

Физиология человека, 2019, T. 45, № 6, стр. 86-102
Инфракрасная тепловизионная диагностика подкожных термогенераторов несократительного теплообразования
А. В. Козлов 1, *, В. Д. Сонькин 1, 2, **
1 Центр спортивных технологий и сборных команд Москомспорта
Москва, Россия
2 ФГБОУ ВО Российский государственный университет
физической культуры, спорта, молодежи и туризма
Москва, Россия
* E-mail: 89165363085a@mail.ru
** E-mail: sonkin@mail.ru
Поступила в редакцию 02.06.2019
После доработки 24.06.2019
Принята к публикации 29.06.2019
Аннотация
У 8 физически активных мужчин (возраст 24.8 ± 4.0 лет, индекс массы тела 23.6 ± 0.44) с помощью динамической инфракрасной термографии количественно описаны изменения кожной температуры различных участков поверхности тела (область шеи спереди и сзади, надключичные ямки, грудина и межлопаточная область) под действием факторов, вызывающих активацию несократительного термогенеза (НСТ). В ходе эксперимента испытуемые в разные дни подвергались глюкозотолерантному тесту, региональному холодовому воздействию (погружение стоп ног в воду 0°C на 1 мин), тесту на произвольную задержку дыхания, тестированию аэробной (рамптест) и анаэробной (вингейтский тест) производительности. В результате проведенных исследований было установлено, что в организме имеется несколько термогенераторов, способных к несократительному термогенезу в ответ на стимулы симпатического и стрессогенного характера. При этом, под термогенератором в данном контексте понимают расположенное подкожно или в более глубоких слоях тканей скопление однородных клеток, обладающих повышенной теплопродукцией, поток инфракрасного излучения которых достигает поверхности тела и формирует термопортрет. Выявлены глубокие индивидуальные различия в реакции термогенераторов на один и тот же набор стимулов. Эти реакции не отличаются ни синхронностью, ни соразмерностью и зависят, вероятно, от адаптивного опыта испытуемого, т.е. от условий его жизни и других эпигенетических факторов. Доказано, что наивысшей чувствительностью к испытанному набору стимулирующих воздействий обладают ассоциируемые с бурой жировой тканью (БЖТ) термогенераторы, расположенные в надключичной области. Между всеми исследованными термогенераторами обнаружены не вполне тривиальные взаимоотношения, которые нуждаются в дальнейшем изучении. В частности, с точки зрения термогенного ответа на воздействие, не выявлено сходства между максимальной аэробной и максимальной анаэробной физической нагрузкой. Реакция на глюкозу оказалась изолированной по отношению к другим стимулам. На основании полученных данных необходимо задуматься о роли не только БЖТ, но и других тканей в регуляции энергетического метаболизма. В частности, следует обратить внимание на мускулатуру, располагающую разобщающим белком UCP3. Кроме того не следует однозначно понимать природу исследованных термогенераторов. Но, возможно, широкое применение неинвазивного и безопасного тепловизора позволит накопить научные факты, необходимые для дифференциальной диагностики различных видов термогенераторов в организме человека.
Инфракрасный тепловизор (ИК) – неинвазивная и абсолютно безвредная для человека техника, позволяющая наблюдать тепловые поля на поверхности тела (термопортрет), несущие информацию о состоянии энергометаболических процессов в организме [1]. Однако для включения в научный оборот эта информация должна быть расшифрована, что представляет собой не вполне тривиальную задачу биомедицинской диагностики.
В организме теплокровных существуют три вида термогенеза:
1) сократительный термогенез (мышечная работа или холодовая дрожь, в той или иной форме имеется у всех позвоночных);
2) несократительный термогенез (например, кратковременная активация бурой жировой ткани в ответ на умеренное охлаждение, – только у плацентарных млекопитающих);
3) постсократительный термогенез (восстановление после выполненной тяжелой физической нагрузки, не сопровождающееся сократительной активностью мышц – у всех позвоночных).
Термопортрет человека, получаемый с помощью инфракрасного тепловизора – сложный конструкт, объединяющий морфологические и функциональные характеристики изучаемого объекта (человека как излучателя тепла) [2]. При этом термопортрет, полученный в стандартных условиях, отражает, в первую очередь, устойчивую морфологическую картину, модулированную изменяющимся функциональным состоянием [3–5]. Фактически термопортрет отражает активность набора термогенераторов, участвующих в его формировании. При этом под термогенератором в данном контексте мы понимаем расположенное подкожно или в более глубоких слоях тканей скопление клеток, обладающих повышенной теплопродукцией, поток инфракрасного излучения которых достигает поверхности тела и формирует термопортрет.
Важно отметить, что лонгитудинальные наблюдения подтверждают устойчивость и консервативность термопортрета, зафиксированного в одном и том же функциональном состоянии [6–8]. При этом индивидуальное разнообразие термопортретов огромно [7]. Сочетание этих двух характеристик – доказательство того, что конкретный термопортрет определяется морфологическими (анатомическими) индивидуальными особенностями данного человека и не подвержен быстрым функциональным флуктуациям при отсутствии соответствующих стимулов.
Соответствующими стимулами в данном случае могут являться:
1) охлаждающее воздействие, достаточное для активации несократительного термогенеза (симпатическая стимуляция через кожные терморецепторы) [9],
2) прием пищевых веществ, вызывающих калоригенное действие (субстратная стимуляция) [10],
3) неспецифическое стрессовое воздействие (симпатическая и/или гормональная стимуляция) [11],
4) напряженная физическая нагрузка и постнагрузочное восстановление (постсократительный термогенез, сопряженный с процессами восстановления) (стимуляция по всем каналам регуляции гомеостаза) [12],
5) воспаление [2] (в данном контексте не рассматривается).
Различного размера и формы изотемпературные зоны термопортрета, как правило, могут быть ассоциированы с конкретными анатомическими структурами, обладающими в той или иной степени термогенной активностью (в том числе бурая жировая ткань и ее аналоги), либо являющимися транспортерами тепла от мест его образования к поверхности (кровеносные сосуды) [13]. Поскольку ИК тепловизор фиксирует поверхностную температуру, постольку ИК термопортрет представляет собой мгновенный интеграл теплообразования, теплопроводности и рассеивания тепла, наблюдаемый как проекция термически активных структур, расположенных на разной глубине, на поверхность тела (кожный покров), которая, в свою очередь, охлаждается за счет испарения пота [14]. По этой причине предпочтительно динамическое наблюдение за изменениями термопортрета в различных модельных ситуациях [3].
Имеются три обстоятельства, сильно усложняющие анализ инфракрасных термограмм.
Первое обстоятельство связано с уже упоминавшимся выше огромным индивидуальным разнообразием термопортретов, устойчивых во времени, что подтверждает тезис об анатомической причине неравномерного распределения температур по поверхности тела человека [7]. При этом реальный размер и глубина залегания термогенной ткани остаются неизвестными [15].
Второе обстоятельство связано с существованием как минимум двух разновидностей жировой ткани с большим количеством митохондрий и наличием в них разобщающего белка UCP1 (термогенин): “классический” бурый жир, характерный для новорожденных, но сохраняющийся у значительной части взрослых [16], а также “бежевый жир”, представляющий собой “побуревший” под воздействием разных факторов белый жир [17]. Уже известны многие детали сходства и различия этих тканей [18], включая их происхождение от разных прогениторных клеток [19], однако полной ясности в их специфике нет [20, 21]. Будут ли они одинаково или по-разному реагировать на стимулирующие воздействия различных модальностей – не очевидно.
Третье обстоятельство ставит под сомнение позитронно-эмиссионную томографию (ПЭТ) как лучший метод выявления активного бурого жира. Дело в том, что для диагностики БЖТ с помощью ПЭТ используется меченая глюкоза, по интенсивности захвата которой и определяется активность БЖТ на скане ПЭТ [22]. Однако известно, что классическая БЖТ активно использует также жирные кислоты [23, 24] – на чем, собственно, основаны перспективы использования БЖТ и ее аналогов в борьбе с метаболическим синдромом. При этом не только БЖТ, но и мышцы, содержащие разобщающие белки семейства UCP, активно окисляют жирные кислоты [25]. С другой стороны, гликолитические и липолитические процессы конкурируют между собой и не происходят одновременно в одной и той же клетке [26, 27]. При регистрации с помощью ПЭТ локусов, активно захватывающие меченую глюкозу, видна, вероятно, только часть клеточной популяции БЖТ, так как для другой части адекватным субстратом в это время будут жирные кислоты. Есть данные, что БЖТ способна утилизировать не только глюкозу и жирные кислоты, но и другие метаболиты – в частности, молочную кислоту [28, 29], в обилии выделяющуюся при мышечной нагрузке. Это еще один вид метаболической активности БЖТ, который пока детально не исследован.
Из сказанного следует, что данные ПЭТ вовсе не являются абсолютно убедительным поводом в отношении активности несократительного термогенеза в организме человека. Безопасный и дешевый тепловизор, напрямую измеряющий тепловое состояние поверхности тела, может с успехом конкурировать с дорогостоящим и опасным методом, основанным на использовании радиоактивных изотопов [6, 7, 30, 31].
Однако для адекватного использования тепловизора при изучении несократительного и постсократительного термогенеза требуется разработка методологии таких измерений и интерпретации получаемых данных [30]. Анализ инфракрасного термопортрета человека и его изменений под воздействием функциональных проб, способных активировать различные формы несократительного термогенеза, позволяет выявлять подкожные термогенераторы разных типов, одни из которых сильнее активируются субстратными стимулами (глюкоза), другие – холодовыми (острое локальное охлаждение конечностей) или другими стрессогенами (задержка дыхания), а третьи активно участвуют в восстановительных процессах после тяжелой физической нагрузки (постсократительный термогенез). Часть из этих термогенераторов, несомненно, представляет собой скопления БЖТ, тогда как другие могут быть сформированы способной к несократительному термогенезу мускулатурой либо представлять собой адаптивно измененные клетки подкожной жировой клетчатки (т.н. “бежевая” жировая ткань).
Целью исследования является выявление локализации подкожных термогенных структур (термогенераторов), осуществляющих несократительный термогенез, и сопоставление их активации в ответ на разные функциональные стимулы.
МЕТОДИКА
Для ответа на поставленные вопросы провели исследование динамических изменений термопортрета при воздействии факторов разной модальности, способных стимулировать термогенез. Для этого вели непрерывную регистрацию температуры определенных участков кожи испытуемых с помощью калиброванных тепловизоров марки NEC. При этом в центре интереса авторов находилась область несократительного и постсократительного термогенеза, поскольку источники этих физиологических реакций менее очевидны, чем источник сократительного термогенеза, каковым являются скелетные мышцы и сердце.
Экспериментальную работу проводили на базе Центра спортивных технологий и сборных команд Москомспорта. В исследованиях принимали участие 8 взрослых здоровых мужчин (средний возраст 24.8 ± 4.0 г., масса тела 74.9 ± 5.1 кг, длина тела 178.3 ± 4.8 см, ИМТ 23.6 ± 0.44). Все испытуемые воздерживались от приема пищи в течение 2 ч и от приема алкоголя не менее чем 24 ч до каждого тестирования. Каждого испытуемого подвергали в разные дни последовательно воздействию одинаковых факторов, которые, по крайней мере у части испытуемых, вызывают активацию факультативного несократительного термогенеза, а именно: холодовая проба (охлаждение стоп ног в воде при температуре около 0° в течение 1 мин); гипоксическая проба (задержка дыхания на спокойном выдохе до отказа); глюкозо-толерантный тест (введение глюкозы натощак перорально в количестве 1 г/кг при 3-часовом наблюдении); тест аэробной производительности (рамптест на беговой дорожке (HP Cosmos) до достижения максимального потребления кислорода (МПК) (рассматривали восстановительный период); тест анаэробной производительности (вингейтский 30-секундный анаэробный тест на велоэргометре (Lode) (рассматривали восстановительный период)). Каждого испытуемого подвергали тестированию с применением различных воздействий с интервалом 3–4 дня. За сутки до тестирования физическую нагрузку испытуемые не выполняли. Подробное обоснование выбора воздействий и описание протоколов тестовых проб представили в предыдущей публикации [30]. Общая характеристика группы испытуемых представлена в табл. 1.
Таблица 1.
Основные характеристики группы испытуемых и результаты эргометрии
| n = 8 | Показатель | Обозначение | Размерность | Среднее значение |
| Возраст | – | лет | 24.8 ± 4.0 | |
| Длина тела | ДТ | см | 178 ± 4.8 | |
| Масса тела | МТ | кг | 74.9 ± 5.1 | |
| Вингейтский анаэробный тест | Максимальная мощность, показанная в тесте | W max | Вт | 769 ± 61.3 |
| Максимальная относительная мощность, показанная в тесте | W max/кг | Вт/кг | 10.2 ± 0.52 | |
| Время достижения максимальной мощности | t W max | с | 6.41 ± 0.81 | |
| Рамптест | Время работы в рамптесте | t max | мин | 16.51 ± 1.4 |
| Максимальная достигнутая скорость бега | V max | км/ч | 16.4 ± 0.8 | |
| Максимальное потребление кислорода в тесте | VO2max | л/мин | 3.761 ± 0.4 | |
| Максимальное относительное потребление кислорода в тесте | VO2max/кг | мл/мин/кг | 50.38 ± 3.73 | |
| Скорость бега на уровне анаэробного порога | АнП V | км/ч | 13.6 ± 0.81 | |
| Потребление кислорода в % относительно максимального на уровне анаэробного порога | %VO2 АнП | % | 86 ± 4.5 |
Перед каждым тестовым воздействием испытуемые проходили температурную адаптацию в течение 15 мин в изолированном помещении с температурой 21–22°С (во время глюкозотолерантного теста 28–30°С) и влажностью 45–50% в состоянии мышечного покоя, раздетые по пояс, в положении сидя. Эта схема проведения исследования была призвана обеспечить умеренную активацию механизмов химической терморегуляции. Во время 15-минутной экспозиции наряду с другими измерительными процедурами с помощью двух бесконтактных тепловизоров NEC TH 9100SL проводили тепловизионную съемку спины и груди (одновременно). Далее испытуемого подвергали одному из тестовых воздействий (см. выше), после чего тепловизионную съемку продолжали согласно протоколу исследования. Во всех тестах, кроме глюкозотолерантного, регистрация температуры и другие измерительные процедуры после воздействия продолжались 10 мин. Во время глюкозотолерантного теста после приема раствора глюкозы съемка и другие измерительные процедуры продолжались 180 мин. Тепловизор располагали на расстоянии 3 м от испытуемого и на высоте 140 см от пола. Полученные термограммы обрабатывали с помощью специализированной программы Image Processor. Калибровку тепловизора осуществляли в соответствии с “Правилами эксплуатации” перед каждым измерением.
Регистрацию температуры спины или груди проводили со скоростью 4 кадра в секунду. Во всех случаях, кроме глюкозотолерантного теста, лактат и глюкозу в периферической крови измеряли до воздействия, по завершении воздействия и через 1 мин после его завершения, далее на 1, 2, 4, 6, 8 и 10 мин. Во время глюкозотолерантного теста, который длился в общей сложности 195 мин, содержание лактата и глюкозы в капиллярной крови измеряли до приема глюкозы и далее каждые 20 мин. Концентрации лактата и глюкозы определяли автоматическим анализатором Biosen C-Line (EKF-diagnostic GmbH., Германия). Протоколы всех 5 воздействий на испытуемого более подробно описаны в предыдущей статье [30].
При выборе регионов для наблюдения (рис. 1) мы в первую очередь ориентировались на данные ряда лабораторий мира, где используется инфракрасное тепловидение для выявления активного бурого жира [31–34], а также на анатомические данные [35]. Показано, что результаты термографирования хорошо согласуются с результатами ПЭТ в области надключичных ямок, где БЖТ расположена поблизости от поверхности тела [31]. По этой причине регион “ключицы” был первым при выборе локусов, в которых мы предполагали фиксировать термогенную активность.
Рис. 1.
Области наблюдения поверхности спины и груди для мониторирования динамических изменений теплового состояния кожного покрова. Области 1 и 2 включают всю измеряемую поверхность со стороны спины и груди, соответственно. Область 3 – шея со стороны спины (в таблицах и диаграммах – шея/сп), область 4 – между лопаток, область 5 – шея со стороны груди (шея/гр), область 6 – ключицы, область 7 – грудина (тепловая проекция сердца), область 8 – проекция щитовидной железы (щитов. ж.).

Наряду с этим регионом включили в исследование также регионы, наблюдаемые при фронтальной съемке: шея со стороны груди, где некоторыми авторами отмечается присутствие БЖТ у взрослых людей [36, 37]; проекция щитовидной железы – поскольку эта железа непосредственно участвует в регуляции энергетического метаболизма и теплопродукции БЖТ [38]; грудина – поскольку здесь может иметь место тепловая проекция активно работающего сердца. Кроме того, включили два локуса, наблюдаемые только при дорсальной съемке: шея со стороны спины; межлопаточная область, в которой локализуется БЖТ у детей с момента рождения, однако и у взрослых могут сохраняться фрагменты активной ткани [39, 40].
Таким образом, мы охватили большую часть потенциальных подкожных локализаций БЖТ в организме взрослого человека. При этом не было возможности зафиксировать реакцию глубоко расположенных фрагментов БЖТ: внутри грудной клетки, в области перикарда, в области почек, в паховых и подмышечных областях – инфракрасный сигнал от этих скоплений БЖТ поглощается окружающими тканями и практически не достигает поверхности тела (по крайней мере, его невозможно выделить среди инфракрасных сигналов других тканей).
Для определения возможной локализации подкожных термогенных структур человека и их реакции на стимулы разной модальности необходима количественная оценка. Мы использовали расчетный показатель – “индекс термогенности” (IT), который является мерой оценки изменений теплового состояния кожи во времени и учитывает динамику температуры поверхности тела в процессе длительных наблюдений [30].
IТ представляет собой отношение разности максимальной температуры определенной области и средней температуры всей измеряемой поверхности кожи в определенный момент времени tk к разности этих температур сразу после температурной адаптации, т.е. до стимулирующего термогенез воздействия на испытуемого в момент времени t0:
(1)
$IT\left( {i,t} \right) = {{{{{\left( {Ti--Tav} \right)}}_{{{{t}_{k}}}}}} \mathord{\left/ {\vphantom {{{{{\left( {Ti--Tav} \right)}}_{{{{t}_{k}}}}}} {{{{\left( {Ti--Tav} \right)}}_{{{{t}_{0}}}}},}}} \right. \kern-0em} {{{{\left( {Ti--Tav} \right)}}_{{{{t}_{0}}}}},}}$Оценка теплового состояния тех или иных областей поверхности тела при помощи IT позволяет количественно определять изменение уровня термоактивации локально расположенных тканей относительно исходного состояния испытуемого.
При помощи индекса IT становится возможным не только оценивать тепловое состояние различных областей поверхности тела, количественно определяя и анализируя изменение уровня их термоактивности при воздействии разной модальности, но и сравнивать уровень термоактивности гомологичных участков у разных испытуемых.
По динамическим рядам данных термографирования для каждого из испытуемых рассчитывали величины IT, из которых для статистической обработки и построения диаграмм учитывали максимальные значения для каждой выделенной области во время стимулирующего термогенез воздействия на испытуемого.
Анализ полученных результатов проводили с использованием статистических методов, реализованных в MS Excel, кроме того, применяли ранговую корреляцию Спирмена для анализа взаимосвязей между характеристиками термогенного ответа различных локусов.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Для корректного использования тепловизора при изучении несократительного и постсократительного термогенеза, требуется разработка методологии таких измерений и интерпретации получаемых данных. В первую очередь, потребовалось разработать инструмент для изучения термогенной активности подкожных структур, позволяющий оценивать относительную интенсивность теплового потока IT в области наблюдения [30]. Соответствующие результаты для всех 5 модальностей стимулов и для 6 локусов наблюдения представлены на рис. 2.
Рис. 2.
Диаграмма, построенная по величинам IT в 6 наблюдаемых локусах при 5 модальностях стимула (среднегрупповые значения). Шкала в ед. IT. На диаграмме и далее обозначения применяемых к испытуемым воздействий: глюкоза – глюкозотолерантный тест, задержка дыхания, холод – холодовое воздействие, вингейт – вингейтский анаэробный тест.

Из данных рис. 2 видно, что самая высокая интенсивность термогенеза регистрируется на поверхности тела в восстановительном периоде после рамптеста до отказа. Особенно высокая теплопродукция характерна для области грудины (проекция сердца) и поверхности шеи со стороны спины, чуть меньше – со стороны груди. Паттерн термогенного ответа на все другие стимулирующие воздействия занимает меньшую площадь, а следовательно, менее интенсивен, и демонстрирует однотипность формы – вне зависимости от характера стимула.
Корреляционный анализ (табл. 2 и 3) показал, что достоверные взаимосвязи по показателю IT наблюдаются между локусом “шея спины” и остальными локусами за исключением ключиц, где наиболее вероятно присутствие БЖТ, и проекции щитовидной железы, тесно связанной с ключицами.
Таблица 2.
Корреляции между локусами несократительного термогенеза по IT
| n = 40, r ≥ 0.31 | Шея/сп | |||||
|---|---|---|---|---|---|---|
| Шея/сп | 1.000 | Между лопаток | ||||
| Лопатки | 0.410** | 1.000 | Шея/гр | |||
| Шея/гр | 0.366* | 0.555** | 1.000 | Ключицы | ||
| Ключицы | 0.132 | 0.277 | 0.512** | 1.000 | Грудина | |
| Грудина | 0.414** | 0.505** | 0.606** | 0.386* | 1.000 | Щитов. ж. |
| Щитов. ж. | 0.233 | 0.541** | 0.687** | 0.516** | 0.387* | 1.000 |
Таблица 3.
Корреляции между стимулами несократительного термогенеза по IT
| n = 48, r ≥ 0.28 | Глюкоза | ||||
|---|---|---|---|---|---|
| Глюкоза | 1.000 | Задержка | |||
| Задержка | 0.202 | 1.000 | Холод | ||
| Холод | 0.330* | 0.363* | 1.000 | Рамп/восст | |
| Рамп/восст | 0.091 | 0.537** | 0.262 | 1.000 | Вингейт |
| Вингейт | 0.350* | 0.154 | 0.414** | 0.166 | 1.000 |
Примечание: обозначения см. табл. 2.
Регион “ключицы” не проявляет связей также с межлопаточной областью, но тесно коррелирует с проекцией сердца и с проекцией щитовидной железы. В свою очередь, проекции сердца и щитовидной железы связаны достоверной корреляцией. Шея со стороны груди и проекция сердца – два региона, коррелирующие со всеми остальными обследованными локусами (по 5 достоверных корреляций). Скорее всего, это свидетельствует о неспецифическом характере термогенеза в этих регионах. Наименьшее число парных корреляций выявлено для регионов “ключицы” и “шея со стороны спины” (по 3 корреляции), при этом между этими двумя регионами достоверной связи нет. Возможно, это отражает специфичность ответа каждого из этих двух рассматриваемых регионов и, вероятно, означает существование важных различий в устройстве термогенераторов в этих двух локусах. Если для ключиц, опираясь на обширную литературу [31, 33, 34], можно предположить в качестве механизма работы термогенератора бурую жировую ткань, то для области шеи со стороны спины приходится предполагать иной источник термогенной активности. Например, это могут быть собственно мышцы шеи [41].
Не менее любопытны результаты корреляционного анализа между различными модальностями стимулов (табл. 3). Глюкоза как стимул НСТ коррелирует с холодовым воздействием и с Вингейтским тестом. Если первая корреляция ожидаема и основана на вероятном участии БЖТ в термогенезе при этих стимулах, то вторая вызывает больше вопросов. Примечательно, что ни с задержкой дыхания, ни с рамптестом значимых корреляций для глюкозы не выявлено.
Зато эти стимулы коррелируют между собой, что, возможно, отражает их неспецифический характер. Холодовое воздействие демонстрирует достоверную связь с задержкой дыхания, но не коррелирует достоверно с рамптестом. Вингейтский тест коррелирует не только с глюкозой, но и с холодовым воздействием. Это наводит на мысль, что в постнагрузочном термогенезе после вингейтского теста участвует БЖТ или ее аналоги – вероятно, в качестве инструмента нормализации внутренней среды организма. Любопытно, что рамптест и вингейтский тест в качестве стимулов термогенеза между собой не коррелируют. Это говорит о том, что с точки зрения генерации несократительного термогенеза рамптест и вингейтский тест являются различными, не идентичными и даже не аналогичными стимулами.
Использованный инструмент (величина IT для разных локусов и стимулов) объективно измеряет локальный тепловой поток на поверхности тела, но не позволяет оценить мощность термогенератора, которая зависит от метаболической активности и объема термогенной ткани, а также от глубины ее расположения. Поэтому IT не дает точной информации об интенсивности теплового ответа на стимул и только косвенно характеризует активность различных подкожных термогенераторов. Если, например, показатель IT невысок, то неизвестно, по какой из причин: или метаболическая активность термогенной ткани невысокая, или масса этой ткани невелика, или стимул слабоват, то ли расположение ткани слишком глубокое. Для характеристики степени активации термогенератора в ответ на различные стимулирующие воздействия необходимо сравнивать его текущую активность (измеряемую при действии данного стимула) с его же максимальной, которая является результатом предельного зафиксированного функционального напряжения данной термогенной структуры.
Иными словами, для оценки функциональной активности термогенератора предлагатся вычислять отношение IT конкретной области во время текущего воздействия к максимальному IT, зарегистрированному когда-либо для данной области (STI – specific thermogenic index = удельный индекс термогенности). При таком подходе, чем выше значение STI, зарегистрированное при конкретном стимулирующем несократительный термогенез воздействии, тем выше термогенный ответ данного термогенератора на данный стимул, независимо от природы термогенной ткани – будь это БЖТ, бежевая жировая ткань (БеЖТ), скелетные мышцы, содержащие разобщающий белок USP3, или что-то иное, и независимо от ее размеров и “глубины залегания”, поскольку все эти параметры остаются неизменными как при максимальном, так и при субмаксимальном ответе термогенератора.
Так поступают в спортивной физиологии, когда принимают максимальное потребление кислорода за максимум аэробной метаболической активности (ММА) и затем интенсивность любой работы выражают в единицах ММА [42].
Из 5 примененных нами видов стимулирующего НСТ воздействия выделяется одно, самое сильное, приводящее к максимальной активации термогенеза во всех 6 областях наблюдения. Таким воздействием оказался рамптест – тяжелая физическая нагрузка нарастающей мощности до достижения МПК и отказа от продолжения работы, сопровождающаяся всеми возможными стимулами несократительного термогенеза: симпатической активацией, субстратной стимуляцией (глюкоза крови), гормональной стимуляцией и метаболической провокацией (лактат). При этом во всех локусах именно при рамптесте (а точнее – в восстановительном периоде после выполнения рамптеста, когда сократительная активность мышц уже отсутствует) были достигнуты максимальные значения индекса термогенности IT. Поэтому в дальнейшем для оценки локальной термоактивности вычисляли STI в долях единицы, где за единицу принимали термоактивность данного локуса во время восстановления после рамптеста.
Такой способ расчета показал, что самый активный термогенератор находится в области ключиц (рис. 3); второе место занимает проекция щитовидной железы; далее следует межлопаточная область; затем – шея со стороны груди и со стороны спины; замыкает ряд грудина вместе с проекцией сердца, которая отражает, по-видимому, активность сократительного термогенеза миокарда (присутствующего всегда).
Рис. 3.
Диаграмма, построенная по величинам STI в 6 исследованных локусах в ответ на 4 стимула различной модальности (среднегрупповые значения). Шкала в ед. STI.

Таким образом, при расчете STI получают “перевернутую” картину паттерна ответа организма на стимулы, однако такая картина, вероятно, наиболее правдоподобна, поскольку она понятна с точки зрения имеющихся данных мировой литературы. Так, именно надключичные ямки, в которых с помощью ПЭТ неоднократно показано типичное расположение БЖТ у взрослого человека [33], демонстрируют в нашем исследовании наибольшую термогенную активность по показателю STI.
Как следует из рис. 3, при всех использованных стимулах соотношение термогенных реакций в различных локусах остается практически одинаковым: на первом месте по величине STI находятся ключицы, на последнем – грудина вместе с проекцией сердца. Сердечная мышца самым активным образом участвует в обеспечении выполнения рамптеста – достаточно длительной нагрузки, при которой ЧСС достигает 180 и более уд./мин и удерживается на таком уровне несколько минут, что, очевидно, достаточно для проведения инфракрасного излучения сквозь ткани организма, несмотря на большую глубину залегания данного термогенератора. А вот при выполнении Вингейтского теста сердце хотя и достигает сходной интенсивности работы, но удерживает ее очень кратковременно: общая длительность активной фазы теста составляет 30 с, а в восстановительном периоде интенсивность работы сердца быстро снижается. В результате, при выполнении Вингейтского теста проекция сердца не является ведущим термогенератором и мало отличается от других локаций термогенеза.
Если проекция сердца отвечает на все стимулы с одинаковой интенсивностью, то этого нельзя сказать про другие локации. Так, для ключиц и проекции щитовидной железы наиболее термогенным стимулом оказалась глюкоза, тогда как для области шеи как со стороны спины, так и со стороны груди, самым сильным стимулом из четырех оказался вингейтский анаэробный тест. Межлопаточная область практически одинаково реагировала на стимулы всех модальностей.
На основании этих наблюдений можно подразделить изучаемые термогенераторы на 3 группы:
1) термогенераторы, наиболее чувствительные к субстратному стимулу (глюкозе) и демонстрирующие высокую активацию – ключицы, проекция щитовидной железы и межлопаточная область (БЖТ?);
2) термогенераторы, равно чувствительные ко всем стимулам и демонстрирующие низкую активацию – проекция сердца на грудине – сердце;
3) термогенераторы, наиболее чувствительные к метаболическим сдвигам в организме и демонстрирующие умеренную активацию – шея со стороны спины и со стороны груди (тонические мышцы?).
Из каких тканей в реальности состоят эти термогенераторы – вопрос, представляющий значительный интерес, но пока не находящий определенного ответа в имеющихся данных. Можно только предполагать, что 1 группа термогенераторов состоит из разновидностей БЖТ; вторая – не имеет отношения к несократительному термогенезу; третья – вероятнее всего, представляет собой тоническую мускулатуру, содержащую разобщающий белок UCP3 [43], что позволяет этой ткани демонстрировать очень высокую метаболическую активность в отсутствии мышечных сокращений.
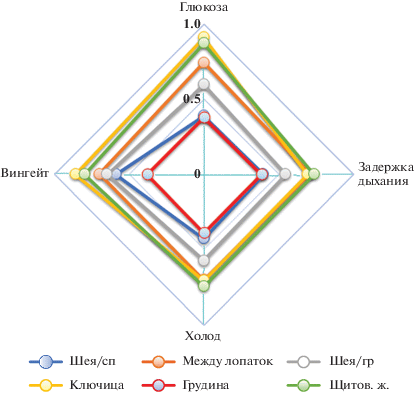
Не менее примечательно сопоставление силы воздействия стимулов различной модальности на каждый из наблюдаемых термогенераторов. Как уже сказано, абсолютный “чемпион” в этом отношении – рамптест, стимулирующий по максимуму все термогенные структуры – во всех локусах зафиксировали максимальный уровень IT в восстановительном периоде после рамптеста. Удельный индекс термогенности STI этого воздействия во всех рассматриваемых локусах равен 1.00. Вслед за ним с существенным отставанием следует вингейтский анаэробный тест, STI которого в среднем по всем областям среди всех испытуемых составил 0.66, и сразу за ним с STI = = 0.65 – глюкозотолерантный тест, классический стимулятор БЖТ. Слабее оказалась активация термогенеза за счет другого классического стимулятора – холодового воздействия (охлаждения ног) (STI = 0.59) или за счет стрессогенного воздействия – задержки дыхания (0.58). Эти результаты наглядно представлены на рис. 4.
Рис. 4.
Степень активации термогенераторов в разных локусах при стимуляции различной модальности (среднегрупповые значения). Шкала в ед. STI.
Проведенный корреляционный анализ (в данном случае – ранговая корреляция по Спирмену) показал значительное отличие взаимосвязей между локусами и стимулами при сопоставлении по STI по сравнению с IT. При IT – это презентация термоактивности генератора на поверхности тела, а при STI – степень активации термогенератора по отношению к максимальной.
Ранговая корреляция (табл. 4) показала, что между разными локусами достоверных связей по степени активации локальных термогенераторов всего три: шея/сп коррелирует с лопатками и сердцем, а шея/гр коррелирует с ключицами. Проекция щитодвидной железы ни с чем не коррелирует. Такая разобщенность локальных термогенераторов может быть следствием их различной анатомо-функциональной природы, которая определяет степень активации при каждом конкретном стимуле.
Таблица 4.
Корреляция (ранговая) между локусами НСТ по STI
| n = 32, r ≥ 0.36 | Шея/сп | |||||
|---|---|---|---|---|---|---|
| Шея/сп | 1.000 | Между лопаток | ||||
| Лопатки | 0.559** | 1.000 | Шея/гр | |||
| Шея/гр | 0.330 | 0.075 | 1.000 | Ключицы | ||
| Ключ | 0.093 | 0.181 | 0.391* | 1.000 | Грудина | |
| Грудина | 0.532** | 0.330 | 0.319 | 0.067 | 1.000 | Щитов. ж. |
| Щитов. ж. | 0.190 | 0.160 | 0.206 | 0.302 | 0.157 | 1.000 |
Примечание: обозначения см. табл. 2.
Что касается взаимосвязей между разными стимулами (табл. 5), то здесь, наоборот, видно полную согласованность всех использованных воздействий. Все выявленные корреляции имеют высокую достоверность и высокие значения, но наиболее сильные связи с другими стимулами демонстрирует холодовое воздействие. Возможно, это отражает наличие компонента β-адренергической стимуляции (важнейшей для холодового стимула) во всех использованных модальностях стимулов. Возможно, также, что это является проявлением общей биологической природы термогенных реакций на использованные воздействия, заключающейся в активности разобщающих белков в различных тканях организма.
Таблица 5.
Корреляция (ранговая) между стимулами НСТ по STI
| n = 48, r ≥ 0.28 | Глюкоза | |||
|---|---|---|---|---|
| Глюкоза | 1.000 | Задержка | ||
| Задержка | 0.594** | 1.000 | Холод | |
| Холод | 0.751** | 0.707** | 1.000 | Вингейт |
| Вингейт | 0.623** | 0.582** | 0.730** | 1.000 |
Примечание: обозначения см. табл. 2.
Анализ представленных данных показывает, что сильнее всего на все воздействия отвечают термогенераторы ключиц и проекции щитовидной железы, а также неожиданно высокую активность проявляет термогенератор в межлопаточной области.
Что касается ключиц – то это вполне ожидаемый эффект. Менее ожидаемым – можно назвать активность щитовидной железы, хотя в литературе имеются данные о тесной взаимосвязи активности БЖТ и щитовидной железы [38] – одного из ключевых регуляторов окислительного метаболизма в организме.
Что касается межлопаточной области, то ее высокая активность не вполне согласуется с данными мировой литературы, согласно которым у взрослых людей межлопаточный бурый жир отсутствует [34], но хорошо корреспондирует с опубликованными ранее данными наших исследований о взаимосвязи температуры спины и аэробной производительности спортсменов [32]. Согласно анатомо-биохимическому исследованию в онтогенезе крысы, выполненному еще 40 лет назад, было показано [44], что к взрослому состоянию межлопаточный бурый жир не обязательно исчезает или полностью заменяется соединительной тканью, но чаще всего между лопатками остаются его небольшие фрагменты, сохраняющие типичную для БЖТ очень высокую метаболическую активность.
Следует обратить внимание на активное участие в термогенезе тканей, расположенных на передней и на задней поверхности шеи. У взрослых грызунов между мышцами шеи обычно располагаются многочисленные фрагменты БЖТ [44]. В то же время на сканах ПЭТ взрослых людей лишь иногда выявляются локусы активной БЖТ в области шеи [36, 37]. Между тем, в наших исследованиях метаболическая активность шеи не была высокой при остром охлаждении и глюкозной нагрузке или при задержке дыхания, что не согласуется с предположением о наличии здесь БЖТ. Зато эта активность сильно увеличивалась в конце и после тяжелой физической нагрузки. В предыдущих публикациях мы предположили, что этот экстраординарный нагрев поверхности шеи в конце тяжелой работы отражает активное поглощение молочной кислоты бурым жиром, расположенным в промежутках между пучками шейных мышц [29]. Однако слабая реакция этих структур на типичные для БЖТ стимуляторы – холод и глюкозу – заставляют усомниться в прежней гипотезе и высказать новое предположение: наблюдаемый термогенный эффект может быть проявлением несократительного термогенеза самих тонических скелетных мышц шеи. Не случайно скелетные мышцы располагают своим собственным разобщающим белком UCP3 [43], который работает так же, как и UCP1 в БЖТ или UCP2 во внутренних органах [45]. Согласно данным молекулярно-генетических исследований, присутствие в организме активного разобщающего белка UCP2 может благоприятно сказываться на спортивных результатах в тех видах, в которых работает преимущественно анаэробная энергетика [46]. Так или иначе, шея представляется локусом максимальной термогенной активности в восстановительном периоде после тяжелой физической нагрузки, и эта постнагрузочная активность не связана с сокращением мышц, т.е. является несократительным термогенезом. Скорее всего, этот термогенез направлен на решение гомеостатических задач на фоне серьезных изменений внутренней среды организма в результате напряженной мышечной деятельности.
Интересно проанализировать индивидуальные особенности реагирования термогенераторов на стимулы различной модальности. Результаты основаны на наблюдениях за 8 испытуемыми, и в табл. 6 приведены диаграммы, построенные по значениям STI каждого из них, которые отражают паттерны индивидуальных реакций на использованные стимулы. Поскольку все представленные в табл. 6 лепестковые диаграммы паттернов индивидуальных реакций на стимулы разной модальности выполнены в одном масштабе, занимаемая ими площадь (S) несет количественную информацию о степени термогенной реактивности организма каждого из испытуемых на предъявленные стимулы. Эта площадь указана в правом верхнем углу диаграмм, представляющих паттерны индивидуальных реакций на стимулы различной модальности.
Таблица 6.
Паттерны индивидуальных реакций на стимулы различной модальности и средняя активность термогенераторов у испытуемого
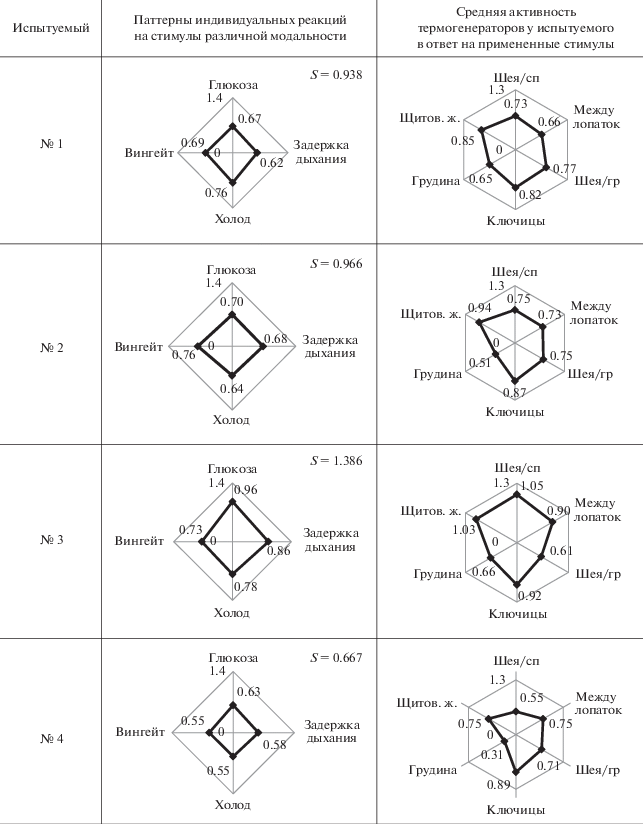

Как видно из приведенных данных, индивидуальная реактивность испытуемых на комплекс использованных стимулов (глюкозный тест, холодовая проба, задержка дыхания и Вингейтский анаэробный тест) может различаться в 3 раза: минимальный зафиксированный уровень площади четырехугольника составляет 0.387, тогда как максимальный результат – 1.387.
Для анализа активности разных термогенераторов выделяли на каждой диаграмме, отражающей среднюю активность термогенераторов у данного испытуемого в ответ на примененные стимулы, по два локуса, показывающих наибольший ответ (в одном случае их оказалось три). При этом не обнаружили каких-либо тенденций в участии различных по локализации термогенераторов в реакциях на оказанные воздействия. Высокая степень реагирования чаще всего демонстрируется локусом “ключицы” – 6 случаев из 8. В тех двух случаях, когда максимум демонстрируют не ключицы, их место занимает “шея спины” – вероятно, это говорит о различии в тканевой природе этих двух термогенераторов. Не менее реактивен локус “щитов. ж.” – также 6 случаев из 8. Только в 2 случаях из 6 эти реакции в данных локусах (“ключицы” и “щитов. ж.”) проявляются асинхронно – либо в одном, либо в другом, а чаще всего они совпадают, что может быть свидетельством в пользу наличия функциональной связи между метаболической активностью щитовидной железы и БЖТ, о чем есть упоминания в литературе [38].
В 3 случаях из 8 высокую реактивность демонстрирует локус “лопатки”; в 2 случаях – локус “шея спина”, а вот “шея грудь” и “проекция сердца” не входят в число наиболее активно реагирующих структур ни разу.
Полученные результаты демонстрируют огромное индивидуальное разнообразие реакций несократительного термогенеза, что, по всей видимости, объясняется его факультативностью и адаптивностью [47]. Из этих результатов вытекает, что наиболее чувствительный термогенератор расположен в области ключиц. Сходную чувствительность демонстрирует локус “щитов. ж.”, что вероятнее всего сопряжено с ее активной метаболической реакцией на примененные стимулы. Существенную реакцию демонстрирует локус “между лопаток”, однако связано ли это с активацией сохранившихся здесь фрагментов БЖТ, или мы видим проявления несократительного термогенеза мышечной ткани, с уверенностью сказать невозможно. Область шеи со стороны спины также демонстрирует свою способность реагировать на использованные стимулы, и здесь так же, как и в случае межлопаточной области, пока нет однозначного понимания природы данного термогенератора: бурый жир или тоническая мускулатура. Ну и, наконец, проекция сердца на поверхность грудной клетки представляет собой термогенератор, наименее чувствительный к тому набору стимулов, которые мы использовали. Самая сильная реакция данного термогенератора была на рамптест, что вполне естественно, но это проявление сократительного термогенеза, который находится вне поля нашего внимания в данном исследовании.
ЗАКЛЮЧЕНИЕ
Что показало проведенное исследование?
Во-первых, наличие в организме нескольких термогенераторов, способных к несократительному термогенезу в ответ на стимулы симпатического и стрессогенного характера. Повышение температуры происходит не тотально и равномерно по всей поверхности тела, а в отдельных локусах с различающейся конфигурацией и интенсивностью – и это обычная реакция организма в ответ на стимулы совершенно различной модальности.
Во-вторых, выявлены глубокие индивидуальные различия в реакции термогенераторов на один и тот же набор стимулов. Эти реакции не отличаются ни синхронностью, ни соразмерностью и зависят, вероятно, от адаптивного опыта испытуемого, т.е. от условий его жизни и других эпигенетических факторов.
В-третьих, доказана наивысшая чувствительность термогенераторов, расположенных в надключичной области, к испытанному набору стимулирующих воздействий. Доказана также тесная функциональная связь этого термогенератора с проекцией щитовидной железы. Все это подтверждает сложившееся уже убеждение в том, что этот термогенератор представляет собой именно БЖТ.
В-четвертых, между исследованными термогенераторами обнаружены не вполне тривиальные взаимоотношения, которые нуждаются в дальнейшем изучении. В частности, с точки зрения температурного ответа не выявлено параллелизма между максимальной аэробной и максимальной анаэробной физической нагрузкой. Реактивность всех исследованных термогенераторов в ответ на все примененные стимулы оказалась синхронизирована. Причины выявленных особенностей в настоящий момент не ясны и требуют дальнейшего изучения.
В-пятых, полученные данные заставляют задуматься не только о роли БЖТ в регуляции энергетического метаболизма, которая сегодня привлекает огромное число исследователей, занятых проблемой борьбы с метаболическим синдромом, но и других тканей, способных участвовать в этой регуляционной программе организма. В частности, следует обратить внимание на мускулатуру, располагающую разобщающим белком UCP3, выполняющим такую же функцию, как UCP1 в буром жире.
На основании полученных данных нельзя однозначно говорить о единстве или различиях природы исследованных термогенераторов. Однако широкое применение неинвазивного и безопасного тепловизора в различных экспериментальных ситуациях, позволит накопить научные факты, необходимые для дифференциальной диагностики различных видов термогенераторов в организме человека. Возможно, это будет способствовать решению как клинических, так и других актуальных прикладных задач современной физиологии.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены локальным биоэтическим комитетом Центра спортивных технологий и сборных команд Москомспорта (Москва).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа выполнена в рамках тематики НИР кафедры физиологии Российского государственного университета физической культуры, спорта, молодежи и туризма на 2015–2020 гг. (Сводный план НИР РГУФКСМиТ, тема 03.00.12. “Гомеостатический несократительный термогенез у человека. Взаимодействие механизмов факультативного несократительного термогенеза и адаптации к физической нагрузке”), а также при материально-технической поддержке ЦСТ и СК Москомспорта.
Благодарности. Авторы благодарят добровольцев – участников экспериментального исследования за плодотворное сотрудничество; к. б. н. Р.С. Андреева и А.В. Якушкина за продуктивное обсуждение методических подходов и полученных результатов, а также О.И. Парфентьеву за помощь в статистическом анализе результатов.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Колесов С.Н., Воловик М.Г. Современная методология тепловизионных исследований и тепловизионная медицинская аппаратура // Оптический журн. 2013. Т. 80. № 6. С. 59.
Колесов С.Н., Воловик М.Г., Прилучный М.А. Медицинское теплорадиовидение: современный методологический подход. Н. Новгород: ННИИТО, 2008. 184 с.
Fernández-Cuevas I., Marins J.C.B., Lastras H.A. et al. Classification of factors influencing the use of infrared thermography in humans // Infrared Physics & Technology. 2015. V. 71. P. 28.
Jang C., Jalapu S., Thuzar M. et al. Infrared thermography in the detection of brown adipose tissue in humans // Physiol Rep. 2014. V. 2. e12167.
Cramer M.N., Jay O. Explained variance in the thermoregulatory responses to exercise: the independent roles of biophysical and fitness/fatness-related factors // J. Appl. Physiol. 2015. V. 119. P. 982.
Акимов Е.Б., Андреев Р.С., Каленов Ю.Н. и др. Температурный портрет человека и его связь с аэробной̆ производительностью и уровнем лактата в крови // Физиология человека. 2010. Т. 36. № 4. С. 89.
Акимов Е.Б., Андреев Р.С., Каленов Ю.Н. и др. Возможности инфракрасной термографии по выявлению морфофункциональных характеристик человека (детей и взрослых) // Вестник Московского университета, Серия 23 “Антропология”. 2016. № 3. С. 49.
Chondronikola M., Beeman S.C., Wahl R.L. Non-invasive methods for the assessment of brown adipose tissue in humans // J. Physiol. 2018. V. 596. № 3. P. 363.
Orava J., Nuutila P., Lidell M.E. et al. Different metabolic responses of human brown adipose tissue to activation by cold and insulin // Cell Metab. 2011. V. 14. P. 272.
Stanford K.I., Middelbeek R.J., Townsend K.L. et al. Brown adipose tissue regulates glucose homeostasis and insulin sensitivity // J. Clin. Invest. 2013. V. 123. P. 215.
Robinson L.J., Law J.M., Symonds M.E. et al. Brown adipose tissue activation as measured by infrared thermography by mild anticipatory psychological stress in lean healthy females // Exp. Physiol. 2016. V. 101. P. 549.
Formenti D., Ludwig N., Rossi A. et al. Skin temperature evaluation by infrared thermography: Comparison of two image analysis methods during the nonsteady state induced by physical exercise // Infrared Physics & Technology. 2017. V. 81. P. 32.
Иваницкий Г.Р., Деев А.А., Пашовкин Т.Н. и др. Особенности теплового проявления подкожных источников нагрева на поверхности тела человека // Докл. АН. 2008. Т. 420. № 4. С. 551.
Хижняк Л.Н., Хижняк Е.П., Иваницкий Г.Р. Диагностические возможности матричной инфракрасной термографии. Проблемы и перспективы // Вестник новых медицинских технологий. 2012. Т. 19. № 4. С. 170.
Gatidis S., Schmidt H., Pfannenberg C.A. et al. Is it possible to detect activated brown adipose tissue in humans using single-time- point infrared thermography under thermoneutral conditions? Impact of BMI and subcutaneous adipose tissue thickness // PLoS One. 2016. V. 11. P. e0151152.
Nedergaard J., Bengtsson T., Cannon B. Unexpected evidence for active brown adipose tissue in adult humans // Am. J. Physiol. Endocrinol. Metab. 2007. V. 293. P. E444.
Кокшарова Е.О., Майоров А.Ю., Шестакова М.В. и др. Метаболические особенности и терапевтический потенциал бурой и “бежевой” жировой ткани // Сахарный диабет. 2014. № 4. С. 5.
Harms M., Seale P. Brown and beige fat: development, function and therapeutic potential // Nature Medicine. 2013. V. 19. № 10. P. 1252.
Spiegelman B.M. Banting Lecture 2012: Regulation of adipogenesis: toward new therapeutics for metabolic disease // Diabetes. 2013. V. 62. № 6. P. 1774.
Sebo Z.L., Rodeheffer M.S. Assembling the adipose organ: adipocyte lineage segregation and adipogenesis in vivo // Development. 2019. V. 146. № 7. https://doi.org/10.1242/dev.172098
Rui L. Brown and Beige Adipose Tissues in Health and Disease // Compr. Physiol. 2017. V. 7. № 4. P. 1281.
Kim K., Huang S., Fletcher L.A. et al. Whole Body and Regional Quantification of Active Human Brown Adipose Tissue Using 18F-FDG PET/CT // J. Vis. Exp. 2019. № 146. https://doi.org/10.3791/58469
Schweizer S., Oeckl J., Klingenspor M. et al. Substrate fluxes in brown adipocytes upon adrenergic stimulation and uncoupling protein 1 ablation // Life Sci. Alliance. 2018. V. 1. № 6. P. e201800136.
Reitman M.L. Of mice and men – environmental temperature, body temperature, and treatment of obesity // FEBS Lett. 2018. V. 592. № 12. P. 2098.
Wang S., Subramaniam A., Cawthorne M.A. et al. Increased fatty acid oxidation in transgenic mice over-expressing UCP3 in skeletal muscle // Diabetes, Obesity and Metabolism. 2003. V. 5. № 5. P. 295.
Волков Н.И., Мелихова М.А., Олейников В.И., Тамбовцева Р.В. Общая биохимия и биохимия физических упражнений: Учебное пособие: В 2 ч. М.: Ф-ГБОУ ВПО “РГУФКСМИТ”, 2015. Ч. 1. 344 с.
Hankir M.K., Klingenspor M. Brown adipocyte glucose metabolism: a heated subject // EMBO Rep. 2018. V. 19. № 9. P. e46404.
De Matteis R., Lucertini F., Guescini M. et al. Exercise as a new physiological stimulus for brown adipose tissue activity // Nutr Metab Cardiovasc 2012. № 6. P. 582.
Son’kin V.D., Akimov E.B., Andreev R.S. et al. Brown Adipose Tissue Participate in Lactate Utilization during Muscular Work // icSPORTS. Proceedings of the 2-nd International Congress on Sports Sciences Research and Technology Support, 2014. P. 97.
Козлов А.В., Сонькин В.Д., Якушкин А.В. Метод исследования активности подкожных термогенных структур при действии стимулов разной модальности // Физиология человека. 2017. Т. 43. № 6. С. 124.
Symonds M.E., Henderson K., Elvidge L. et al. Thermal imaging to assess age-related changes of skin temperature within the supraclavicular region co-locating with brown adipose tissue in healthy children // J. Pediatr. 2012. V. 161. № 5. P. 892.
Акимов Е.Б., Андреев Р.С., Каленов Ю.Н. и др. Температурный портрет человека и его связь с аэробной производительностью и уровнем лактата в крови // Физиология человека. 2010. Т. 36. № 4. С. 89.
Law J., Chalmers J., Morris D.E. et al. The use of infrared thermography in the measurement and characterization of brown adipose tissue activation // Temperature (Austin). 2018. V. 5. № 2. P. 147.
Ong F.J., Ahmed B.A., Oreskovich S.M. et al. Recent advances in the detection of brown adipose tissue in adult humans // Clin. Sci. (Lond.). 2018. V. 132. № 10. P.1039.
Nedergaard J., Cannon B. Brown adipose tissue as a heat-producing thermoeffector. Chapter 9. Handbook of Clinical Neurology. V. 156 (3rd series) / Thermoregulation: From Basic Neuroscience to Clinical Neurology, Part I. Ed. A.A. Romanovsky. 2018. https://doi.org/10.1016/B978-0-444-63912-7.00009-6
Jaksic P.V., Grizelj D., Livun A. et al. Neck adipose tissue – tying ties in metabolic disorders // Horm. Mol. Biol. Clin. Investig. 2018. V. 33. № 2. https://doi.org/10.1515/hmbci-2017-0075
Lee P., Werner C.D., Kebebew E. et al. Functional thermogenic beige adipogenesis is inducible in human neck fat // Int. J. Obesity. 2014. V. 38. № 2. P. 170.
Weiner J., Hankir M., Heiker J.T. et al. Thyroid hormones and browning of adipose tissue // Mol. Cell. Endocrinol. 2017. V. 458. P. 156.
Cannon B., Nedergaard J. Brown Adipose Tissue: Function and Physiological Significance // Physiol. Rev. 2004. V. 84. P. 277.
Lidell M.E., Betz M.J., Enerbäck S. Brown adipose tissue and its therapeutic potential // J. Intern. Med. 2014. V. 276. № 4. P. 364.
Astrup A. Thermogenesis in human brown adipose tissue and skeletal muscle induced by sympathomimetic stimulation // Acta Endocrinol. Suppl. (Copenh.). 1986. V. 278. P. 1.
Волков Н.И., Попов О.И., Габрысь Т., Шматлян-Габрысь У. Физиологические критерии нормирования тренировочных и соревновательных нагрузок в спорте высших достижений // Физиология человека. 2005. Т. 31. № 5. С. 125.
Oliveira B.A., Pinhel M.A., Nicoletti C.F. et al. UCP1 and UCP3 Expression Is Associated with Lipid and Carbohydrate Oxidation and Body Composition // PLoS One. 2016. V. 11. № 3. e0150811.
Грибова Н.В., Сонькин В.Д. Развитие бурой жировой ткани в онтогенезе / Морфофункциональные особенности растущего организма. М.: АПН СССР, 1974. С. 120.
Hagen T., Vidal-Puig A. Mitochondrial uncoupling proteins in human physiology and disease // Minerva Med. 2002. V. 93. № 1. P. 41.
Бондарева Э.А., Парфентьева О.И., Козлов А.В. и др. Полиморфизм гена UCP2 реципрокно ассоциирован с аэробной и анаэробной производительностью у спортсменов // Физиология человека. 2018. Т. 44. № 6. С. 79.
van Marken Lichtenbelt W.D., Schrauwen P. Implications of nonshivering thermogenesis for energy balance regulation in humans // Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011. V. 301. P. R285.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека