

Ботанический журнал, 2022, T. 107, № 8, стр. 731-748
РОД DRABA (BRASSICACEAE) В МУРМАНСКОЙ ОБЛАСТИ
А. В. Разумовская 1, *, Э. М. Мачс 2, М. Н. Кожин 3
1 Институт проблем промышленной экологии Севера КНЦ РАН
184209 Мурманская область, Апатиты, мкр. Академгородок, 14А, Россия
2 Ботанический институт им. В.Л. Комарова РАН
197022 Санкт-Петербург, ул. Профессора Попова, 2, Россия
3 Полярно-альпийский ботанический сад-институт им. Н.А. Аврорина КНЦ РАН
184209 Мурманская область, Апатиты, мкр. Академгородок, 18А, Россия
* E-mail: anna-lynx@mail.ru
Поступила в редакцию 23.03.2022
После доработки 04.06.2022
Принята к публикации 14.06.2022
- EDN: PAWJXP
- DOI: 10.31857/S0006813622080105
Аннотация
На основе гербарных и литературных данных, а также наблюдений авторов проведен анализ и составлены карты географического распространения в Мурманской области 11 видов рода Draba L. Внутригеномный полиморфизм критических образцов Draba и степень родства отдельных таксонов определялись с помощью молекулярно-генетического анализа по Сенгеру и методом NGS. Приводятся сведения о находках 2 новых для области видов: Draba oxycarpa Sommerf. и Draba micropetala Hook., а также о Draba cinerea Adams, не упоминавшейся ранее в русскоязычных региональных публикациях. Описанный с островов и побережья Белого моря вид Draba insularis Pissjauk предложено рассматривать в составе Draba hirta L. в качестве формы, встречающейся в условиях благоприятного микроклимата и минерального питания. Виды Draba, обитающие в Мурманской области, по типам ареалов разделяются на “арктическую” и “гипоарктомонтанную” группы с разными адаптационными характеристиками в условиях современного климата. Для обеих групп, несмотря на различия во встречаемости видов, свойственен преимущественно приморский характер распространения. В континентальных районах они приурочены к палео-фьордам и краевым зонам морских трансгрессий и имеют компактный, рефугиальный характер произрастания, что согласуется с гипотезой их перигляциального и шельфового расселения в Арктике четвертичного периода. Выявленные популяции крупок арктической группы, вследствие своего реликтового характера, заслуживают охраны и дальнейшего изучения. Выявленные закономерности современного расселения видов позволяют сформулировать рекомендации по корректировке системы природоохранных мероприятий в Мурманской области.
Виды рода Draba редки в Мурманской области, находки многих из них исчисляются единицами. Со времени составления карт распространения видов и последних региональных флористических сводок (Pis’yaukova, 1956, Ramenskaya, 1983) обобщающая информация появлялась лишь для некоторых из них в рамках периодически издаваемых областных Красных Книг. В последующие десятилетия благодаря флористическим исследованиям в различных районах области накапливались новые сведения о географии и экологии популяций, происходили важные находки, не всегда публиковавшиеся, а иногда лишь пополнявшие гербарные коллекции. С развитием интернет-коммуникаций стали более открытыми данные скандинавских исследователей XIX – начала XX века, что заметно повысило объем информации по западной части области. По мере накопления литературных и полевых данных стала очевидной неравномерность распределения даже более обычных видов рода Draba в отдельных районах области, равно в северотаежных и в (горно-) тундровых условиях. Находки последних лет довершили картину. Появилась возможность систематизировать и анализировать разрозненные сведения, чему и посвящена эта статья.
МАТЕРИАЛЫ И МЕТОДЫ
Основная часть данных по распространению видов рода Draba в Мурманской области получена в процессе изучения гербарных образцов коллекций INEP, KPABG, KAND, LE, MW, PTZ, H, OULU, TUR, литературных источников, включая флористические списки отдельных регионов и открытых интернет-ресурсов, в том числе, фондов гербариев Н, OULU, TUR (FinBIF…). Сбор информации о присутствии видов на побережьях и в континентальных районах Мурманской области проводился также в экспедициях 2011–2021 гг. С 2018 г. ведутся наблюдения по состоянию популяций редких и уникальных видов рода Draba на полуостровах Рыбачий и Средний (мониторинг численности и ареалов популяций, подтверждение и картирование находок начала XX века и др.). Карты распространения Draba в Мурманской области построены с использованием геоинформационной системы ArcGIS ArcMap 10.2 ESRI.
Молекулярно-генетические исследования
Для выяснения видовой принадлежности критических образцов Draba, а также установления степени родства Draba insularis и Draba hirta применялся молекулярно-генетический анализ по Сенгеру. Секвенирование было выполнено на генетическом анализаторе AbiPrism 3130 c использованием образцов тотальной ДНК (Doyle, Doyle, 1987) и праймеров ITS-1p (Ridgway et al., 2003) и ITS-4 (White et al., 1990) на том же амплификаторе, с теми же реактивами и по такому же протоколу. По результатам анализа выполнено построение филогенетического дерева методом максимального правдоподобия в программе Mega 7 (Kumar et al., 2018). Для его построения были привлечены последовательности представителей исследуемых и близких к ним таксонов, взятые из базы данных NCBI (National Center for Biotechno-logy Information) GenBank (Benson et al., 2005).
Выяснение возможных родительских видов для предположительно гибридных особей Draba oxycarpa проводилось методом NGS. На площадке наблюдений в месте их обнаружения (п-ов Средний) были взяты образцы совместно произрастающих явно гибридных и “фенотипически чистых” особей Draba oxycarpa, D. lanceolata и D. incana. Кроме того, контрольные образцы D. incana отобраны в изолированном местонахождении на побережье Кутовой губы в 12.5 км от площадки. Внутригеномный полиморфизм был изучен на основе локус-специфичного секвенирования NGS на платформе Illumina MiSeq. Подготовка библиотек проводилась с использованием образцов тотальной ДНК (Doyle, Doyle, 1987) и праймеров ITS-1p (Ridgway et al., 2003) и ITS-2 (White et al., 1990). Амплификация фрагмента проводилась на амплификаторе BioRad T-100 с использованием набора Plant PCR Master Mix Thermo Scientific по следующему протоколу: начальная денатурация 98°С 5 минут; 30 циклов амплификации (98°С 5 секунд, 56°С 5 секунд, 72°С 15 секунд); финальный синтез 72°С 1 минута; хранение 12°С. Обработка данных проводилась с использованием программ FastQC (Babraham Bioinformatics), Trimmomatic (Bolger et al., 2014), Fastq-join (Aronesty, 2013) и Vsearch (Rognes et al., 2016). Фильтрация полученных риботипов OTU проводилась вручную по выравниваниям с использованием программы Mega 7 (Kumar et al., 2018) и сервиса BLAST NCBI.
РЕЗУЛЬТАТЫ
Распространение видов Draba L. в Мурманской области
Draba alpina L. – Крупка альпийская. Редкий в Мурманской области вид, занесен в Красную книгу Мурманской области (далее ККМО) с категорией 3 (Red…, 2014). Известны всего несколько его местонахождений (рис. 1a), преимущественно на Баренцевоморском побережье: на крайнем северо-западе в Расватунтури (Печенгский р-н) – сборы П. Каллио, 1941, TUR (FinBIF…); в районе Святого Носа (Восточный Мурман, Ловозерский р-н) – сборы Ф. Ниландера, 1844, H (FinBIF…) и в устье реки Восточная Лица (Pis’yaukova, 1956). Указание “п. Гремиха” (Ramenskaya, Andreeva, 1982), по всей вероятности, относится к точке сборов Ниландера, отмечаемой, также, в Атласе сосудистых растений Северной Европы (Hultén, Fries, 1986). Приводится для Хибин (Tolmachev, 1975, Hultén, Fries, 1986). Находки последних лет состоялись в “континентальных” районах Мурманской области: в заповеднике “Пасвик” на скальных склонах северо-восточной части горы Калкуля к озеру Каскамаярви (Kravchenko et al., 2016) и в Лапландском заповеднике – на северных склонах массива Мончетундра (озеро Вайкис: INEP (V) LID1504038, Borovichev et al., 2020a). Новые находки дополняют представление об экологии и характере распространения видов. Аннотации сборов свидетельствуют о приуроченности вида к скальным местообитаниям в ущельях, долинах ручьев и по берегам озер и предпочтению склонов северных вариантов экспозиции, по крайней мере, при удалении от морского побережья.
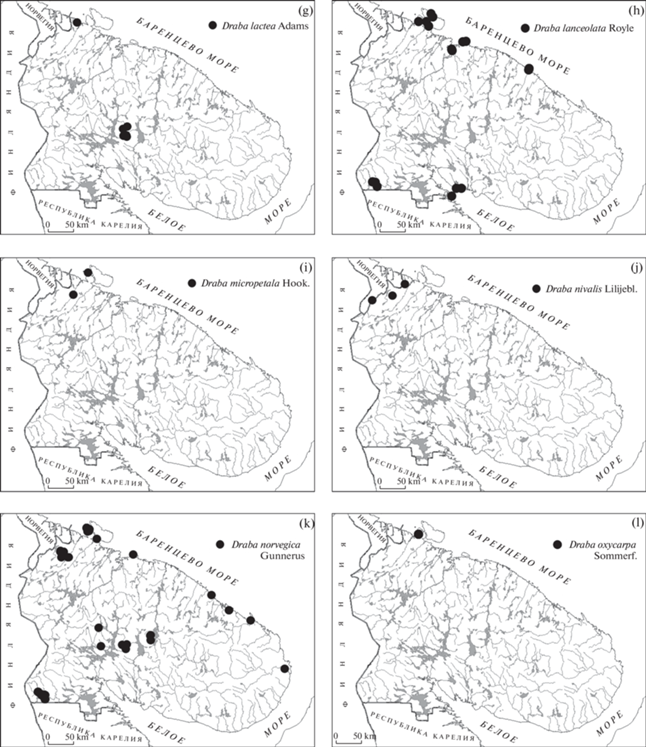
Рис. 1.
Карты распространения видов рода Draba в Мурманской области.
Fig. 1. Distribution maps of Draba species in the Murmansk Region
a – Draba alpina, b – D. cinerea, c – D. fladnizensis, d – D. hirta, e – D. insularis, f – D. incana, g – D. lactea, h – D. lanceolata, i – D. micropetala, j – D. nivalis, k – D. norvegica, l – D. oxycarpa.

Рис. 1.
Окончание
D. alpina, рассматриваемая как часть одноименного циркумполярного аркто-монтанного агрегата, является амфиатлантическим видом (Checklist…, 2007). Ее ближайшие к Мурманской области местонахождения расположены в Скандинавских горах и на Свальбарде (Шпицберген), в Европейской России – на Новой Земле, Канином Носу и на Полярном Урале.
Draba cinerea Adams – Крупка седая. Встречается только на юго-западе Мурманской области в приграничном с Карелией районе: в заказнике “Кутса” (рис. 1b). В отечественной литературе для Мурманской области вид ранее не указывался, но известен по многочисленным сборам и наблюдениям финских исследователей первой половины XX века (Н, TUR, OULU, Ulvinen, 1996). Косвенным подтверждением современного существования данной популяции являются сборы и наблюдения Т. Ульвинена 1994, 1996 гг. (OULU, Ulvinen, 1996), а также гербарный сбор А.А. Похилько 1998 г. “…окрестности п. Вуориярви, ущелье Пюхякуру, на уступах скал” (KPABG №№ 025348, 025361 (д)), ошибочно отнесенный автором к Draba hirta L. Draba cinerea предпочитает богатые кальцием экотопы. Уступы и расщелины скал, ущелья, скалистые склоны речных долин и берегов озер – основные местообитания вида в заказнике – характерны для “лесных” частей его общего ареала (Petrovskiy, 2018).
D. cinerea – вид с гипоарктомонтанным, почти циркумполярным типом ареала, который в Северной Европе становится фрагментарным и представлен несколькими локусами реликтового характера (Hultén, Fries, 1986, Petrovskiy, 2018, Draba cinerea…, 2021): новоземельским, колгуевским и популяциями тундровых возвышенностей Северной Норвегии. В лесной зоне известны 3 изолированных местонахождения – в Приладожье (var. ladogensis H. Lindb.), на севере Лапландии и в районе Оуланка (Финляндия), где довольно крупная популяция имеет трансграничный характер и продолжается на территории Карелии (скалы Рускеакаллио в НП “Паанаярви”: Kravchenko, 2007). По всей вероятности, местонахождение D. cinerea в Мурманской области является относительно изолированной частью оуланкско–паанаярвской части ареала (рис. 1b). Вид является редким и подлежит охране в соседних Финляндии и Карелии (Hyvärinen et al., 2019, Red…, 2020). Популяция, выявленная в Мурманской области, также нуждается в изучении и номинации в списки региональной Красной книги.
Draba fladnizensis Wulfen – Крупка фладницийская. Редкий вид, охраняется в Мурманской области (категория 3: RED…, 2014). Встречается в горно-тундровом поясе массивов центральной части области – в Хибинских и Ловозерских горах (Kozhin et al., 2020, коллекции KPABG, INEP, MW), найден на севере Мончетундры (озеро Вайкис: сбор Е.А. Боровичева, 2009, INEP LID-1504039). В последние годы найден в тундре Баренцевоморского побережья: окрестностях Лиинахамари (Kostina, Borovichev, 2014), на севере полуострова Рыбачий (Borovichev et al., 2020a) и в губе Ивановка (Восточный Мурман: Borovichev et al., 2018) (рис. 1c). Растет по сухим скалам и осыпям мелкозема глубоких ущелий: горные перевалы, геологические разломы периферии горных систем и морских побережий; необлигатный кальциефил. Ближайшие местонахождения вне области – северное побережье Норвегии и Скандинавские горы (где вид становится нередким), Канино-Печорский регион и Полярный Урал, общий ареал – циркумполярный арктоальпийский.
Draba hirta L. (D. daurica DC., D. glabella Pursh) – Крупка мохнатая. Встречается по всему Кольскому побережью Баренцева и Белого морей, найдена в более чем в половине территорий, посещавшихся ботаниками (KPABG, MW, INEP, KAND, H, TUR, Breslina, 1969, 1985, Bubenets et al., 1993, Vorob’yeva, 1996; рис. 1d). Ее основные местообитания сосредоточены вблизи берега моря: уступы и трещины скал, обнаженные пески, пятна мелкозема, в т.ч. антропогенного характера, приморские вороничники, тимьянники и редкотравные луговины, вторичные орнитогенные ценозы.
В материковой части находки D. hirta значительно реже и связаны, в большинстве случаев, с горно-тундровым и верхней частью горно-лесного пояса крупных массивов – Хибин, Ловозерских тундр (KPABG, MW) и Чунатундры (Лапландский заповедник: KPABG, Berlina, 1997): на каменистых и щебнистых склонах, в скальных ущельях, на галечниках по берегам рек и ручьев. На крайнем северо-западе найдена в Печенгских тундрах (сбор A. Cajander, 1928, H) и в заповеднике Пасвик: по скальным уступам склонов тундровых возвышенностей (сборы М.А. Фадеевой и А.В. Кравченко, 2014, PTZ), а также в лесной зоне: по скалистым берегам озера Каскамаярви (сборы В.А. Костиной, 1994, KPABG и А.В. Кравченко, 2009, PTZ). Многократно отмечалась в бассейне р. Кутсайоки (заказник “Кутса”: KPABG, TUR, H, Ulvinen, 1996), где растет по каменистым склонам, расщелинам и карнизам скал в ущельях, долинах рек и по берегам озер, часто совместно с другими видами рода Draba.
D. hirta – одна из самых распространенных крупок с циркумполярным ареалом арктобореально-монтанного типа. Как все таксоны с обширным, но фрагментированным ареалом, приводящим к формированию множества обособленных микропопуляций, она характеризуется большой морфологической вариабельностью (в разное время выделялись до 15 ее вариантов и 2 подвида) и запутанной синонимикой (более 35 наименований, по данным GBIF).
При просмотре Кольского гербария, кроме типичных, обнаружены образцы одного из вариантов D. hirta, отличающегося мелкими стручочками – var. brachycarpa Rupr.: сборы И.П. Бреслиной, В.Т. Царевой, 18 VII 1984, KPABG №№ 025341, 025342, KAND-7830. Образцы относятся к единственному местонахождению: “Кандалакшский залив Белого моря, р-н острова Великого, луда Наумиха. Южная часть плато, скальное обнажение среди вороничников, дернина вторичной орнитогенной растительности”.
С островов Кандалакшского залива в середине XX века был описан эндемичный вид Draba insularis Pissjauk. (D. valida Pissjauk., о. Ламбин, Pis’yaukova, 1956) – Крупка островная. Он встречается по островам Белого моря и в прибрежных районах Терского берега (Порья губа, Турий мыс и др.: KAND, MW, KPABG; рис. 1e). Растения, относимые к этому таксону, обладают значительным сходством с D. hirta , и имеют полный набор специфических признаков, отличающих D. hirta от других представителей рода. К наиболее значимым отличиям, позволяющим отделить D. insularis от D. hirta относят (Pis’yaukova, 1956, Lutskina, Bochansteva, 2004): рослость растения и мощность дернины с обилием цветоносов (размеры растения), большую длину столбика – 0.5–1.0 мм (против 0.3–0.5 мм у D. hirta), не выделяющееся (не утолщенное) и не раздвоенное рыльце, в качестве дополнительного признака – отсутствие опушения створок стручочков (иногда развивающегося у D. hirta).
У просмотренных нами беломорских образцов все признаки варьировали в соответствии с пределами, заявленными для обоих видов. Однако распределение их было хаотичным, так что лишь единичные экземпляры из выборки обладали их комплексом и могли быть с уверенностью отнесены к тому или иному виду. Проведенный (на выборке из 50 гербарных листов) ранговый корреляционный анализ Спирмена подтвердил отсутствие сопряженности данных признаков (табл. 1). Исключение составляет связь раздвоенности рыльца и его расширенности относительно столбика, обусловленная их морфологической созависимостью, однако, сила этой связи r = 0.42135 (статистически значимая при доверительной вероятности P = 0.95; выделена шрифтом) недостаточна, чтоб рассматривать даже эти два признака как единый и надежный диагностический комплекс.
Таблица 1.
Сопряженность (r) диагностических признаков беломорских образцов Draba insularis и Draba hirta. Table 1. Contingency (r) of diagnostic features in Draba insularis and Draba hirta samples from the White Sea.
| Размеры куртины Cluster size |
Длина столбика Style length |
Рыльце, расширение Stigma, extension |
Рыльце, раздвоение Stigma, bifurcation |
|
|---|---|---|---|---|
| Размеры куртины Cluster size |
Х | 0.077982 | 0.069072 | 0.23102 |
| Длина столбика Style length |
0.077982 | Х | 0.164204 | 0.109325 |
| Рыльце, расширение Stigma, extension |
0.069072 | 0.164204 | Х | 0.42135 |
| Рыльце, раздвоение Stigma, bifurcation |
0.23102 | 0.109325 | 0.42135 | Х |
Молекулярно-генетический анализ образцов обоих видов, выбранных из разных – материковой и островной – популяций, в настоящем исследовании не может дать однозначный ответ на вопрос, единый ли это таксон, хотя скорее говорит об их тесной родственной близости, чем о разобщенности. Образцы с Терского берега и Медвежьего острова идентичны не только между собой но и с последовательностями нескольких других видов (рис. 2). При этом сравниваемые с ними наиболее близкие к ним последовательности образца D. hirta из ГенБанка (isolate L310, DQ467486) принадлежат совсем другой ветви, в несколько раз перекрывая различия наших образцов, что вполне согласуется с отмечаемой фенотипической полиморфностью D. hirta из разных частей ареала.
Рис. 2.
Результаты сравнительного анализа секвенированных геномных последовательностей изученных образцов видов р. Draba (обозначены серыми кружками) и наиболее близких к ним последовательностей из ГенБанка.
Fig. 2. Results of molecular sequence analysis of the studied Draba samples (marked with gray circles) and the closest sequences from the GenBank.

Следует отметить, что крупные экземпляры D. hirta c довольно длинным (0.5–0.7 мм) столбиком и нераздвоенным рыльцем встречаются в Хибинах (KPABG 025355, 025365) и в Печенгском районе (заповедник Пасвик, PTZ №№ 22963, 22099), где также присутствуют и “короткостолбиковые” особи среднего размера (PTZ № 26933). Это сильно расширяет границы распространения растений с признаками D. insularis. Как правило, их местообитания связаны с относительным благополучием среды (скальные полки в ущельях низкогорий, в условиях достаточного увлажнения почвы и воздуха), что может объяснить их рослость и разрастание куртин, равно как и в случаях островных популяций крупок.
В мировых систематических обзорах северных крупок D. insularis не приводится в силу своего более южного, северотаежного ареала, находящегося за границами Арктической области (Checklist…, 2007). Исключение составляет Атлас сосудистых растений Северной Европы (Hultén, 1971) где она не рассматривается в качестве самостоятельного видового таксона (включена в D. hirta). В переиздании атласа (Hultén, Fries, 1986) она уже не упоминается. Полученные нами результаты свидетельствуют в пользу этого мнения: по всей вероятности, D. insularis Pissjauk. является одной из форм полиморфного вида D. hirta, встречающейся в условиях благоприятного микроклимата и минерального питания.
Draba incana L. – Крупка седеющая. Преимущественно приморский вид, в Мурманской области – самый распространенный из рода Draba (рис. 1f). Встречается практически во всех районах Баренцевоморского и Беломорского побережья и на островах с небольшим разрывом ареала в районе от Восточной Лицы до Йоканьги (Восточный Мурман). На “материк” достаточно далеко заходит по долинам крупных рек: Колы, Печенги, Поноя; наблюдалась в заказнике “Кутса” на скалистых берегах озера Ниваярви (FinBIF…). Во Флоре Мурманской области (Pis’yaukova, 1956) приводится местонахождение в Хибинах (отмеченное на карте вопросительным знаком: рис. 1g). Однако, единственный известный нам хибинский образец – сбор с восточного склона г. Юкспор Фоминой, определенный как D. incana L. Б.А. Мишкиным (1937, KPABG) – не был учтен при составлении флоры Хибин (Mishkin, 1953), а в 1955 г. при обработке коллекции для Флоры Мурманской области был отнесен В.В. Письяуковой к D. hirta (авторы данной работы придерживаются того же мнения). В связи с этим, нахождение Draba incana в Хибинах вызывает сомнения.
Основные местообитания D. incana сосредоточены у моря: луговины на береговом валу и морских террасах низкого уровня, где она становится массовой и иногда достигает значительного обилия, песчаные осыпи и дюны, устьевые зоны рек, расщелины и уступы скал, участки орнитогенной растительности и реже – приморские вороничники. На местах бывших поселений встречается на развалинах, обнажениях грунта, по лугам антропогенного происхождения, в т.ч., по бывшим сенокосам, при этом может “заходить” на несколько км от моря. Все находки в “континентальных” районах связаны с долинными экотопами крупных рек: злаково-разнотравные луга на террасах, песчаные склоны, скалистые берега, либо имеют антропогенное происхождение, как известная уже почти 100 лет популяция с зимнего Лумбовского погоста, занесенная туда во время перекочевок оленеводов с летних, приморских пастбищ (Tsinzerling, 1935, Borovichev et al., 2020b).
Приморский тип ареала характерен для D. incana в Фенноскандии в целом (FinBIF…), однако в ее западной части с более океаничным климатом вид более распространен в глубине “материка” по долинам современных или плейстоценовых рек и фьордов (как, например, в регионе Инари-Паз), по западному макросклону Скандинавских гор и спорадически появляется во “внутренних” районах Финской Лапландии. Такой же стратегии расселения вид придерживается на всем протяжении своего арктобореально-монтанного амфиатлантического ареала, за исключением пиренейских популяций. (Hultén, Fries, 1986, Checklist…, 2007, GBIF…).
Draba lactea Adams – Крупка молочно-белая. В Мурманской области очень редкий вид (рис. 1g), занесен в ККМО (категория 2). Местонахождения в Хибинских горах, известные по сборам 30–70-х годов XX века (KPABG), расположенные на вершинах и предвершинных частях склонов в южной и центральной части массива (сырые скалы, россыпи камней), уже отчасти уничтожены горными разработками (г. Расвумчорр, г. Юкспорр). В 2013 г. найден на крайнем северо-западе области в районе п. Лиинахамари (мыс Немецкий): в тундровой возвышенности на уступе влажной скалы близ озера оз. Кантеярви (Kostina, Borovichev, 2014). Значимость этой находки велика не только в природоохранном отношении, но и как связующее звено в интерпретации возможного расселения этого вида: связи между изолированными Кольскими популяциями с основным ареалом в Фенноскандии – Скандинавскими горами. Другими ближайшими местами произрастания вида являются Канино-Печорская тундра, Новая Земля и Уральский хребет. Общий ареал вида циркумполярный арктомонтанный.
Draba lanceolata Royle (D. cana Rydb.) – Крупка ланцетоплодная. Приводимый во Флоре Мурманской области (Pis’yaukova, 1956; авторы придерживаются описания таксона, сделанного в данном источнике), довольно редкий в регионе вид, ареал которого состоит из 3 кластеров (рис. 1h). Большинство находок сделано в приморских районах Баренцевоморского побережья: полуострова Рыбачий и Средний – сборы А.В. Разумовской и К.Б. Поповой 2014–2018 гг., (INEP); о-ва Екатерининский и Медведка Кольского залива – сборы В. Кудряшова, 1920 (MW), Н. Прохорова и О. Кузеневой, 1921 (KPABG); о. Кильдин – указание во Флоре Мурманской области (Pis’yaukova, 1956) и сбор В.Т. Царевой, 1977 (KPABG), архипелаг Семь островов – остров Харлов – сборы М.В. Мельникова и Р.В. Плотникова 2015, 2017 гг. (KAND) и тундра на материковом побережье (Н.Г. Журавлева, 1964, MW). Здесь его местообитания в основном связаны с берегом моря: луга на песчаных дюнах и береговом валу, трещины приморских скал, осыпи морских террас. Заходит на антропогенно-нарушенные участки: в центральной возвышенности п-ова Средний обнаружен на территории бывшего военного городка, где расселяется по насыпям и зарастающим угольным кучам. Вторая часть ареала – условно “прибеломорская” – представлена отдельными местонахождениями в Порьей губе (И.П. Бреслина, 1984, KPABG, М.Н. Кожин, 2008, KAND) и на о-ве Великом (Н.С. Парфентьева, 1951, KAND) на приморских сухих луговинах, в том числе, орнитогенного характера. На “материке” D. lanceolata встречается на крайнем юго-западе области (территории заказника “Кутса”) по скалистым берегам рек Кутсайоки и Онтонйоки, а также в урочище Ущелье Пюхякуру, в расщелинах скал и по крутым склонам (сборы А.А. Похилько, 1998, KPABG №№ 025346, 025347, 025349).
Возможно, таксон более распространен в области, т.к. на протяжении всего ареала он встречается в одних и тех же местообитаниях, а в ряде случаев произрастает совместно с близким (и гораздо более распространенным) видом D. incana L. и может просматриваться или идентифицироваться в его рамках. Так, гербарные образцы с Екатерининского острова, острова Харлов и с Порьей губы (MW №№ 0372024, 0372027, 0372018, KPABG 025383, часть особей на листах KAND-4417 и KAND-4355) были отнесены авторами сборов и первоначальных определений к D. incana. Мелкие растения со слабоолиственным стеблем c территории ГПЗ “Кутса” и с о. Кильдин (KPABG №№ 025333, 025346, 025347, 025349, 025354, 025360) внешне сходны с D. hirta, и были идентифицированы в гербарии как этот последний вид, однако отсутствие на листьях и цветоносе волосков специфического типа звездчатости с вторичными отростками (“гиртоидный тип”) и некоторые другие признаки позволили при пересмотре коллекции с уверенностью отнести их к Draba lanceolata.
Достаточно широкий характер расселения D. lanceolata в Мурманской области предполагает также присутствие этого вида в соседних регионах, особенно в западном направлении, в Фенноскандии. Однако мурманские популяции Draba lanceolata оказались в полной изоляции относительно основного, азиатско-североамериканского ареала этого вида. На Европейском континенте имеется еще одно указание на местонахождение D. cana: Восточные Пиренеи, на границе Испании и Франции (“Núria. Cami d’Eyne”: Guardia, 2021; к сожалению, нам не удалось видеть гербарный образец). При этом в высокогорьях Азии, Кордильерах и в субарктических районах восточной Сибири вид отмечается довольно часто и иногда заходит в Арктические районы Аляски, Канады, и на побережье Гренландии (Tolmachev, 1975, Checklist…, 2007, GBIF…).
Draba micropetala Hook. – Крупка мелколепестная. Новый вид для Мурманской области. Обнаружен при просмотре гербарной коллекции Ботанического Музея Университета Хельсинки (H) на двух гербарных листах:
1. H № 302191, сбор Alexander Berg 16 VII 1937, определенный как Draba hirta L. Местонахождение: Печенгский р-н, гора Спасительная (ныне гора Генеральская) близ Луостари, скальная стенка;
2. H № 804696, сбор Ernst Häyrén 6 VIII 1926, определенный как Draba rupestris R. Br. Местонахождение: Печенгский р-н, п-ов Средний, возвышенность Кивиайдантунтури, в альпийском поясе.
Детальным обследованием окрестностей г. Кивиайдантунтури на п-ове Среднем в 2021 г. удалось отчасти подтвердить сбор Эрнста Хайрена. Среди многочисленной популяции Draba norvegica (с присутствием D. hirta) на скальных стенках уступов, обращенных к морю, были обнаружены особи, значительно уклоняющиеся к D. micropetala (69.814786°с.ш., 31.837453°в.д., сбор А.В. Разумовской INEP LID-1505313). От D. norvegica они отличаются деталями опушения листа (гораздо большим размером штока 4-х лучевых волосков), преобладающей формой плодов – от обратно-яйцевидной до овальной (заостренная у D. norvegica) и листьев – с несколько закругленной верхушкой в отличие от заостренных листьев D. norvegica. Установить принадлежность этих экземпляров к типичной D. micropetala не дает довольно широкая форма лепестков (одним из диагностических признаков этого вида являются узкие лепестки, равные или слегка превышающие ширину чашелистиков), сходная с таковой основного вида местонахождения. Остальные признаки являются общими для обоих видов (имеется в виду арктическая биоморфа D. norvegica с полностью безлистным цветоносом). На гербарных листах из коллекции H все экземпляры были на стадии плодоношения, из-за отсутствия лепестков сравнить их с найденным в поле материалом не удалось. Возможность гибридизации между D. micropetala и указанными выше видами в литературе еще не определена, несмотря на обилие данных, подтверждающих, в том числе экспериментально, высокую вероятность образования фертильных гибридов у большинства видов рода (Brochmann, 1992).
D. micropetala – высокоарктический вид с циркумполярным типом ареала, распространен на островах Арктического бассейна (арх. Свальбард, Земля Франца-Иосифа, Северная Земля, Новая Земля и др.). На материке появляется на Полярном Урале и в тундровой зоне Северной Азии, на Юконе и в Канаде (Hultén, Fries, 1986, Checklist…, 2007, GBIF…). Ее местонахождения в Мурманской области (рис. 1i) являются единственными в материковой части Северной Европы. Необходимы дальнейшие исследования по их актуализации в природе, выяснению количественных параметров и экотопологической приуроченности популяций, генетических особенностей (гибридности), а также определение комплекса мер по их сохранению (номинация в ККМО).
Draba nivalis Liljebl. – Крупка снежная. Очень редкий вид в Мурманской области (ККМО – 1b), встречающийся на тундровых возвышенностях крайнего северо-запада (рис. 1j). В настоящее время достоверно известны два его местонахождения: в заповеднике Пасвик – северо-восточная часть горы Калкупя, на скальной гриве северо-западного берега оз. Каскамаярви – сбор Е.А. Боровичева, 23 VIII 2015 (PTZ; Kravchenko et al., 2016) и на востоке Печенгских тундр – окрестности озера Хаукилампи, гора Вуоггоайви – сбор Niilo Söyrinki, 1933 (OULU). Следует отметить, что в настоящее время территория вокруг озера Хаукилампи, и в частности, северные склоны горы Вуоггоайви входят в черту города Заполярный и сильно нарушены деятельностью горно-металлургических предприятий (карьеры, отвалы и пр.). Велика вероятность утраты данного местонахождения.
Сведения о распространении D. nivalis на п-ове Рыбачий (Hultén, 1971, Ramenskaya, 1983), к сожалению, не подтвердились. Все гербарные образцы, относящиеся к этой точке: сборы Justus Montell 1935 г., TURA370646 и TURA370799 при ревизии оказались принадлежащими к Draba norvegica.
Крупка снежная считается довольно широко распространенным аркто-альпийским циркумполярным видом (Checklist…, 2007). Она нередка в Скандинавских горах и на северном побережье Норвегии, и очевидно, что по северо-западу Мурманской области проходит восточная граница ее фенноскандийской части ареала (GBIF…).
Draba norvegica Gunnerus (D. rupestris W.T. Aiton) – Крупка норвежская. Редкий в Мурманской области вид (ККМО – 3). Встречается по Баренцевоморскому побережью и в крупных горных массивах области (Хибины, Ловозерские и Печенгские тундры: INEP, KPABG, H, TUR, NS, Ramenskaya, Andreeva, 1982; рис. 1k). Единичные находки сделаны в Чунатундре (Berlina, 1997) и Мончетундре (Е.А. Боровичев, 2017, INEP). Нередок, судя по обилию наблюдений и гербарных сборов, в бассейне р. Кутсайоки (заказник Кутса: H, OULU, TUR, Ulvinen, 1996). На Беломорском побережье не найден, и в целом, более распространен в западной части области. Его основные местообитания – сухие скальные уступы, расщелины, реже – осыпи и россыпи мелкозема, сырые скалы по долинам рек и ручьев, очень редко – дорожные насыпи и откосы (Хибины, п-ов Рыбачий).
D. norvegica – амфиатлантический арктоальпийский вид, почти обычный в западной Фенноскандии (GBIF…). Мурманские популяции находятся близ восточной границы основного ареала. Восточнее он становится очень редким и известен из отдельных локусов – о-ва Колгуев, Вайгач, Новая земля, Полярный Урал и восток Большеземельской тундры (Tolmachev, 1975, Checklist…, 2007)
Draba oxycarpa Sommerf. – Крупка остроплодная. Новый вид для Мурманской области. Найден в 2015 г. на центральной возвышенности п‑ова Средний (рис. 1l) в 500 м к югу от оз. Тунтури, на территории бывшего военного городка, среди редкотравной луговины по россыпи щебня и на угольной куче, растущий совместно с Draba incana (69.704125° с.ш., 31.776772° в.д., сборы А.В. Разумовской, К.Б. Поповой, INEP). Был ошибочно определен как D. nivalis, затем рассматривался как очень нетипичный образец Draba norvegica. Уточнить видовую принадлежность удалось только с помощью молекулярно-генетических исследований, которые показали, что анализированные образцы находятся в неразрешенном положении по отношению к изученным образцам D. incana и D. norvegica (достаточно далеки от обоих, рис. 2). Натурные наблюдения 2019 г. в момент цветения (а именно, бледно-желтый цвет лепестков, в отличие от белоцветковых D. norvegica и D. nivalis) полностью подтвердили результаты секвенирования и позволили, наряду с другими диагностическими признаками (опушение створок плодиков по рамке, форма чашелистиков и лепестков и др.), утвердиться в определении. При тщательном обследовании популяции в 2018–2021 гг. были определены ее границы и размер. D. oxycarpa заселяет небольшую площадь 100 × 20 м по пологому краю холма (древняя морская терраса) и по перегибу склона (69.703932° с.ш., 31.775197° в.д.), большая часть особей – на естественной, относительно ненарушенной, выровненной поверхности незадернованного сланцевого щебня и мелкозема, а также, по брустверам, стенкам и днищу окопов старых огневых точек, всего не менее 150 особей. В последующие за первой находкой годы D. oxycarpa у строений более не наблюдалась, но непосредственно в развалинах и на зарастающей бетонной крыше ангара выявлена небольшая популяция Draba lanceolata Royle и многочисленные гибридные Draba, образующие ряд переходных форм между D. incana, D. lanceolata и D. oxycarpa. Результаты сравнительного молекулярно-генетического анализа (сравнительного анализа внутригеномного полиморфизма первого внутреннего транскрибируемого спейсера ITS1) показали, что анализируемые участки генома всех, даже морфологически полярных (насколько это возможно в роде Draba) фенотипически “чистых” образцов D. oxycarpa и отобранного из удаленной контрольной точки образца D. incana, принадлежат единому гибридному комплексу (около 96% во всех четырех образцах – это риботипы Otu 1 (Otu – operational taxonomic unit): рис. 3). Внутри комплекса по признакам минорных риботипов (Otu 3, 4, 5, 6, 7) можно выделить две пары образцов. Пара ×1 и ×3 соответствует “чистой” D. oxycarpa (×1) и “контрольной” D. incana (×3). Вторая пара образована произрастающими рядом друг с другом Draba lanceolata (×4) и гибридной формой D. oxycarpa (×2). Наиболее близким к этому комплексу, как и к изучаемым образцам D. oxycarpa, согласно результатам секвенирования по Сенгеру, оказался образец Draba juvenilis DQ467228 из ГенБанка. Интерпретировать последний результат представляется затруднительным, т.к. Draba juvenilis морфологически достаточно хорошо отличим от всех видов комплекса, включая и Draba oxycarpa, и наши образцы соответствовали принятой трактовке этих различий (Rønning, 1979, Al-Shehbazz et al., 2010).
Рис. 3.
Гистограмма относительного числа риботипов (Otu 1–7) в образцах Draba oxycarpa (×1), ее гибридной формы (×2), D. incana (×3) и D. cana (×4).
Fig. 3. Histogram of a relative number of ribotypes (Otu 1–7) in the samples of Draba oxycarpa (×1), its hybrid form (×2), D. incana (×3) and D. cana (×4).

В традиционной трактовке систематики крупок D. oxycarpa (D. gredinii E. Ekman) долгое время рассматривался как вид, очень близкий к Draba alpina L. (Rønning, 1979, Mossberg, Stenberg, 2003., Al-Shehbazz et al., 2010), а в российских исследованиях не выделялся из этого крупного и морфологически полиморфного вида (аггрегата видов). Впервые о распространении Draba oxycarpa в Российской Арктике заговорили при создании проекта “Панарктической Флоры” (Checklist…, 2007), когда ревизия арктических Draba выявила его гербарные образцы с Новой Земли и Полярного Урала. Пересмотр коллекции D. alpina LE (сборы первой половины XX века) подтвердил наличие экземпляров с признаками, отличающими этот вид. Кроме того, растения, относящиеся к D. oxycarpa, обнаружились среди современных новоземельских и вайгачских сборов D. alpina Е.Ю Чураковой, 2013 (AR) и с водораздельной части Полярного Урала (река Большая Пайпудына, сборы Н.А. Секретаревой, С.С. Холода, 2005).
Таким образом, распространение D. oxycarpa в России, по современным сведениям, ограничивается Европейским сектором Арктики. Общий ареал вида – амфиатлантический, преимущественно островной с заходом в материковые тундры по возвышенностям севера Европы. При этом кольская популяция с полуострова Средний географически скорее расширяет восточную границу фенноскандинавской части ареала (вид встречается в горах севера Норвегии), чем является изолированным анклавом новоземельско-полярно-уральского. Наши данные секвенирования поддерживают это. Несмотря на характерные внешние признаки особей, позволяющие с уверенностью отнести их к комплексу D. alpina–D. oxycarpa, их сравнение с новоземельскими и восточно-сибирскими образцами данных видов не выявило такого же сходства на молекулярно-генетическом уровне. Найденная популяция, пока единственная на северо-западе Европейской России, с особенностями генома, вызывающими ряд вопросов, важных для понимания процессов эволюции арктических крупок, заслуживает дальнейшего изучения и нуждается в охране.
Молекулярно-генетические построения. Сводная таблица результатов сравнительного анализа секвенированных геномных последовательностей видов р. Draba представлена на рис. 2. В таблице указаны парсимонично-информативные нуклеотидные замены в некодирующих (ITS1, ITS2) и кодирующем участке (5.8S) рибосомного гена по референсному образцу D. nivalis DQ467598. Нуклеотидных замен крайне мало, большая часть замен располагается в первом спейсере. В кодирующей части имеется только одна замена (позиция 305), однако она характерна для разных видов из разных мест сбора – D. oxycarpa, D. juvenilis, D. norvegica, D. incana и D. lactea. Такая же картина наблюдается и в случае замен в декодирующих участках – разные виды имеют одинаковые нуклеотидные замены. На наш взгляд, это признак сложного гибридного комплекса, выявленного на примере группы Draba oxycarpa–Draba lanceolata–Draba incana.
ОБСУЖДЕНИЕ
Виды рода Draba, встречающиеся в Мурманской области, можно условно разделить на две группы – виды преимущественно арктические, чей оптимум ареала приходится на высокоширотные Арктические острова и материковые тундры за пределами влияния Гольфстрима, и виды гипоаркто- и арктобореально-монтанные, распространенные более широко в Гипоарктике и в Бореальной области, в высокую Арктику заходящие лишь в условиях мягкого морского климата. К первой группе принадлежат все самые редкие в Мурманской области Draba: D. alpina, D. lactea, D. micropetala, D. nivalis, D. oxycarpa, до некоторой степени D. fladzinensis (с заметной горной частью ареала за пределами Арктики). Их находки единичны, у некоторых уникальны, локальные популяции исчисляются первыми десятками особей. Здесь проходит южная граница европейской части их ареала, пунктирно (и не для всех!) продолжающаяся в Скандинавских горах Северной Норвегии. Эти виды, согласно концепции истории развития и распространения рода Draba А.И. Толмачева (Tolmachev, 1957), принадлежат рядам относительно эволюционно продвинутых таксонов, по всей вероятности, сформировавшихся уже в Арктике с приобретением характерных признаков ксерофитизиции и криофитизации форм.
Самым ярким представителем второй группы является амфиатлантический D. incana – активный вид, прекрасно акклиматизировавшийся на морском берегу, не избегающий ни орнитогенных, ни рудеральных местообитаний. Другие – D. hirta и D. lanceolata – чуть менее распространены, но и менее климатически стенотопны, в несколько больших пропорциях встречаясь в “континентальных” районах области. Несмотря на явно приморский характер экотопических предпочтений амфиатлантический тип ареала вовсе не характерен для видов этой группы. D. lanceolata – преимущественно азиатско-западно-североамериканский вид (в связи с его очевидной фенотипической близостью с Draba incana правомерно предположение о их викариате в пределах Голарктики с зоной перекрытия ареалов в восточной Фенноскандии). И D. hirta, и D. cinerea (единственный, по-настоящему редкий вид этой группы) также имеют ареал, не ограничивающийся Атлантическим регионом. По мнению А.И. Толмачева, именно эти виды в наибольшей степени сохранили архаичные черты, присущие анцестральным формам, существовавшим в условиях относительно мягкого климата среднегорий. При этом D. incana рассматривалась в качестве одного из наиболее архаичных таксонов. Аналогичный вывод насчет анцестральности D. incana, по крайней мере, по отношению к современным полиплоидным Leucodraba, в т.ч. из нашей “второй группы”, был сделан на основании молекулярно-генетических и палинологических исследований в конце XX века (Brochmann, 1992).
По характеру распространения в Мурманской области амфиатлантический, заходящий в высокоарктические районы вид Draba norvegica занимает промежуточное положение между этими двумя группами. Его двойственность подчеркивается (а возможно и объясняется) присутствием двух биоморф, различающихся по набору архаичных и продвинутых признаков, главными из которых является безлистность цветоноса, уплотненная розетка листьев и общий мелкий габитус, появляющиеся у арктических популяций. По нашим наблюдениям, на Баренцевоморском побережье обе формы могут встречаться в пределах одного местообитания.
Несмотря на отмечаемые различия встречаемости, зональной приуроченности и истории формирования таксонов, в географии видов обеих групп в пределах исследуемого региона прослеживается единая закономерность. Ключевую роль в их расселении играет морское побережье. Все места находок редких видов арктической группы, массового распространения гипоарктических Draba и в целом повышенного видового разнообразия рода в Мурманской области, так или иначе, связаны с морем. На картах очевидна связь большинства точек местонахождений с современной береговой линией. Находки же, сделанные в “континентальной” части области, приурочены к нескольким разобщенным регионам со сходными, как оказалось, событиями истории плейстоцена – морскими трансгрессиями.
Долина реки Паз (заповедник “Пасвик”), как и Печенги, и других, менее протяженных рек крайнего северо-запада Мурманской области, является плейстоценовым фьордом. Современные озера Имандра, Умба и Ловозеро были морскими заливами, и Хибины, Ловозерские тундры, массивы Мончетундра и Чунатундра становились приморскими перигляциальными территориями (островами), как минимум, дважды в неогене (Kiselev, 1979, Evzerov, 2015). Одним из самых значительных центров сосредоточения Draba (D. lanceolata, D. cinerea, D. hirta, D. incana, D. norvegica) является район природного парка “Кутса”, расположенный в удалении от моря на юго-западе области. Здесь прослеживается стадиальная граница покровного Скандинавского ледника (стадия Сальпауселька II: Gosudarstvennaya…, 2012). По данным палеогеографии этого региона, в Микулинское межледниковье его долины рек, озерные котловины и ущелья отчасти (до современных отметок рельефа 200–211 м) заливались морскими водами (Kiselev, 1979), а судя по соседству моренных, флювиогляциальных и морских отложений верхнего звена неоплейстоцена с протяженными скальными выходами нерасчлененных дочетвертичных пород и характерным формам рельефа, в позднеледниковое время (и также в пребореал), по всей вероятности, были фьордами (Gosudarstvennaya…, 2012, Karta…, 2014).
Современный характер расселения видов рода Крупка в Мурманской области представляет собой остатки некогда обширных ареалов видов перигляциальных ландшафтов, передвигавшихся вслед за обнажавшимися поверхностями покровного ледника, включая и шельфовые области, осушавшиеся во время его наступления (когда по шельфовым пространствам происходил активный взаимообмен с флорами островов Полярного бассейна). Многократные перемещения и смены климатической обстановки не изменили изначально ореофитный “характер” крупок, и по сей день выражающийся в предпочтениях открытых щебнистых грунтов и скальных местообитаний. Одни (виды II группы) сумели адаптироваться в постледниковье и занять подходящие (приморские) экологические ниши в разогревающейся Гольстримом Фенноскандии, другие сохраняются лишь точечно, в пределах рефугиумов, являясь реликтами. При этом, в неблагоприятных для них условиях (в удалении от моря), характер расселения видов II группы также становится рефугиальным. Виды растут компактно, часто совместно друг с другом, в одном или нескольких изолированных друг от друга, как правило, скальных местообитаниях.
Совместное произрастание разных крупок в удаленных друг от друга рефугиумах, отмечается как для отдельных аггрегатов, так и для менее близкородственных видовых групп не только на Кольском п-ове, но и в других районах Арктической области (Al-Shehbazz et al., 2010, Petrovskiy, 2018), где сходные геоморфологические процессы плейстоценового и ранне-голоценового времени приводили к пульсациям и подвижкам ареалов. Периоды изоляции, равно как и резкого расширения ареалов активизировали дивергентную направленность процессов видообразования, в ряде случаев приводящую к возникновению в разных частях ареала автогамных внутривидовых форм, обладающих репродуктивными барьерами (cryptic species, Brochmann et al., 2004, Gustafsson et al., 2014), формированию серий близкородственных таксонов (Petrovskiy, 2018) и таких сложных явлений как рекуррентное и полифилетичное формирование полиплоидных видов (Brochmann, 1992, Grundt et al., 2004). Этим во многом обуславливается кажущаяся запутанность филогенетических построений у крупок (включая и дерево, приводимое в данной работе), где полиморфные таксоны могут далеко расходиться по разным кладам. С другой стороны, перекрестное опыление тесно соседствующих видов не исключает также возможность запуска de novo процессов межвидовой гибридизации. Признаками этого на современном временном срезе могут служить не только присутствие в Кольских популяциях фертильных отклоняющихся, гибридных и уродливых особей, но и формирование единых гибридогенных комплексов в геномах растений, по фенотипу принадлежащих к разным, не близким таксонам. Вопрос о роли вторичной гибридизации в процессах видообразования в роде Draba остается открытым, однако, по всей вероятности, она также вносит свою лепту в сложность интерпретации результатов молекулярно-генетического анализа, представленных на рис. 2. Выбранный для данного исследования маркер показал недостаточно высокую информативность для решения сложных проблем филогенетики Draba. Привлечение более широкого арсенала молекулярных методов позволит внести ясность в эти вопросы.
Реликтовый характер современного распространения видов арктической группы Draba, равно как и возможная потеря видоспецифичности их генофонда путем вторичной гибридизации требуют повышенного внимания к этим видам со стороны биологов. Они, безусловно, заслуживают охраны, как и ранее не отмечавшийся в составе флоры Мурманской области Draba cinerea. Виды обеих групп, особенно в континентальных районах области могут служить маркерами перспективных мест сосредоточения других редкостей и реликтов: Arenaria humifusa, Arnica alpina, Erigeron borealis, Saxifraga tenuis, Thymus subarcticus и др. – в уже выявленных местоположениях они также отмечаются и часто растут совместно (наши данные, Ulvinen, 1996, Kozhin et al., 2020). Такие места, как правило, характеризуются повышенными уровнями видового богатства, и могут рассматриваться в качестве ключевых ботанических территорий (Borovichev et al., 2011, Kozhin et al., 2019, Belkina et al., 2021). В связи с этим, привлечение методов реконструкции палеоландшафтов (поиск сочетания краевых зон морских трансгрессий с сохранившимися скальными комплексами перигляциальных зон) может быть использовано как в прогнозировании новых находок редких Draba (и им подобных видов), так и при оценке перспективности территорий для создания новых и корректировки границ существующих ООПТ.
Список литературы
Al-Shehbaz A.I., Windham M.D., Elven R. 2010. Draba Linnaeus. – In: Flora of North America. Vol. 7: Magnoliophyta: Salicaceae to Brassicaceae. New York, Oxford. P. 269–347.
Aronesty E. 2013. Comparison of Sequencing Utility Programs. – Open Bioinform. J. 7: 1–8. https://doi.org/10.2174/1875036201307010001
[Belkina et al.] Белкина О.А., Обабко Р.П., Боровичев Е.А., Лихачев А.Ю. 2020. Мхи района озера Вайкис (горный массив Мончетундра, Мурманская область) – ключевой ботанической территории. – Новости сист. низш. раст. 54 (2): 479–495. https://doi.org/10.31111/nsnr/2020.54.2.479
Benson D.B., Karsch-Mizrachi I., Lipman D.J., Ostell J. Wheeler D.L. 2005. GenBank. – Nucleic Acids Res. 33: 34–38.
[Berlina] Берлина Н.Г. 1997. Сосудистые растения Лапландского заповедника (аннотированный список видов). Сер. Флора и фауна заповедников. Вып. 64. М. 38 с.
Bolger A.M., Lohse M., Usadel B. 2014. Trimmomatic: A flexible trimmer for Illumina sequence data. – Bioinformatics. 30: 2114–2120. https://doi.org/10.1093/bioinformatics/btu170
[Borovichev et al.] Боровичев Е.А., Кожин М.Н., Игнашов П.А., Кириллова Н.Р., Копеина Е.И., Кравченко А.В., Кузнецов О.Л., Кутенков С.А., Мелехин А.В., Попова К.Б., Разумовская А.В., Сенников А.Н., Фадеева М.А., Химич Ю.Р. 2020a. Значимые находки растений, лишайников и грибов на территории Мурманской области. – Труды Карельского научного центра РАН. 1: 17–33. https://doi.org/10.17076/bg1078
[Borovichev et al.] Боровичёв Е.А., Королёва Н.Е., Кожин М.Н., Зацаринный И.В., Химич Ю.Р., Петрова О.В., Сенников А.Н., Костина В.А. 2020b. Наземные экосистемы Мурманской области под воздействием человеческой деятельности и изменений климата. – В кн.: Природа и коренное население Арктики под влиянием изменения климата и индустриального освоения: Мурманская область. М. С. 18–41.
[Borovichev et al.] Боровичев Е.А., Костина В.А., Шалыгин С.С. 2011. Некоторые ключевые ботанические территории Лапландского заповедника (Мурманская область). – Труды Карельского научного центра РАН. 12 (2): 48–55.
[Borovichev et al.] Боровичев Е.А., Разумовская А.В., Белкина О.А., Обабко Р.П. 2018. Новые находки охраняемых видов растений в Мурманской области: Баренцевоморское побережье. – Труды Карельского научного центра РАН. 1: 23–32. https://doi.org/10.17076/bg668
[Breslina] Бреслина И.П. 1969. Флора и растительность семи островов и прилегающего побережья Восточного Мурмана. – В кн.: Труды Кандалакшского государственного заповедника. Выпуск VII. Ботанические исследования. Мурманское книжное издательство. С. 259–382.
[Breslina] Бреслина, И.П. 1985. Флора острова Наумихи (Кандалакшский залив Белого моря). – В кн.: Ботанические исследования за Полярным кругом. Апатиты. С. 18–23.
Brochmann C. 1992. Polyploid evolution in Artic-Alpine Draba (Brassicaceae). Sommerfeltia Supplement. 4: 1–37.
Brochmann C., Brysting A.K., Alsos I.G., Borgen L., Grundt H.H., Scheen A.-C., Elven R. 2004. Polyploidy in arctic plants. – Biol. J. Linn. Soc. 82: 521–536.
[Bubenets et al.] Бубенец В.Н., Похилько А.А., Царева В.Т. 1993. Сосудистые растения Турьего мыса. – В кн.: Флористические исследования в Мурманской области. Апатиты. С. 45–73.
Checklist of the Panarctic Flora (PAF). Vascular plants. 6721 Draba L. 2007. Oslo. http://panarcticflora.org
Doyle J.J., Doyle J.L. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. – Phytochem. Bull. 19: 11–15.
Draba cinerea. Occurrence. 2021. FinBIF. https://laji.fi/en/taxon/MX.38432/occurrence (Accessed 7.03.2021).
[Evzerov] Евзеров В.Я. Геология четверичных отложений Кольского региона. 2015. Апатиты. 196 с. https://www.geokniga.org/bookfiles/geokniga-geologiya-chetvertichnyh-otlozheniy-kolskogo-regiona.pdf
FinBIF Submission. http://tun.fi/MKA.99158529 (Accessed 12.03.2021).
FinBIF: Finnish Biodiversity Information Facility. https://laji.fi (Accessed 20.10.2021).
GBIF: Global Biodiversity Information Facility. https://www.gbif.org/ (Accessed 20.10.2021).
GenBank: DQ467486.1. Draba hirta isolate L310. https://www.ncbi.nlm.nih.gov/nuccore/DQ467486.1
[Gosudarstvennaya…] Государственная геологическая карта Российской Федерации масштаба 1:1 000 000 (третье поколение). Балтийская сер.. Геологическая карта четвертичных образований. Лист Q-(35), 36. Апатиты. 2012. Сост: ОАО “МАГЭ”, ФГУП “ВСЕГЕИ”. СПб.
Grundt H.H., Popp M., Brochmann C., Oxelman B. 2004. Polyploid origins in a circumpolar complex in Draba (Brassicaceae) inferred from cloned nuclear DNA sequences and fingerprints. – Mol. Phylogenet. Evol. 32: 695–710.
Guardia R. 2021. CeDoc de Biodiversitat Vegetal: BCN-Cormophyta. CeDoc of Plant Biodiversity (CeDocBIV), Univ. Barcelona. Occurrence dataset accessed via GBIF.org on 2021-04-06https://doi.org/10.15468/x5ljvi
Gustafsson A.L.S., Skrede I., Rowe H.C., Gussarova G., Borgen L., Rieseberg L.H., Brochmann C., Parisod C. 2014. Genetics of Cryptic Speciation within an Arctic Mustard, Draba nivalis. – PLoS ONE. 9 (4): e93834. https://doi.org/10.1371/journal.pone.0093834
Hultén E. 1971. Atlas över växternas utbredning i Norden. 2nd ed. Stockholm. 531 p.
Hultén E., Fries M. 1986. Atlas of North European vascular plants. North of the Tropic of Cancer. Vol. I–III. Königstein.
Hyvärinen E., Juslén A., Kemppainen E., Uddström A., Liukko U.-M. (eds.). 2019. The 2019 Red List of Finnish Species. Helsinki. 704 p.
[Karta…] Карта четвертичных образований территории Российской Федерации, М 1:2 500 000. 2014. Министерство природных ресурсов и экологии РФ, Федеральное агентство по недропользованию, ФГУП “ВСЕГЕИ”, ФГУП “ВНИИОкеангеология”.
[Kiselev] Киселев И.И. 1979. Палеогеография западной части Кольского полуострова в неогене и антропогене. – Известия ВГО. 4: 316–324.
[Kostina, Borovichev] Костина В.А., Боровичев Е.А. 2014. Находки редких видов сосудистых растений в Мурманской области. – Труды Карельского научного центра РАН. 2: 155–159.
[Kozhin et al.] Кожин М.Н., Боровичев Е.А., Белкина О.А., Мелехин А.В., Костина В.А., Константинова Н.А. 2020. Редкие и охраняемые виды растений и лишайников памятников природы “Ущелье Айкуайвенчорр”, “Криптограммовое ущелье” и “Юкспорлак” (Мурманская область). – Труды Карельского научного центра РАН. 1: 34–48. https://doi.org/10.17076/bg939
[Kozhin et al.] Кожин М.Н., Боровичев Е.А., Белкина О.А., Мелехин А.В., Давыдов Д.А., Костина В.А., Константинова Н.А. 2019. К флоре памятников природы “Ущелье Айкуайвенчорр”, “Криптограммовое ущелье” и “Юкспоррлак” (Мурманская область). – Труды Карельского научного центра РАН. 8: 62–79. https://doi.org/10.17076/bg936
[Kravchenko et al.] Кравченко А.В., Кожин М.Н., Боровичев Е.А., Костина В.А. 2016. Новые данные о распространении редких видов сосудистых растений в Мурманской области. – Труды Карельского научного центра РАН. 3: 84–89.
[Kravchenko] Кравченко А.В. 2007. Конспект флоры Карелии. Петрозаводск. 403 с.
Kumar S., Stecher G., Li M., Knyaz C., Tamura K. 2018. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. – Mol. Biol. Evol. 35: 1547–1549.[Lutskina, Bochansteva] Луцкина Л.Ю., Бочанцева В.В. 2004. Редкий вид Севера Европейской части России – Draba insularis Pissjauk. (Brassicaceae Juss.). – Вестник СПбГУ. 3 (1): 99–100.
[Mishkin] Мишкин Б.А. 1953. Флора Хибинских гор, ее анализ и история. М.-Л. 112 с.
Mossberg B., Stenberg L. 2003. Den nya Nordiska Floran. Tangen. 928 p.
[Petrovskiy] Петровский В.В. 2018. Виды агрегата Draba cinerea (Brassicaceae) на севере России. – Новости систематики высших растений. 49: 71–86. https://doi.org/10.31111/novitates/2018.49.71
[Pis’yaukova] Письяукова В.В. 1956. Род Крупка – Draba L. – В кн.: Флора Мурманской области. Вып. III. М. С. 330–346.
[Ramenskaya] Раменская М.Л. 1983. Анализ флоры Мурманской области и Карелии. Л. 216 с.
[Ramenskaya, Andreeva] Раменская М.Л., Андреева В.Н. 1982. Определитель высших растений Мурманской области и Карелии. Л. 435 с.
[Red…] Красная книга Мурманской области. Изд. 2-е, перераб. и доп. 2014. Кемерово. 584 с.
[Red…] Красная книга республики Карелия. 2020. Белгород. 448 с.
Ridgway K.P., Duck J.M., Young J.P.W. 2003. Identification of roots from grass swards using PCR-RFLP and FFLP of the plastid trnL (UAA) intron. – BMC Ecol. 3(1): 8. https://doi.org/10.1186/1472-6785-3-8
Rognes T., Flouri T., Nichols B., Quince C., Mahé F. 2016. VSEARCH: A versatile open source tool for metagenomics. – Peer. J. 4: e2584. https://doi.org/10.7717/peerj.2584
Rønning O.I. 1979. Svalbards flora. 2nd edn. Oslo. 128 p.
[Tolmachev] Толмачев А.И. 1957. К истории развития и географического распространения рода Draba L. – Бот. журн. 42(9): 1446–1456.
[Tolmachev] Толмачев А.И. 1975. Род Draba L. – В кн.: Арктическая флора СССР. Т. 7. Л. С. 106–155.
[Tsinzerling] Цинзерлинг Ю.Д. 1935. Материалы по растительности северо-востока Кольского полуострова. М.–Л. 163 с.
Ulvinen T. 1996. Vascular plants of the former Kutsa Nature Reserve – Oulanka Reports. 16: 39–52.
[Vorob’yeva] Воробьева Е.Г. 1996. Флора островов в вершине Кандалакшского залива. – В кн.: Флора и растительность островов Белого и Баренцева морей. Мурманск. С. 57–89.
White T.J., Bruns T., Lee S., Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. – In PCR Protocols: A Guide to Methods and Applications. New York. P. 315–322.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал