

Физиология человека, 2019, T. 45, № 4, стр. 5-12
Эффекты повторения и рассогласования стимулов в сенсорных зрительных компонентах потенциалов, связанных с событиями
Ю. Д. Кропотов 1, *, В. А. Пономарев 1, М. В. Пронина 1, Н. В. Полякова 2
1 ФГБУН Институт мозга человека им. Н.П. Бехтеревой РАН
Санкт-Петербург, Россия
2 Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
* E-mail: yurykropotov@yahoo.com
Поступила в редакцию 26.11.2018
После доработки 19.12.2018
Принята к публикации 03.04.2019
Аннотация
Для исследования психических операций в условиях сенсорного повторения и рассогласования использовались три модификации парадигмы сравнения зрительных стимулов с образцом. В исследованиях участвовало 97 здоровых испытуемых, у которых анализировали 31-канальные потенциалы, связанные с событиями, в монтаже плотности источников тока. Показано, что повторение стимулов в активной, требующей ответной реакции, конструкции теста вызывает N250 компонент, локализованный в задних височных отведениях (T5, T6) и связанный предположительно с синаптическим привыканием. Напротив, рассогласование в физических характеристиках стимула вызывает положительный компонент P2, локализованный в тех же отведениях и связанный предположительно с возвратным приходом информации из высших отделов к низшим отделам коры. Сходные корреляты повторения/рассогласования наблюдаются и в пассивной, не треюущей ответной реакции, конструкции теста, однако амплитуда компонент в пассивной конструкции была ниже.
Один из важнейших сенсорных процессов заключается в сравнении нейронных следов сенсорного стимула в рабочей памяти с реально предъявляемым стимулом [1]. Для исследований этого процесса используется парадигма сравнения с образцом (A delayed match-to-sample task) и его модификации [2]. В этой парадигме стимулы предъявляются последовательными парами, в которых первый стимул служит в качестве образца (template), а второй может совпадать с первым (match) или не совпадать (mismatch) с ним. Задача испытуемого реагировать по-разному при совпадении и несовпадении стимулов, например, нажимать на различные кнопки. Если в ответ на несовпадение испытуемый должен воздержаться от ответа, парадигма превращается в двустимульную Go/NoGo парадигму (cued Go/NoGo paradigm) [3]. Потенциалы, связанные с событиями (ПСС – Event-related potentials – ERPs) являются адекватным методом для изучения психических процессов в общем аспекте, и процессов сенсорного рассогласования в частности [4, 5]. В обеих парадигмах условие рассогласования связывается с затылочно-височными положительными флуктуациями вызванных потенциалов [3, 6].
В предыдущих исследованиях мы использовали два метода слепого разделения источников: 1) метод независимых компонент (ICA) [7] и метод совместной диагонализации матриц ковариаций [8]. Оба метода на разностных кривых (несовпадение-совпадение) выявили положительный компонент с пиковой латенностью около 250 мс и локализацией в задних височных областях (T5/T6 в системе 10/20). Поскольку в статьях использовали разностные кривые (несовпадение-совпадение) вопрос о том, какая динамика потенциала связана с совпадением, а какая с несовпадением стимулов, оставался открытым. Кроме того, не был решен вопрос о влиянии активного, требующего ответной реакции, состояния человека на изучаемые мозговые корреляты в сравнении и пассивным восприятием сходных проб, не предполагающих ответной реакции испытуемых.
Для ответа на эти вопросы в данной работе мы модернизировали парадигму в виде трех различных модификаций теста так, чтобы иметь возможность исследовать различные парные условия: а) активное состояние в сравнении с пассивным состоянием, б) ситуацию, когда образ ожидаемого стимула находится в рабочей памяти, по сравнению с условиями, когда ожидаемый стимул не может быть предсказан. Поскольку в предыдущих работах мы достаточно точно локализовали источник искомой компоненты (электроды T5 и T6) в данной работе мы использовали монтаж плотности источников тока (Current Source Density – CSD) [9].
МЕТОДИКА
В исследованиях участвовали 97 здоровых испытуемых в возрасте от 18 до 55 лет (61 женщина). На экране монитора предъявляли две категории зрительных стимулов: по 20 различных изображений животных (Ж) и растений (Р). Изображения с угловыми размерами порядка 3.8° предъявляли в центре экрана на белом фоне. Между предъявлением изображений – белый фон. Предъявление изображений не сопровождали миганием экрана монитора. Для предъявления стимулов использовали программу Psytask.
Испытуемые выполняли три различные модификации базового теста (рис. 1). Тесты состояли из нескольких сотен проб. Число проб в тестах 1 и 2 – 400, а в тесте 3 – 600. Общая длительность тестов 1 и 2 – 21 мин, а теста 3 – 31 мин. В каждой пробе стимулы предъявляли парами. Длительность предъявления стимулов – 100 мс, интервал между стимулами в паре – 1000 мс, а интервал между началами пар стимулов – 3100 мс. Все пробы разделили на четыре равные группы, в зависимости от сочетания стимулов в паре. Использовали следующие сочетания пар стимулов: Ж–Ж, Ж–Р, Р–Р и Р–Ж. Пробы предъявляли в квазислучайном порядке.
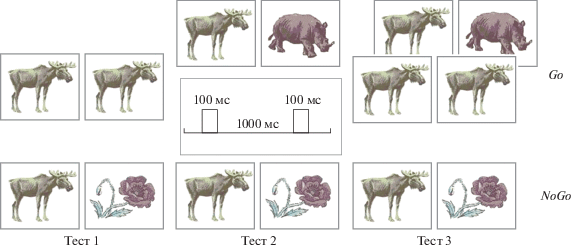
Тест 1. Физическое совпадение стимулов в парах Ж–Ж и Р–Р. В каждой паре стимулов Ж–Ж или Р–Р изображения животных или растений были одинаково физическими, например, изображение одного и того же медведя на первом и втором месте в паре Ж–Ж.
Тест 2. Физическое несовпадение стимулов в парах Ж–Ж и Р–Р. В каждой паре стимулов Ж–Ж или Р–Р изображения разных животных или растений.
Тест 3. Совпадение стимулов в половине пар Ж–Ж и Р–Р и несовпадение стимулов в других парах. В 50% пар стимулов Ж–Ж или Р–Р изображения животных или растений были одинаковыми, в остальных 50% пар стимулов Ж–Ж или Р–Р изображения животных или растений были разными.
Испытуемые нажимали на кнопку правой рукой как можно точнее и быстрее после предъявления пары изображений Ж–Ж. В тестах 1 и 2 нажимали левую кнопку. В тесте 3 на одинаковые пары нажимали левую кнопку, а на разные пары – правую кнопку. Пробы, в которых испытуемые неправильно выполняли задание, исключили из анализа. Перед началом теста испытуемый выполнял несколько десятков проб для тренировки. В ходе теста делали одну или несколько кратковременных пауз (длительностью 1–2 мин) для отдыха испытуемого.
ЭЭГ регистрировали с помощью компьютерного электроэнцефалографа “Мицар-ЭЭГ”. Электроды располагали в соответствии с модифицированной системой 10-10 в точках Fp1; Fpz; Fp2; F7; F3; Fz; F4; F8; FT7; FC3; FCz; FC4; FT8; T3; C3; Cz; C4; T4; TP7; CP3; CPz; CP4; TP8; T5; P3; Pz; P4; T6; O1; Oz; O2. Референт – объединенные электроды, расположенные на мочках ушей, заземляющий – в лобной области. Сопротивление электродов не превышало 5 кОм. Частота квантования сигналов – 250 Гц. Параметры ФВЧ и ФНЧ – 0.53 Гц (τ = 0.3 с) и 50 Гц соответственно. Для подавления сетевой электромагнитной помехи использовали цифровой режекторный фильтр с полосой непропускания 45–55 Гц.
Для обработки данных использовали программу WinEEG. Коррекцию артефактов моргания проводили путем обнуления соответствующих независимых компонент ЭЭГ [10]. Также из дальнейшего анализа исключали пробы, содержащие артефакты. Считали, что проба содержит артефакты, если в ней наблюдаются: 1) потенциалы более 50 мкВ для электродов Fp1 и Fp2 и более 100 мкВ для остальных электродов; 2) медленные волны в диапазоне от 0.53 до 1 Гц, с амплитудой более 50 мкВ; 3) быстрые колебания сигнала в интервале частот от 20 до 35 Гц, с амплитудой более 35 мкВ. Эти пороговые значения подбирали эмпирически, путем многократного выполнения обработки с различными параметрами и последующего визуального анализа результатов.
ЭЭГ, записанную по отношению к объединенным ушным референтам, преобразовывали в “плотность источников тока” (CSD) с помощью алгоритма, основанного на интерполяции распределения потенциалов сферическими сплайнами [11], и дающего адекватные результаты при анализе ЭЭГ, записанной с помощью относительно редкой сетки электродов [12].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Поведенческие показатели. Усредненные по группе испытуемых показатели выполнения тестов и их стандартные отклонения (SD) представлены в табл. 1. По данным табл. 1 видно, что число пропусков и ложных нажатий в трех тестах примерно одинаковое, однако время реакций в третьем тесте значительно превосходит таковое в первых двух.
Таблица 1.
Качество выполнения трех тестов
| Номер теста и условие |
Пропуски в % и стандартное отклонение (SD) |
Ложные нажатия в % и (SD) | Время реакции (ВР) в мс и (SD) | Стандартное отклонение ВР в мс и (SD) |
|---|---|---|---|---|
| Тест 1 | 4.5 (5.8) | 0.4 (0.6) | 360 (77) | 98 (33) |
| Тест 2 | 5.1 (7.5) | 0.6 (1.1) | 379 (74) | 93 (35) |
| Тест 3
Совпадение Несовпадение |
– 3.2 (4.5) 4.6 (5.3) |
0.4 (0.8) – – |
– 511 (82) 558 (80) |
– 120 (25) 114 (24) |
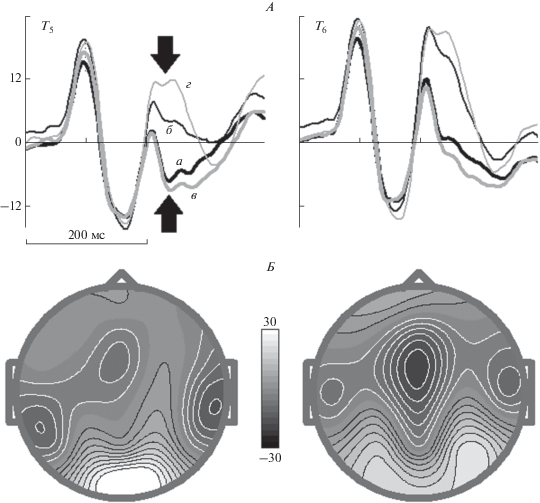
Go условие – совпадение и несовпадение. Рис. 2 представляет усредненную плотность источников тока для височных электродов (T5 и T6) для Go (Ж–Ж) стимулов во всех трех тестах. Отчетливо видно, что различия в динамике ответов на физически разные и одинаковые изображения животных наблюдаются спустя 200 мс после начала воторого стимула в паре. При физическом совпадении стимулов наблюдается отрицательное отклонение потенциала с минимумом около 250 мс слева – волна N250. При физическом несовпадении стимулов наблюдается положительное отклонение в интервале 200–300 мс – волна P200. Статистическая значимость различий среднего электрического потенциала в интервале 210–280 мс для Go условий в трех тестах, полученная с помощью двухфакторного дисперсионного анализа для повторных измерений с коррекцией Гринхауза−Гейсера (факторы: “условие” – 4 уровня, “локализация” – 2 уровня), следующая: основной эффект для фактора “условие” – F[3,288] = 151.0, ε = 0.77, p < 10–6, взаимодействие факторов – F[3,288] = 12.3, ε = 0.86, p < 10–5.
Рис. 2.
Усредненные по группе ПСС в ответ на Go условия в отведениях T5 и T6 (А) и топографии волн N250 (Б, слева) и P200 (Б, справа). По горизонтали – время, по вертикали – плотность источника тока в мкВ/м2. Стрелки указывают на соответствующие волны N250 и P200. а – тест 1, б – тест 2, в – тест 3, совпадение, г – тест 3, несовпадение.
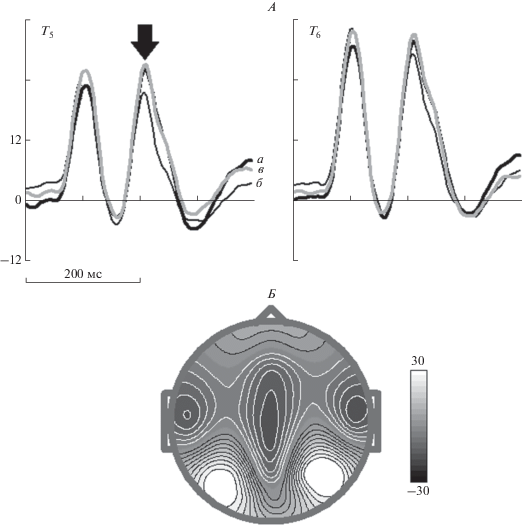
NoGo условие – несовпадение. Рис. 3 демонстрирует усредненные по группе вызванные потенциалы для трех NoGo условий (Ж–Р). Из рис. 3 видно, что NoGo условие сопровождается высоко-амплитудной P200 волной. Статистическая значимость различий среднего электрического потенциала в интервале 210–260 мс для NoGo условий в трех тестах следующая: основной эффект для фактора “условие” – F[2,192] = 28.4, ε = 0.96, p < 10–6, взаимодействие факторов – F[2,192] = = 3.3, ε = 0.99, p < 0.05.
Рис. 3.
Усредненные по группе ПСС в ответ на NoGo условия в отведениях T5 и T6 (А) и топография волны P200 (Б). Стрелка указывает на соответствующую волну P200. а – тест 1, б – тест 2, в – тест 3. Остальные обозначения см. рис. 2.
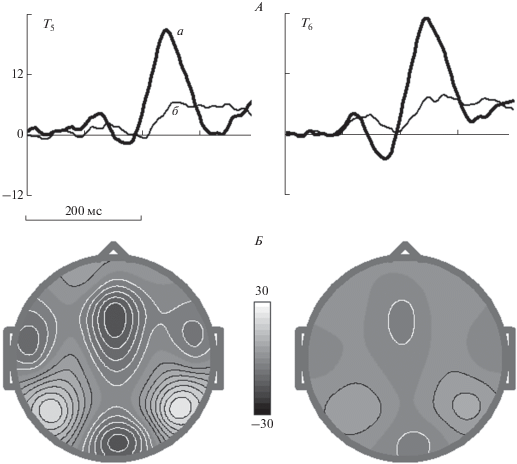
Активное по сравнению с пассивным. Для демонстрации влияния уловия (активное по сравнению с пассивным) были вычислены разностные кривые для третьего теста. Выбраны оба Go варианта (совпадения или несовпадения изображений животных) и оба условия, когда пары игнорируются (совпадения или несовпадения изображений растений). Отчетливо видно влияние условий – амплитуда разностной кривой в активном условии значительно превосходит амплитуду разностной кривой в пассивном условии. Статистическая значимость различий средней разницы электрического потенциала в интервале 210–280 мс для этих условий следующая: основной эффект для фактора “условие” – F[1,96] = 191.3, ε = 1.0, p < 10–6, взаимодействие факторов – незначимо (рис. 4).
Рис. 4.
Усредненные по группе ПСС для контраста разности несовпадение – совпадениие в 3 тесте) в активном условии (а) и пассивном условии (б) для электродов T5 и T6 (А) и топографии, определенные для максимальных значений в активном (слева) и пассивном условиях (Б). Остальные обозначения см. рис. 2.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Компонент N250 как индекс операции повторения. Первое описание влияния эффекта повторения на зрительную волну N250 было опубликовано в 2002 г. [13]. Авторы исследовали эффекты повторения на распознавание знакомых лиц с помощью регистрации ПСС. Никакого влияния повторений не наблюдалось для более раннего компонента N170, но наблюдалось значительная модуляция волны N250. Волна N250 была наибольшей при повторном предъявлении лиц людей, но наблюдалась и для других изображений (в частности, морд обезьян). В последующих исследованиях было доказано, что волна N250 не специфична к лицам, но скорее является показателем хранения внутренних образов в зрительной рабочей памяти [14]. Далее было показано, что волна N250 является показателем сходства зрительного стимула и образа в рабочей памяти: чем больше стимул похож на образ, тем выше N250 [15]. В этих исследованиях изображения лиц предъявляли парами, так что степень различия между стимулами в паре менялась от пробы к пробе. Предъявление стимула вызывало появление волн N170 и N250, причем сходство первого и второго стимулов влияло только на волну N250. Амплитуда N250 была наибольшей для проб, в которых второй стимул был похож на предыдущий, и постепенно снижалась как функция уменьшения подобия первого и второго стимулов.
Результаты данного исследования находятся в полном согласии с предыдущими работами. Действительно, в наших исследованиях повторение стимула в паре вызывает отчетливую волну N250, регистрируемую в плотности источников тока в задней височной области (электроды T5 и T6). Возможно, она отражает механизм привыкания, когда повторная активация корковых синапсов вызывает депрессию пост-синаптических потенциалов и растормаживание незадействованных синапсов.
Компонент P200 как индекс операции рассогласования. Во многих ситуациях детекция рассогласования жизненно необходима для адаптивного поведения. Эволюция создала несколько механизмов для детекции разных типов рассогласования. Один такой механизм связан со слуховой и зрительной негативностями рассогласования [16, 17]. Этот механизм отвечает за автоматическую детекцию рассогласования, когда нарушается регулярный паттерн зрительной или слуховой стимуляции. Другой механизм не является автоматическим, а требует когнитивного усилия для формирования в рабочей памяти модели ожидаемого стимула. Этот когнитивный механизм детекции рассогласования выражается в волне P2, распределенной в височной области (рис. 3).
Волна P2 в отличие от автоматических негативных волн рассогласования плохо описана в литературе. В контексте данной работы следует упомянуть исследования по ПСС-коррелятам зрительной маскировки, вызванной одновременным предъявлением стимула и маски (common onset masking) [18]. В этих исследованиях был обнаружен височно-затылочный компонент P2, амплитуда которого менялась в зависимости от степени маскировки. Авторы связывали этот компонент с повторной активацией зрительной коры, вызванной поступлением информации по нисходящим путям от высших зрительных областей к низшим областям. Схожая гипотеза была предложена в нашей книге на основе сравнительного анализа данных нейронной активности, локальных потенциалов и ПСС [19]. Согласно этой гипотезе переработка зрительной информации выполнялась двумя последовательными этапами, более поздний из которых связан с возвратным приходом информации из высших отделов, включая нижнюю префронтальную кору, к низшим отделам коры, включая височные области в составе вентрального зрительного пути. В рамках этой гипотезы мы можем предположить, что P2 компонент в ответ на рассогласование отражает повторную активацию зрительной коры, когда задействуются новые синапсы и тормозятся старые адаптированные синапсы.
ВЫВОДЫ
1. Нейроны задневисочных областей коры головного мозга обладают свойством привыкания к последовательному предъявлению одинаковых стимулов в активной парадигме теста, что отражается в величине компонента N250, локализованного в задних височных отведениях (T5, T6).
2. Рассогласование в физических характеристиках стимула вызывает положительный компонент P2, локализованный в задних височных отведениях (T5, T6) и связанный предположительно с возвратным приходом информации из высших отделов к низшим отделам коры.
3. Сходные корреляты повторения/рассогласования наблюдаются и в пассивной конструкции теста, однако амплитуда компонент в активной конструкции остается выше.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Комитетом по этике Институтом мозга человека им. Н.П. Бехтеревой РАН (Санкт-Петербург).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа поддержана РНФ (грант № 16-15-10 213).
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Rensink R.A. Change detection // Annual Review of Psychology. 2002. V. 53. P. 245.
Romo R., Salinas E. Flutter discrimination: neural codes, perception, memory and decision making // Nature Reviews Neuroscience. 2003. V. 4. № 3. P. 203.
Kropotov J.D., Ponomarev V.A. Decomposing N2 NOGO wave of event-related potentials into independent components // Neuroreport 2009. V. 20. P. 1592.
Kappenmann E.S., Luck S.J. ERP components: the ups and downs in brainwave recordings. The Oxford handbook of event-related potentials Oxford University Press, 2012. P. 3.
Kropotov J.D. Quantitative EEG, event-related potentials and neurotherapy. Academic Press, Elsevier, 2009. 542 p.
Kimura M., Katayama J., Murohashi H. Positive difference in ERPs reflects independent processing of visual changes // Psychophysiology. 2005. V. 42. № 4. P. 369.
Kropotov J.D., Ponomarev V.A., Hollup S., Mueller A. Dissociating action inhibition, conflict monitoring and sensory mismatch into independent components of event related potentials in GO/NOGO task // Neuroimage. 2011. V. 57. № 2. P. 565.
Kropotov J.D., Ponomarev V.A. Differentiation of neuronal operations in latent components of event-related potentials in delayed match-to-sample tasks // Psychophysiology. 2015. V. 52. № 6. P. 826.
Kayser J., Tenke C.E. On the benefits of using surface Laplacian (current source density) methodology in electrophysiology // International J. Psychophysiology. 2015. V. 97. № 3. P. 171.
Vigário R.N. Extraction of ocular artefacts from EEG using independent component analysis // Electroencephalography and Clinical Neurophysiology. 1997. V. 103. P. 395.
Perrin F., Pernier J., Bertrand O., Echallier J.F. Spherical splines for scalp potential and current density mapping // Electroencephalography and Clinical Neurophysiology. 1989. V. 72. P. 184.
Kayser J., Tenke C.E. Principal components analysis of Laplacian waveforms as a generic method for identifying ERP generator patterns: II. Adequacy of low – density estimates // Clinical Neurophysiology. 2006. V. 117. P. 369.
Schweinberger S.R., Pickering E.C., Jentzsch I. et al. Event-related brain potential evidence for a response of inferior temporal cortex to familiar face repetitions // Brain Research. Cognitive Brain Research. 2002. V. 14. № 3. P. 398.
Pierce L., Scott L.S., Boddington S. et al. The N250 brain potential to personally familiar and newly learned faces and objects // Frontiers in Neuroscience. 2011. V. 5. № 111. P. 1.
Sreenivasan K.K., Sambhara D., Jha A.P. Working memory templates are maintained as feature-specific perceptual codes // J. Neurophysiology. 2011. V. 106. P. 115.
Näätänen R., Gaillard A.W., Mantysalo S. Early selective-attention effect on evoked potential reinterpreted // Acta Psychologica. 1978. V. 42. № 4. P. 313.
Kimura M., Schröger E., Czigler I. Visual mismatch negativity and its importance in visual cognitive sciences // Neuroreport. 2011. V. 22. № 14. P. 669.
Kotsoni E., Csibra G., Mareschal D., Johnson M.H. Electrophysiological correlates of common-onset visual masking // Neuropsychologia. 2007. V. 45. № 10. P. 2285.
Kropotov J. Functional neuromarkers for psychiatry. 1st Edition. Applications for Diagnosis and Treatment. Academic Press, Elsevier, 2016. 498 p.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека