

Ботанический журнал, 2023, T. 108, № 2, стр. 97-110
СТРУКТУРА МОХОВОГО ЭПИФИТНОГО ПОКРОВА ДЕРЕВЬЕВ СРЕДНЕТАЕЖНОГО ЕЛЬНИКА ЧЕРНИЧНОГО
Р. П. Обабко 1, 2, *, А. М. Крышень 1
1 Институт леса КарНЦ РАН
185910 Петрозаводск,
ул. Пушкинская, 11, Республика Карелия, Россия
2 Петрозаводский государственный университет
185910 Петрозаводск, пр. Ленина, 33, Республика Карелия, Россия
* E-mail: romaparrot@mail.ru
Поступила в редакцию 13.06.2022
После доработки 22.12.2022
Принята к публикации 17.01.2023
- EDN: EXJMOW
- DOI: 10.31857/S0006813623020084
Аннотация
В работе представлены результаты исследования пространственной структуры эпифитов на пробной площади и на стволе Picea abies. Распределение всех деревьев и отдельных пород на пробной площади, как и большинства видов эпифитных мохообразных не отличается от случайного типа. Только один вид Dicranum fuscescens, который в данном сообществе произрастал только на стволах живых деревьев, имеет агрегированное распределение. Все остальные эпифитные виды мохообразных встречены также и в напочвенном покрове и их распределение в сообществах относится к случайному типу. Предложена структура строения эпифитного покрова на стволе ели в условиях среднетаежного ельника черничного, которая состоит из различных видов, произрастающих полосами и пятнами вдоль градиента влажности.
Эпифитный покров бореальных лесов формируется лишайниками, мохообразными и в меньшей степени сосудистыми растениями. Роль его в устойчивости и динамике всего сообщества неочевидна. Его рассматривают, как индикатор состояния лесного сообщества (Shi et al., 2017 и др.). В то же время структура и динамика формируемых на стволах деревьев сообществ представляют интерес (Billings, Drew 1938; Cain, Sharp, 1938; Ashton, 1986; Isakova, 2009; Putna, Mežaka, 2014; Tarasova et al., 2017; Rykovskij и et al., 2020; Obabko, Tarasova, 2021 и др.). В бореальных лесах основная масса эпифитного покрова сосредоточена в нижней части ствола (Smith, 1982). В этой части ствола доминируют, как правило, факультативные эпифиты (Smith, 1982; Mežaka et al., 2008; Rykovskij et al., 2020). Комплекс внешних факторов обеспечивает сосуществование относительно большого числа видов, с одной стороны, и определяет сложную пространственную структуру эпифитного сообщества – с другой (Billings, Drew 1938; Barcman, 1958; Ashton, 1986; Ezer et al., 2019; Isakova, 2009 и др.).
Многие авторы, используя методы математического аппарата пространственной статистики в пакете spatstat (Baddeley, Turner, 2005; Baddeley et al., 2015), пришли к выводу, что итоговое расселение эпифитов в сообществе, которое для облигатных эпифитов часто может быть групповым, определяется не только биологическими особенностями видов и их взаимоотношениями (Snäll et al., 2005a,b; Akhavan et al., 2012; Sales et al., 2016; Tarasova, 2017 и др.), но и внешними факторами (Belinchón et al., 2011; Hedenås et al., 2003 и др.).
Главными факторами, определяющими структуру эпифитного покрова, являются те, которые напрямую или косвенно изменяют уровень увлажнения: видовая принадлежность дерева (Barkman, 1958; Putna, Mežaka, 2014), диаметр ствола (Gustafsson, Eriksson, 1995; Strazdina, 2010), угол наклона ствола, (Barkman, 1958; Øvstedal, 1980; Moe, Botnen, 1997; Gorshkov, 1986; Stepanova, 2004; Mežaka et al., 2010, 2012), экспозиция ствола (Sales et al., 2016, и др.).
В среднетаежных лесных экосистемах на территории Республики Карелии всего обнаружено 60 видов эпифитных мхов, из них только 7 считаются облигатными видами Lewinskya elegans (Schwägr. ex Hook. et Grev.) F. Lara, Garilleti et Goffinet, Neckera pennata Hedw., Nyholmiella obtusifolia (Brid.) Holmen et E. Warncke, N. gymnostoma (Bruch ex Brid.) Holmen et E. Warncke, Pylaisia polyantha (Hedw.) Bruch et al., P. selwynii Kindb., Ulota intermedia Schimp. (Obabko, Tarasova, 2021). Из факультативных эпифитов ряд видов имеют широкую экологическую амплитуду, являясь одновременно доминантами напочвенного и эпифитного покровов. К ним относятся, например, Pleurozium schreberi (Brid.) Mitt., Hylocomium splendens (Hedw.) Bruch et al., Rhytidiadelphus triquetrus (Hedw.) Warnst.
Мы поставили задачу исследовать пространственное распределение видов эпифитных мхов в ельнике черничном, как на уровне фитоценоза, так и отдельных стволов живых деревьев.
ОБЪЕКТЫ И МЕТОДЫ
Исследования проводились на постоянной пробной площади Института леса Карельского научного центра РАН, размером 25 × 55 м2, расположенной в ельнике черничном на территории заповедника “Кивач” (62.284° с.ш. 33.959° в.д.). Возраст насаждения: ель – 140–180 лет, береза – 40–60 лет. Породный состав насаждения 8E1Б1Ос. Было исследовано 50 деревьев, из которых 8 берез, 1 осина и 41 ель.
На пробной площади были закартированы все деревья (рис. 1) при помощи буссоли и ультразвукового дальномера DME Haglof. Для каждого дерева фиксировались: вид, высота, диаметр, угол наклона ствола с 4 сторон света. Угол наклона регистрировался при помощи угломера в телефоне Samsung galaxy a52.
Рис. 1.
Схема расположения деревьев на пробной площади. Цветом показаны породы: синим – Picea abies, зеленым – Betula sp., красным – Populus tremula.
Fig. 1. Location of tree species on the sample plot. The species are colored by blue – Picea abies, green – Betula sp., red – Populus tremula.

При описании эпифитного покрова каждого ствола мы фиксировали линейный размер куртин каждого вида в вертикальной проекции с четырех сторон света (4 описания на 1 дерево) и, как следствие, высоту поднятия эпифитных обрастаний по стволу (Degtyareva, 2012). Измерение линейного размера куртины мха происходило путем фиксации высоты над землей, где вид начинает встречаться (нижняя граница куртины) и где перестает встречаться (верхняя граница куртины) (рис. 2). Учитывались только сплошные обрастания, единичные растения, которые могут быть встречены на большей высоте ствола, не учитывались. Были собраны гербарные образцы мхов. Названия видов мхов даны по Ignatov et al. (2007) с некоторыми изменениями (Ignatov, Milyutina, 2007; Ignatov et al., 2018; Ignatova et al., 2019), названия печеночников по Söderström et al. (2016).
Рис. 2.
Линейные размеры эпифитных обрастаний стволов. На рисунке отмечены: черная вертикальная линия – трансекта, на которой производились измерения куртин. 1 – Ptilidium pulcherrimum, 2 – Lepraria spp., 3 – Cladonia spp., 4 – Dicranum sp.
Fig. 2. Linear dimensions of epiphytic canopy on trunks. Marked in the figure: black vertical line – the transect on which the measurements of canopy were made. 1 – Ptilidium pulcherrimum, 2 – Lepraria spp., 3 – Cladonia spp., 4 – Dicranum sp.

В качестве изучаемых характеристик эпифитного покрова были выбраны: число видов в каждом описании и линейные размеры куртин в вертикальной проекции наиболее часто встречающихся на стволах видов, а именно: Pleurozium schreberi, Hylocomium splendens, Plagiothecium rossicum Ignatov et Ignatova, Ptilidium pulcherrimum (Weber) Vain., Dicranum scoparium Hedw., D. fuscescens Turner, Cladonia spp.
СТАТИСТИЧЕСКИЕ МЕТОДЫ
Обработку полученных данных выполняли на основе стандартных методов вариационной статистики. Проверка нормальности распределения данных выполнена при помощи теста Колмогорова–Смирнова. Для всех анализируемых величин рассчитывали средние значения, выборки средних сравнивали друг с другом методом Краскела–Уоллиса. Попарное сравнение групп производилось методом Данна.
Характеристики (угол наклона ствола, диаметр дерева, линейный размер куртин доминантных видов), вдоль градиента которых изучались реакции эпифитных организмов, разбивали на 5 групп, с равным шагом характеристики. Для каждой группы рассчитывалось среднее значение и стандартная ошибка случайной переменной (линейный размер куртин отдельных видов). Число групп выбрано эмпирически на основании правила Стерджесса.
Для исследования пространственного распределения деревьев на пробной площади мы применили функцию J(r) реализованную в пакете spatstat (Baddeley et al., 2015). Данная функция была выбрана, так как она хорошо работает на локальном масштабе, что позволяет выявить агрегацию (Savel’ev et al., 2014). Функция J(r) стационарного точечного процесса определяется, как J(r) = (1–G(r))/(1–F(r)), где G(r) – функция распределения расстояний до ближайших соседей точечного процесса, а F(r) – функция пустого пространства, т.е. вероятность встречи точки от случайного пустого пространства.
Для полностью случайного (равномерного пуассоновского) точечного процесса J-функция равна 1 (красная пунктирная линия на графике, рис. 3 ). Отклонения J(r) < 1 или J(r) > 1 обычно указывают на пространственную кластеризацию или пространственную регулярность соответственно. Статистическая значимость работы функции определяется при помощи генераций конвертов Монте-Карло (Baddeley et al., 2015), и отображается на графике в виде серой области (в нашем случае число генераций равно 999, что соответствует уровню значимости p < 0.001). Выход функции (черная линия) из серой области, обозначает статистически значимые отличия наблюдаемого процесса от случайного.
РЕЗУЛЬТАТЫ
В эпифитном покрове нижней части стволов деревьев исследованного сообщества ельника черничного было выявлено 16 видов мхов, 4 вида печеночников и макролишайники, представленные родами Cladonia и Peltigera. Среди мохообразных только на ели встречено 8 видов. Видов, строго приуроченных к стволам березы и осины, в исследованном ельнике черничном не обнаружено (табл. 1). Меньшее число видов, обнаруженных на стволах лиственных деревьев, по всей видимости, связано с малым количеством исследованных деревьев. Все обнаруженные виды являются обычными для данной местности.
Таблица 1.
Видовой состав мохообразных на стволах живых деревьев Table 1. Species composition of bryophytes on the trunks of living trees
| Picea abies | Betula spp. | |||
|---|---|---|---|---|
| Минимальное, максимальное и среднее значение (± SE) ЛРК** на стволах деревьев, см. Minimum, maximum and mean value (± SE) of LDF** on a tree trunks, cm |
Встречаемость на стволе дерева ели, на ПП***, % Occurrence on a spruce tree trunk, on TP***, % |
Минимальное, максимальное и среднее значение (± SE) ЛРК** на стволах деревьев, см. Minimum, maximum and mean value (± SE) of LDF** on a tree trunks, cm |
Встречаемость на стволе дерева березы, на ПП***, % Occurrence on a birch tree trunk, on TP***, % |
|
| Lophocolea heterophylla (Schrad.) Dumort. | 0–5 (>0.01) | 4.9 | – | – |
| Dicranum fuscescens Turner | 0–53 (0.3 ± 0.12) | 19.5 | 0–46 (13.3 ± 4.49) | 100.0 |
| Dicranum majus Turner | 0–2 (>0.01) | 4.9 | – | – |
| Dicranum montanum Hedw. | 0–15 (0.1 ± 0.09) | 14.6 | – | – |
| Dicranum scoparium Hedw. | 0–26 (0.8 ± 0.3) | 39.0 | 0–12 (2.3 ± 1.81) | 25.0 |
| Hylocomium splendens (Hedw.) Bruch et al. | 0–56 (4.7 ± 1.16) | 36.6 | 0–46 (7.7 ± 3.13) | 75.0 |
| Plagiomnium cuspidatum (Hedw.) T.J. Kop. | 0–6 (0.1 ± 0.07) | 2.4 | 0–1 (>0.01) | 12.5 |
| Plagiochila sp. | 0–3 (>0.01) | 2.4 | – | – |
| Plagiothecium rossicum Ignatov et Ignatova | 0–35 (2.6 ± 0.63) | 70.7 | 0–9 (1.3 ± 0.89) | 25.0 |
| Pleurozium schreberi (Brid.) Mitt. | 0–90 (17.8 ± 2.09) | 82.9 | 0–74 (15.4 ± 5.08) | 87.5 |
| Pohlia nutans (Hedw.) Lindb. | 0–1 (>0.01) | 2.4 | – | – |
| Ptilidium pulcherrimum (Weber) Vain. | 0–29 (0.6 ± 0.19) | 39.0 | 0–37 (6.0 ± 2.61) | 87.5 |
| Rhytidiadelphus triquetrus (Hedw.) Warnst. | 0–22 (0.5 ± 0.36) | 4.8 | – | – |
| Rhodobryum roseum (Hedw.) Limpr. | 0–4 (0.1 ± 0.04) | 7.3 | 0–1 (>0.01) | 12.5 |
| Sanionia uncinata (Hedw.) Loeske | 0–1 (>0.01) | 4.8 | – | – |
| Sciuro-hypnum spp. (S. oedipodium, S. starkei, S. reflexum.)* | 0–24 (1.5 ± 0.44) | 63.4 | 0–9 (1.3 ± 1.11) | 25.0 |
| Tetraphis pellucida Hedw. | 0–1 (>0.01) | 4.8 | – | – |
Примечание: * Представители рода Sciuro-hypnum нередко встречались на одном дереве, и в полевых условиях не всегда возможно уверенно различить эти виды, в связи с этим в таблице приводятся данные для рода в целом. ** ЛРК – линейные размеры куртин. *** ПП – пробная площадь.
Note: * representatives of the genus Sciuro-hypnum were often found on the same tree, and in the field it is not always possible to confidently distinguish between these species, therefore, the table provides data for the genus as a whole. **LDF – Linear dimensions of foulings. *** TP – Trial plot.
В эпифитном покрове на разных породах рассматривались только мохообразные. Для ели число видов в описании варьировало от 0 до 5, для березы от 0 до 4. На пробной площади произрастало всего одно взрослое дерево осины, на котором отмечены 6 видов мохообразных.
ПРОСТРАНСТВЕННОЕ РАСПРЕДЕЛЕНИЕ В ФИТОЦЕНОЗЕ
Исследовано пространственное распределение деревьев на пробной площади – всего древостоя и только ели, отдельно для березы расчеты не проводились в связи с малым количеством особей на пробной площади. Для древостоя в целом не было обнаружено статистических отличий от случайного распределения (рис. 3А), ель в составе древостоя так же характеризуется случайным типом распределения (рис. 3В). При исследовании распределения на пробной площади деревьев с наиболее распространенными видами мохообразных отдельно для каждого вида (Plagiothecium rossicum, Dicranum fuscescens, Ptilidium pulcherrimum, Pleurozium schreberi, Hylocomium splendens) обнаружены значимые отличия от случайного типа распределения только для Dicranum fuscescens (p < 0.001). В случае D. fuscescens (рис. 3C) значение функции J(r) < 1, что говорит о наличии агрегации деревьев, на которых произрастает D. fuscescens. Схема распределения деревьев с D. fuscescens представлена на рис. 4.
Рис. 3.
J(r) – для всех пород (А), – для ели (В), – для Dicranum fuscescens (C). На рисунке черная линия – значение функции J(r), полученное на реальных данных, пунктирная линия – теоретическое значение J(r), соответствующее абсолютно случайному точечному процессу. Серая область – область принятия нулевой гипотезы о случайном распространении особей.
Fig. 3. J(r) – for all wood species (A), – for spruce (B), – for Dicranum fuscescens (C). In the figure the black line is the value of the function J(r) obtained on real data, the dotted line is the theoretical value of J(r) corresponding to an absolutely random point process. The grey area – the area of acceptance of the null hypothesis about the random distribution of individuals.

Рис. 4.
Схема расположения деревьев с Dicranum fuscescens в пределах пробной площади. На рисунке кружками большего диаметра отмечены деревья, на которых произрастает D. fuscescens, точками – деревья без D. fuscescens. Цветом выделены породы: синим – Picea abies, зеленым – Betula sp., красным – Populus tremula.
Fig. 4. Spatial distribution of trees with Dicranum fuscescens within the sample plot. In the figure circles of larger diameter indicate trees with D. fuscescens, dots indicate trees without D. fuscescens. The species are colored by: blue – Picea abies, green – Betula sp., red – Populus tremula.

УГОЛ НАКЛОНА СТВОЛА
Угол наклона ствола в значительной степени определяет увлажнение местообитания и физически влияет на возможность мха закрепиться на поверхности коры. Статистические закономерности обнаружены для линейных размеров куртин Pleurozium schreberi, которые имели минимальные значения при минимальном угле наклона ствола и по мере увеличения угла наклона ствола увеличивали свои показатели, достигая максимальных значений при угле наклона в 19° (рис. 5). Для остальных видов (Plagiothecium rossicum, Dicranum fuscescens, Ptilidium pulcherrimum, Cladonia spp.) не обнаружены статистические закономерности, но прослеживаются схожие тренды, что, скорее всего, связано с недостаточным объемом данных. Для разных видов оптимум угла наклона поверхности ствола отличается. Так, куртины Cladonia spp. достигают максимальных линейных размеров при средних значениях угла наклона около 9°, Dicranum fuscescens и D. scoparium – при 15°, Plagiothecium rossicum – в диапазоне от 3° до 9° и т.д. В то же время линейные размеры куртин Hylocomium splendens не показали значимой зависимости от угла наклона. Максимальное видовое разнообразие на стволах деревьев ели наблюдается при средних значениях угла наклона ствола – от 3° и до 9°.
Рис. 5.
Зависимость линейных размеров куртин Pleurozium schreberi от угла наклона ствола ели. Ось абсцисс – величина угла наклона, ось ординат – линейный размер куртин. Предел погрешности – стандартная ошибка. Буквами обозначены различия между средними (на основе результата теста Данна)
Fig. 5. Dependence of linear sizes of curtains Pleurozium schreberi from spruce trunk angle. Axis OX – the value of the angle of inclination, axis OY – the linear dimensions of the moss canopy. Margin of error – standard error. Letters indicate differences between means (based on Dunn’s test result)

ДИАМЕТР СТВОЛА
Диаметр ствола обычно связан с возрастом дерева, но в случае Picea abies эта связь непрямая, часто ель долгое время (до 100 лет) может быть угнетена материнским пологом. Возможно, поэтому из всех исследованных видов мхов только Hylocomium splendens и Dicranum fuscescens продемонстрировали связь линейных размеров куртин с диаметром ствола.
ВЗАИМООТНОШЕНИЯ ВИДОВ ВНУТРИ ЭПИФИТНОГО ПОКРОВА
Для исследования взаимоотношений видов эпифитного покрова нами в качестве независимых переменных были выбраны доминанты эпифитного покрова ели ‒ Pleurozium schreberi и Cladonia spp., которые, с одной стороны, имели максимальные линейные размеры куртин в вертикальной проекции, а с другой, – занимали в сообществах эпифитов крайние позиции по высоте ствола (рис. 5, табл. 1).
Методом сравнения средних был выявлен тренд, при котором линейный размер куртин Dicranum fuscescens (рис. 7А) и Plagiothecium rossicum (рис. 7B) сокращается при увеличении линейного размера куртин Pleurozium schreberi. Другие виды не показали зависимости от линейного размера куртин Pleurozium schreberi.
Рис. 6.
Изменение линейных размеров куртин Dicranum fuscescens (A) и Hylocomium splendens (B) при увеличении диаметра дерева ели. По оси абсцисс – диаметр ствола в см, по оси ординат – линейный размер куртин. Предел погрешности – стандартная ошибка. Буквами обозначены различия между средними (на основе результата теста Данна)
Fig. 6. Change in the linear dimensions of foulings of Dicranum fuscescens (A) and Hylocomium splendens (B) with a change in the diameter of a spruce tree. On the abscissa axis – the diameter of the trunk in cm, on the axis of the ordinate – the linear dimensions of the moss canopy. Margin of error – standard error. Letters indicate differences between means (based on Dunn’s test result)
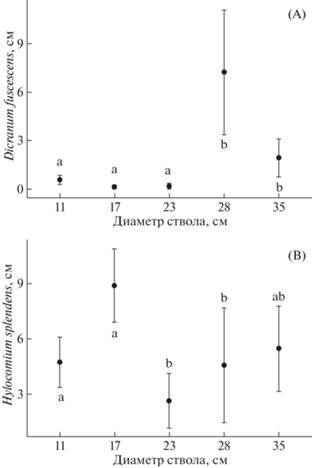
Рис. 7.
Изменения линейных размеров куртин Dicranum fuscescens (A) и Plagiothecium rossicum (B) при изменении линейных размеров куртин P. schreberi. По оси абсцисс – линейный размер куртин P. schreberi, по оси ординат – линейный размер куртин Dicranum fuscescens и Plagiothecium rossicum. Предел погрешности – стандартная ошибка. Буквами обозначены различия между средними (на основе результата теста Данна)
Fig. 7. Changes in the linear dimensions of canopy of Dicranum fuscescens (A) and Plagiothecium rossicum (B) with a change in the linear dimensions of canopy of P. schreberi. On the axis – canopy P. schreberi, on the axis – canopy Dicranum fuscescens and Plagiothecium rossicum. Margin of error – standard error. Letters indicate differences between means (based on Dunn’s test result)

При изучении зависимостей линейного размера куртин видов эпифитного покрова ели с линейным размером куртин видов рода Cladonia обнаружена положительная связь с Ptilidium pulcherrimum (рис. 8A) и Dicranum fuscescens (рис. 8B). Оба эти вида достигали максимальных линейных размеров куртин при максимальных размерах куртин Cladonia spp. Для остальных видов использованными методами взаимосвязи не обнаружены.
Рис. 8.
Изменения линейных размеров куртин Dicranum fuscescens (A) и Ptilidium pulcherrimum (B) при изменении линейных размеров куртин Cladonia spp. Предел погрешности – стандартная ошибка. Буквами обозначены различия между средними (на основе результата теста Данна)
Fig. 8. Changes in the linear dimensions of canopy of Dicranum fuscescens and Ptilidium pulcherrimum with a change in the linear dimensions of canopy of Cladonia spp. Margin of error – standard error. Letters indicate differences between means (based on Dunn’s test result)

ОБСУЖДЕНИЕ
В нашем исследовании основное внимание было уделено мхам, лишайники рассматривались только на уровне рода, в связи с этим данные по числу видов приводятся только для мохообразных. На ели и березе отмечалось примерно одинаковое число видов на отдельном дереве, для ели это 4.3 ± 0.21 SE, для березы 4.5 ± 0.42 SE. По сравнению с другими лесообразующими породами из района исследования береза и ель обладают низким видовым разнообразием эпифитов (Rykovskij et al., 2020). К примеру, в том же районе исследования на 1 дереве осины по нашим неопубликованным данным, может произрастать до 20 видов мохообразных. Низкое количество видов на ели и березе рядом авторов связывается с низким pH коры (Rykovskij et al., 2020), который у обеих пород варьирует в схожих пределах: для березы 3.2–5 (Coppins, 1984), для ели 3.5–5 (Kermit, Gauslaa, 2001).
Среди всех изученных видов групповое размещение в сообществе ельника черничного показал только Dicranum fuscescens, распределение других изученных видов значимо не отличается от случайного. Групповое размещение Dicranum fuscescens схоже с размещением облигатных эпифитов, для которых деревья являются островами в понятии Макартура (Patino et al., 2018), т.е. поверхность дерева является средой обитания эпифитов и окружена территорией, непригодной для их произрастания. Распределение видов на пробной площади, которое не отличается от случайного, можно объяснить тем фактом, что это факультативные эпифиты. Их экологическая амплитуда достаточно широка, что позволяет расти им на разнообразных субстратах: на почве, гниющей древесине и т.д. Для таких видов пространство для расселения ограничено только свободным местом и собственными конкурентными преимуществами, а не типом субстрата. Для облигатных видов ситуация иная. Их расселение ограничено, помимо свободного места и конкурентных связей, еще и наличием строго определенного субстрата. Пространственный паттерн таких видов имеет в большинстве случаев групповое размещение (Hedenås et al., 2003; Ignatenko et al., 2018). Несмотря на то что Dicranum fuscescens не является облигатным эпифитом и может произрастать на почве, в изученном сообществе он на почве не был обнаружен и при пространственном распространении демонстрировал групповое размещение.
УГОЛ НАКЛОНА ПОВЕРХНОСТИ СТВОЛА
Угол наклона поверхности влияет на перераспределение стока (Gorshkov, 1986; Stepanova, 2004; Mežaka et al., 2010, 2012), и, следовательно, определяет степень увлажнения поверхности ствола. Положительно наклоненные поверхности стволов получают в 2–3 раза больше осадков, чем отрицательно наклоненные (Barkman, 1958; Gorshkov, 1986). Так как местообитания эпифитов в целом относительно сухие, влага является определяющим фактором (Smith, 1982) и положительно наклоненные участки ствола наиболее благоприятны для произрастания эпифитов (рис. 9). При этом по нашим данным число видов достигает максимальных показателей при средних значениях угла наклона ствола от 5° до 12°. При дальнейшем увеличении угла наклона число видов снижается. Причина этого пока не ясна и требует специальных экспериментальных исследований. Здесь могут влиять и изменение физических параметров субстрата и взаимоотношения видов.
Рис. 9.
Эпифитный покров на стволе ели (слева положительный наклон ствола, справа отрицательный наклон ствола).
Fig. 9. Epiphytic cover on a spruce trunk (on the left a positive inclination of the trunk, on the right, a negative inclination of the trunk).

ДИАМЕТР СТВОЛА
Среди изученных видов связь с диаметром ствола выявлена только для Hylocomium splendens, причем место, занимаемое этим видом на стволе, снижается с увеличением диаметра дерева. Для других видов взаимосвязи не были обнаружены. Диаметр ствола дерева связан с размерами кроны, которая, увеличиваясь, интенсивнее отводит воду от ствола и одновременно снижает освещенность, что ухудшает условия произрастания Hylocomium splendens на стволе дерева, т.к. он отрицательно реагирует на затенение (Tarhova, Ipatov, 1975) и положительно – на увеличение увлажненности поверхности ствола (Bush, Abolin’, 1968).
В литературе имеются описания, как на стволе дерева может формироваться определенная структура эпифитного покрова, состоящая из последовательных полос различных видов (Billings, Drew 1938; Cain, Sharp, 1938; Ashton, 1986). Причина такого явления кроется в наличии градиента условий на стволе дерева и разных экологических оптимумов у разных видов, слагающих эпифитный покров. В общем виде градиент условий заключается в том, что увлажнение субстрата (коры) снижается с увеличением высоты над землей. С другой стороны, эпифитный покров имеет свою внутреннюю динамику, вызванную отмиранием нижней части куртины мха (лишайника), и отшелушивание коры дерева. Оба эти фактора разрушают контакт эпифита и коры дерева, и приводят к обвалу / выпадению части эпифитного покрова со ствола дерева (Ashton, 1986), обнажая участки коры, которые затем вновь заселяются мхами.
По совокупности наших наблюдений, полученных при проведении исследования, мы предположили структуру и функциональные связи в наиболее типичном эпифитном покрове крупного дерева ели, произрастающем в бореальном лесу в условиях Карелии. Общая схема строения эпифитного покрова на крупных елях дана на рис. 10.
Рис. 10.
Схема структуры эпифитного покрова ствола Picea abies. А – теоретическая схема эпифитного покрова; B – пример эпифитного покрова реального дерева. Обозначения в тексте.
Fig. 10. Scheme of the structure of the epiphytic cover of the trunk of the Picea abies. A – theoretical scheme of the epiphytic cover; B – an example of the epiphytic cover of a real tree. Explanation in the text.

На больших елях в достаточно увлажненных условиях эпифитный покров имеет общие черты, и его структура схематично представлена на рисунке 10А, пример реального дерева – на рисунке 10B. В нижней части отрицательно наклоненного отрезка ствола произрастает Plagiothecium spp. (10А, 1), в самом основании на положительно наклоненных участках разрастаются крупные виды, такие как Pleurozium schreberi, Hylocomium splendens и др. (10А, 2). Над ними, как правило, растут представители рода Dicranum (10А, 3), выше – лишайники родов Cladonia, Lepraria (10А, 4), над которыми может расти печеночный мох Ptilidium pulcherrimum (10А, 5). Как правило, на этом моховой покров заканчивается и выше P. pulcherrimum мохообразные не встречаются. Там (10А, 6) в зависимости от условий местообитания либо произрастают лишайники Coniocybaceae spp., Hypogymnia spp. и другие, либо эпифиты отсутствуют.
Pleurozium schreberi растет в нижней части ствола и окаймляет эпифитный покров снизу, максимально заполняя собой достаточно увлаженную область, возможную для произрастания мохообразных, тем самым вытесняя другие виды (рис. 8). Dicranum fuscescens и Plagiothecium rossicum снижают линейные размеры своих куртин с увеличением линейных размеров куртин Pleurozium schreberi. Данное местообитание отличается повышенной влажностью и низкой освещенностью, и пока P. schreberi имеет небольшие линейные размеры, факторы, влияющие на увеличение его обилия (влажность через угол наклона и т.д.), так же положительно сказываются и на Plagiothecium rossicum, который растет вслед за увеличением линейного размера куртин Pleurozium schreberi до значений 5–17 см. При дальнейшем увеличении линейных размеров P. schreberi Plagiothecium rossicum снижает свои показатели вплоть до полного отсутствия. В свою очередь, Cladonia spp. не оказывает такого воздействия на соседствующие виды. Линейные размеры Dicranum fuscescens и Plagiothecium rossicum увеличиваются при увеличении Cladonia spp., а линейные размеры Plagiothecium rossicum, который сильно разведен с Cladonia spp. (т.е. растут в разных зонах эпифитного покрова) в эпифитном покрове, никак не отреагировал на изменение линейного размера Cladonia spp. Возможный механизм, объясняющий взаимосвязь, заключается в том, что на стволе дерева есть градиент влажности, который зависит от определенных характеристик ствола. В случае, описанном Billings, Drew (1938), таким фактором была структура коры, которая с возрастом становится более грубой и толстой, что способствует лучшему удержанию влаги. В нашем случае для ели фактором, влияющим на условия увлажнения, является угол наклона ствола. Вдоль этого градиента распределяются эпифиты, но на определенной высоте снижение влажности воздуха (Molchanov, 1961; Ipatov, Kirikova, 1997) нивелирует положительное влияние факторов, повышающих увлажненность ствола. Подобный набор внешних воздействий, совместно с конкуренцией растений и приводит к видимой нами структуре, в которой крупный наиболее влаголюбивый мох, растущий внизу, наползает на выше растущие так же влаголюбивые виды, зажимая их между собой и сухой зоной, не пригодной для произрастания мхов. В то время как увеличение линейных размеров куртин засухоустойчивых, не способных к конкуренции со мхами, видов Cladonia spp., позитивно сказывается на линейных размерах куртин мхов, в связи с тем, что фактор (влажность), увеличивающий линейные размеры Cladonia spp., так же оказывает положительное влияние на мхи. Кроме того, лишайниковый покров в целом не столь плотный и позволяет сосуществовать видам.
Это схема “идеального” эпифитного покрова, встречающегося в условиях Южной Карелии. В реальности, данная структура подвержена многочисленным изменениям под влиянием внешних факторов. Так, снижение влажности (которое может быть вызвано в том числе повышенной инсоляцией в разреженных древостоях и т.д.) приводит к снижению видового состава и мощности/высоты поднятия эпифитов и линейных размеров в вертикальной проекции каждого вида в порядке их расположения на стволе. Первыми страдают крупные влаголюбивые виды, занимающие самую нижнюю часть ствола (на рис. 10 обозначено цифрой 2). При снижении влажности эти виды будут замещаться более мелкими видами рода Sciuro-hypnum, которые так же исчезнут при дальнейшем снижении влажности. За ними Dicranum spp. и Ptilidium pulcherrimum снижают свой линейный размер куртин. При этом Plagio-thecium spp. и Cladonia spp., которые в наименее развитых вариантах эпифитного покрова произрастают в непосредственном контакте на стволе, способны произрастать в условиях, не пригодных для других эпифитов.
ЗАКЛЮЧЕНИЕ
В условиях ельника черничного распределение большинства эпифитных видов мохообразных на уровне фитоценоза значимо не отличается от случайного типа распределения. Только Dicranum fuscescens имеет групповое распределение по пробной площади. Подобная пространственная закономерность характерна для облигатных эпифитов, расселяющихся с “материнского” дерева на ближайшие путем переноса диаспор. D. fuscescens, несмотря на то что способен произрастать на почве, в исследуемом нами сообществе не был встречен в мохово-лишайниковом ярусе.
При исследовании влияния диаметра дерева взаимосвязь обнаружена только для Hylocomium splendens, покрытие вида снижалось при увеличении диаметра дерева. Для других видов не обнаружено связи их линейного размера куртин с диаметром ствола. Возможно, это объясняется тем, что у ели нет прямой зависимости диаметра ствола от возраста дерева. Угол наклона ствола оказывал значимое положительное влияние на всем диапазоне своего изменения на линейные размеры куртин Pleurozium schreberi, в то время как виды р. Cladonia увеличивали свои показатели до значений угла наклона в диапазоне от 4 до 8 градусов. Дальнейшее увеличение угла наклона не влияло на линейные размеры куртины Cladonia, которые держались на уровне 12.5 см по высоте ствола. В целом, среди видов эпифитного покрова можно выделить группы, достигающие максимального развития при разных углах наклона. Так, Dicranum fuscescens, D. scoparium, Pleurozium schreberi, Ptilidium pulcherrimum достигают максимальных линейных размеров при больших углах наклона ствола (15 и более градусов), в то время как Cladonia spp., Plagiothecium rossicum при меньших (менее 15 градусов).
Результаты исследований подтвердили значимость внутренней структуры сообществ эпифитных организмов – статистическими методами подтверждено наличие взаимодействия растений различных видов внутри эпифитного покрова. Наибольшее число значимых связей обнаружено у Pleurozium schreberi и видов р. Cladonia.
В целом, структура растительного покрова отдельного живого дерева определяется одновременным влиянием субстрата (видовая принадлежность и параметры ствола) и взаимоотношением видов, слагающих сообщество эпифитных мохообразных и лишайников. Сообщество находится под одновременным влиянием множества внешних и внутренних факторов, определяющих его структуру, сходную для большинства описанных деревьев.
Список литературы
Akhavan R., Sagheb-Talebi Kh., Zenner E.K., Safavimanesh F. 2012. Spatial patterns in different forest development stages of an intact old-growth Oriental beech forest in the Caspian region of Iran. – European Journal of Forest Research. 131 (5): 1355–1366. https://doi.org/10.1007/s10342-012-0603-z
Ashton D.H. 1986. Ecology of bryophytic communities in mature Eucalyptus regnans F Muell forest at Wallaby Creek, Victoria. – Australian Journal of Botany. 34 (2): 107–129. https://doi.org/10.1071/BT9860107
Baddeley A., Rubak E., Turner R. 2015. Spatial point patterns: methodology and applications with R. CRC press. 828 p.
Baddeley A., Turner R. 2005. Spatstat: an R package for analyzing spatial point patterns. – Journal of Statistical Software. 2 (12): 1–42. https://doi.org/10.18637/jss.v012.i06
Barkman J.J. 1958. Phytosociology and Ecology of Cryptogamic Epiphytes. Van Gorcum, Assen. 628 p.
Belinchón R., Martínez I., Aragón G., Escudero A., De la Cruz M. 2011. Fine spatial pattern of an epiphytic lichen species is affected by habitat conditions in two forest types in the Iberian Mediterranean region. – Fungal Biology. 115 (12): 1270–1278. https://doi.org/10.1016/j.funbio.2011.09.003
Billings W.D., Drew W.B. 1938. Bark factors affecting the distribution of corticolous bryophytic communities. – American Midland Naturalist. 20 (2): 302–330.
[Bush, Abolin’] Буш К.К., Аболинь А.А. 1968. Строение и изменение растительного покрова важнейших типов леса под влиянием осушения. – В кн.: Вопросы гидролесомелиорации. Рига. С. 67–89.
Cain S.A., Sharp A.J. 1938. Bryophytic unions of certain forest types of the Great Smoky Mountains. – American Midland Naturalist. 20 (2): 249–301.
Coppins B.J. 1984. Epiphytes of birch. – Proceedings of the Royal Society of Edinburgh. Section B: Biological Sciences. 85 (1–2): 115–128.
[Degtyareva] Дегтярева С.И. 2012. Количественные учеты эпифитных группировок. – В кн.: Воспроизводство, мониторинг и охрана природных, природно-антропогенных и антропогенных ландшафтов. Воронеж. С. 51–56.
Ezer T., Alataş M., Batan N. 2019. Successional trends of some epiphytic bryophytes in Mediterranean Basin. – Acta Biologica Turcica. 32(4): 181–193.
[Gorshkov] Горшков В.В. 1986. Эпифитные лихеносинузии сосновых лесов Кольского полуострова (Формирование, экология, влияние антропогенных факторов): aвтореф. дис. канд. биол. наук. Л. 21 с.
Gustafsson L., Eriksson I. 1995. Factors of importance for the epiphytic vegetation of Aspen Populus tremula with special emphasis on bark chemistry and soil chemistry. – Journal of Applied Ecology. 32: 412–424. https://doi.org/10.2307/2405107
Hedenås H., Bolyukh V.O., Jonsson B.G. 2003. Spatial distribution of epiphytes on Populus tremula in relation to dispersal mode. – Journal of Vegetation Science. 14 (2): 233–242. https://doi.org/10.1111/j.1654-1103.2003.tb02148.x
[Ignatenko et al.] Игнатенко Р.В., Тарасова В.Н., Обабко Р.П. 2018. Пространственная структура ценопопуляций лишайника Lobaria pulmonaria в лесных сообществах Карелии. – В кн.: Труды XIV съезда РБО и конференции “Ботаника в современном мире” Т. 3. Махачкала. С. 105.
Ignatov M.S., Afonina O.M., Ignatova E.A., Abolina A., Akatova T.V., Baisheva E.Z., Bardunov L.V., Baryakina E.A., Belkina O.A., Bezgodov A.G., Boychuk M.A., Cherdantseva V.Ya., Czernyadjeva I.V., Doroshi-na G.Ya., Dyachenko A.P., Fedosov V.E., Goldberg I.L., Ivanova E.I., Jukoniene I., Kannukene L., Kazanovsky S.G., Kharzinov Z.Kh., Kurbatova L.E., Maksimov A.I., Mamatkulov U.K., Manakyan V.A., Maslovsky O.M., Napreenko M.G., Otnyukova T.N., Partyka L.Ya., Pisarenko O.Yu., Popova N.N., Rykovsky G.F., Tubanova D.Ya., Zheleznova G.V., Zolotov V.I. 2007. Check-list of mosses of East Europe and North Asia. – Arctoa. 15: 1–130. https://doi.org/10.15298/arctoa.15.01
[Ignatov, Milyutina] Игнатов М.С., Милютина И.А. 2007. Ревизия рода Sciuro-hypnum (Brachytheciaceae, Bryophyta) в России. – Arctoa. 16: 63–86. https://doi.org/10.15298/arctoa.16.07
[Ignatov et al.] Игнатов М.С., Игнатова Е.А., Федосов В.Э., Золотов В.И., Копонен Т., Чернядьева И.В., Дорошина Г.Я., Тубанова Д.Я., Белл Н.Э. 2018. Флора мхов России. Т. 4. Bartramiales – Aulacomniales. М. 543 с.
Ignatova E.A., Fedorova A.V., Kuznetsova O.I., Ignatov M.S. 2019. Taxonomy of the Plagiothecium laetum complex (Plagiotheciaceae, Bryophyta) in Russia. – Arctoa. 28: 28–45. https://doi.org/10.15298/arctoa.28.05
[Ipatov, Kirikova] Ипатов В.С., Кирикова Л.А. 1997. Фитоценология. СПб. 316 с.
[Isakova] Исакова Н.А. 2009. Видовое и синузиальное разнообразие листостебельных мхов восточного склона Ильменских гор. Ильмен. Екатеринбург: Миасс. 128 с.
Kermit T., Gauslaa Y. 2001. The vertical gradient of bark pH of twigs and macrolichens in a Picea abies canopy not affected by acid rain. – The Lichenologist. 33 (4): 353–359.
Mežaka A., Brūmelis G., Piterāns A. 2008. The distribution of epiphytic bryophyte and lichen species in relation to phorophyte characters in Latvian natural old-growth broad leaved forests. – Folia Cryptogamica Estonica. 44: 89–99.
Mežaka A., Brūmelis G., Piterāns A. 2010. Epiphytic bryophyte and lichen communities in relation to tree and forest stand variables in Populus tremula forests of south-east Latvia. – Acta Biol. Univ. Daugavp. 2: 1–8.
Mežaka A., Brūmelis G., Piterāns A. 2012. Tree and stand-scale factors affecting richness and composition of epiphytic bryophytes and lichens in deciduous woodland key habitats. – Biodiversity and Conservation. 21 (12): 3221–3241. https://doi.org/10.1007/s10531-012-0361-8
Moe B., Botnen A. 1997. A quantitative study of the epiphytic vegetation on pollarded trunks of Fraxinus excelsior at Havrå, Osterøy, western Norway. – Plant Ecol. 129(2): 157–177.
[Molchanov] Молчанов А.А. 1961. Лес и климат. M. 279 с.
[Obabko, Tarasova] Обабко Р.П., Тарасова В.Н. 2021. Эпифитная бриофлора Южной Карелии. – Труды Карельского научного центра РАН. 8: 41–49.
Øvstedal D.O. 1980. Lichen communities on Alnusn incana in north Norway. – Lichenologist. 12(2): 189–197.
Patino J., Gómez-Rodríguez C., Pupo-Correia A., Sequeira M., Vanderpoorten A. 2018. Trees as habitat islands: Temporal variation in alpha and beta diversity in epiphytic laurel forest bryophyte communities. – Journal of Biogeography. 45 (8): 1727–1738. https://doi.org/10.1111/jbi.13359
Putna S., Mežaka A. 2014. Preferences of epiphytic bryophytes for forest stand and substrate in North-East Latvia – Folia Cryptogamica Estonica. 51: 75–83. https://doi.org/10.12697/fce.2014.51.08
[Rykovskij] Рыковский Г.Ф., Малько М.С., Сакович А.А. 2020. Эпифитный компонент бриофлоры Полесского региона. – Природные ресурсы. 1: 49–57.
Sales K., Kerr L., Gardner J. 2016. Factors influencing epiphytic moss and lichen distribution within Killarney National Park. – Bioscience Horizons: The international journal of student research. 9. https://doi.org/10.1093/biohorizons/hzw008
[Savel’ev] Савельев А.А., Мухарамова С.С., Чижикова Н.А., Пилюгин А.Г. 2014. Теория пространственных точечных процессов в задачах экологии и природопользования (с применением пакета R). Казань. 146 с.
Shi X.M., Song L., Liu W.Y., Lu H.Z., Qi J.H., Li S., Chen X., Wu J.F., Liu S., Wu C.S. 2017. Epiphytic bryophytes as bio-indicators of atmospheric nitrogen deposition in a subtropical montane cloud forest: Response patterns, mechanism, and critical load – Environmental Pollution. 229: 932–941. https://doi.org/10.1016/j.envpol.2017.07.077
Smith A.J.E. 1982. Epiphytes and epiliths. – In: Bryophyte ecology. Springer. Dordrecht. P. 191–227.
Snäll T., Ehrlén J., Rydin H. 2005a. Colonization–extinction dynamics of an epiphyte metapopulation in a dynamic landscape. – Ecology. 86 (1): 106–115. https://doi.org/10.1890/04-0531
Snäll T., Pennanen J., Kivistö L., Hanski I. 2005b. Modelling epiphyte metapopulation dynamics in a dynamic forest landscape. – Oikos. 109 (2): 209–222. https://doi.org/10.1111/j.0030-1299.2005.13616.x
Söderström L., Hagborg A., von Konrat M., Bartholomew-Began S., Bell D., Briscoe L., Brown E., Cargill D.C., Costa D.P., Crandall-Stotler B.J., Cooper E.D., Dauphin G., Engel J.J., Feldberg K., Glenny D., Gradstein S.R., He X., Heinrichs J., Hentschel J., Ilkiu-Borges A.L., Katagiri T., Konstantinova N.A., Larraín J., Long D.G., Nebel M., Pócs T., Puche F., Reiner-Drehwald E., Renner M.A., Sass-Gyarmati A., Schäfer-Verwimp A., Moragues J.G., Stotler R.E., Sukkharak P., Thiers B.M., Uribe J., Váňa J., Villarreal J.C., Wigginton M., Zhang L., Zhu R.L. 2016. World checklist of hornworts and liverworts. – PhytoKeys. 27 (59): 1–828. https://doi.org/10.3897/phytokeys.59.6261
[Stepanova] Степанова В.И. 2004. Эпифитный лишайниковый покров ели европейской (Picea abies (L.) Karst) в еловых лесах Южной Карелии: Автореф. дис. … канд. биол. наук. СПб. 29 с.
Strazdina L. 2010. Bryophyte community composition on an island of Lake Cieceres, Latvia: dependence on forest stand and substrate properties. – Environmental and Experimental Biology. 8: 49–58.
Tarasova V.N., Obabko R.P., Himelbrant D.E., Boychuk M.A., Stepanchikova I.S., Borovichev E.A. 2017. Diversity and distribution of epiphytic lichens and bryophytes on aspen (Populus tremula) in the middle boreal forests of Republic of Karelia (Russia). – Folia Cryptogamica Estonica. 54: 125–141.
[Tarhova, Ipatov] Тархова Т.Н., Ипатов В.С. 1975. Влияние освещенности и опада на развитие некоторых мхов. – Экология. 1: 58–65.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал