–121.
Tomlinson P.B., Gill A.M. 1973. Growth habits of tropical trees: some guiding principles. Tropical forest ecosystems in Africa and South America: a comparative review. a comparative review. Wash. (D. C.). P. 129–143.
Troll W. 1957. Praktische Einführung in die Pflanzenmorphologie. Die blühende Ppflanze. Teil II. Jena. 420 p.
Troll W. 1964. Die Infloreszenzen. Typologie und Stellung im Aufbau des Vegetationskörpers. Bd. 1. Jena. 615 s.
Troll W. 1969. Die Infloreszenzen. Typologie und Stellung im Aufbau des Vegetationskörpers. Bd. 2. Jena. 630 s.
[Valikhodjaeva] Валиходжаева С.П. 1979. Интродуцированные виды рода Ulmus L. в Ботаническом саду АН УзССР: Биологические особенности видов рода Ulmus L., интродуцированных в Ботанический сад АН УзССР г. Ташкента: Дис. … канд. биол. наук. Ташкент. 201 с.
Weberling F. 1981. Morphologie der Blüten und der Blütenstände. 1981. Ulmer, Stuttgart. 45 p. https://doi.org/10.18756/edn.35.45.1
Weberling F. 1982. Current problems of modern inflorescence morphology. – Australian Systematic Botany Society Newsletter. 30: 5–21.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал