

Физиология человека, 2020, T. 46, № 3, стр. 90-98
Иллюзия наклона и ориентационная чувствительность
В. М. Бондарко *
ФГБУН Институт физиологии имени И.П. Павлова РАН
Санкт-Петербург, Россия
* E-mail: vmbond@gmail.com
Поступила в редакцию 25.02.2019
После доработки 03.04.2019
Принята к публикации 05.10.2019
Аннотация
Оценивали ориентацию линий в разных условиях наблюдения. В первом эксперименте получено искажение в оценке ориентации линий на 1°–9° в случае присутствия соприкасающихся линий с другой ориентацией (иллюзия наклона). При уменьшении длины линий до 6 угл. мин иллюзия увеличивалась, а пороги различения ориентации повышались. Для линий с длиной в 6 угл. мин иллюзия аппроксимирована моделью, где ориентация задавалась наклоном наименьших по размеру рецептивных полей (РП) максимально реагирующих на изображения нейронов. Адекватность модели подтвердили данные второго эксперимента, в котором определяли минимальную длину линий, необходимую для различения вертикальных линий от горизонтальных. Минимальные длины оказались согласованными с шириной зон модельных РП. Взаимодействия между РП, влияющие на иллюзию, оценивали в третьем эксперименте при различении ориентации линий с длиной в 60 угл. мин. Полученные ориентационные зависимости от возраста наблюдателей позволяет предположить, что взаимодействия между РП способствуют уменьшению иллюзии наклона.
Еще в XIX в. был описан класс зрительных иллюзий, которые могут возникать вследствие искажения оценки ориентации линий из-за присутствия линий с другой ориентацией. К ним относят известные иллюзии Поггендорфа, Цольнера, Фрэйзера. Неправильная оценка ориентации из-за влияния добавочных изображений носит название иллюзии наклона. В первоначальных работах J.J. Gibson и M. Radner [1, 2] показано, что после адаптации к наклонной линии вертикальная или горизонтальная линии кажутся повернутыми в сторону, противоположную к наклонной, т.е. наблюдается эффект отталкивания. При разнице в ориентации между тестируемым и индуцирующим стимулом более чем на 90° был обнаружен противоположный эффект – эффект притягивания. Линия казалась наклоненной к дополнительной линии. В более поздних работах были получены иллюзии, когда стимулами являлись решетки или линии с ориентацией, отличной от вертикальной и горизонтальной.
Тема взаимодействия между ориентациями изображений получила интенсивное развитие после опубликования работ по оценке ориентационной чувствительности отдельных РП нейронов стриарной коры и психофизических данных, свидетельствующих в пользу существования в зрительной системе ориентационно избирательных пространственно-частотных каналов. Были уточнены ранее полученные зависимости. Показано, что максимальное искажение в оценке ориентации варьирует от 1° до 10° [3–6]. При разнице в ориентации тестируемого и дополнительного изображений в 90° искажение отсутствует. Наблюдалась разница в использовании в качестве тестируемых изображений линий и решеток. При использовании решеток и разнице в ориентации окружения на 65°–80° получен эффект притягивания [7]. Некоторые исследователи выявили эффект притягивания и при малой разнице между ориентациями тестируемых и добавочных изображений [8, 9].
При объяснении иллюзии наклона наибольшее распространение получила гипотеза C. Blakemore, R.H.S. Carpenter и M.A. Georgeson [10] о тормозном латеральном взаимодействии между ориентационными каналами. Предполагается, что основной тестируемый стимул активизирует один ориентационный канал, а дополнительный стимул – другой. Суммарное взаимодействие этих каналов приводит к искажению оценки ориентации основного стимула. Были сделаны попытки проверить гипотезу о взаимодействии между ориентационными каналами [9, 11, 12] и промоделировать ориентационные зависимости [13]. В модели [13] сначала происходит фильтрация изображений отдельными РП, а затем в ассоциативном поле происходит объединение их реакций.
Несмотря на большое количество исследований, иллюзию продолжают изучать и предлагают новые интерпретации этого феномена. В данной работе представлены результаты, позволяющие высказать предположение о роли взаимодействий между РП, влияющих на иллюзию наклона. Сначала в ней исследовали влияние на иллюзию размера стимула. Оказалось, что максимальная по величине иллюзия наблюдается для самых коротких из числа исследованных линий с длиной в 6 угл. мин. Для этого случая была разработана модель, которая объясняла полученные данные. В основу модели взяты результаты исследований H.R. Wilson и D.J. Gelb [14], описавших весовые функции РП. Затем сопоставили параметры модельных РП с результатами исследований по оценке ориентационной избирательности, а также по нахождению минимальных длин линий, при которых происходит грубое различение ориентации. Эти исследования были проведены на наблюдателях различных возрастных групп с целью изучения формирования взаимодействий между ориентационно избирательными РП в онтогенезе. Полученные результаты позволили высказать предположение о том, что взаимодействия между РП не способствуют возникновению иллюзии наклона, а скорее наоборот уменьшают ее последствия.
МЕТОДИКА
Стимулами служили отрезки прямых линий. В первом эксперименте использовали либо одну, либо две линии с примыкающими друг к другу концами (рис. 1, А). Дополнительная линия была в 1.2 раза длиннее. Ее ориентация отличалась от ориентации референтной тестируемой линии на ±10°, ±20° или ±30°. Наклон короткой линии в стимуле сравнивали с наклоном одиночной линии такой же длины. Шаг изменения ориентации этой линии (тестовой) подбирали индивидуально для каждого наблюдателя, варьировался в пределах 1°–3°. Длина референтных линий была равна 6, 12 или 24 угл. мин, ориентация 30°.
Рис. 1.
Примеры используемых в исследовании стимулов. А, В – референтные и тестовые стимулы первого и третьего экспериментов. Тестовые линии показаны пунктиром. Б – стимулы второго эксперимента.
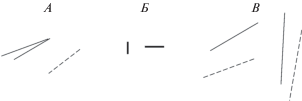
Во втором эксперименте определяли минимальную длину линий, при которой возможно отличить вертикальную линию от горизонтальной. Вертикальные или горизонтальные линии предъявляли в случайном порядке (рис. 1, Б). Ширина линий составляла 0.94 угл. мин, длина менялась в пределах от 1.4 до 4.2 угл. мин с шагом 0.47 угл. мин случайным образом. Наблюдатель должен был ответить, какую линию он видит: горизонтальную или вертикальную.
В третьем эксперименте оценивали ориентационную чувствительность с использованием линий с длиной 60 угл. мин. Референтные линии были повернуты на 30° или 87° относительно горизонтали. Тестовые линии имели ту же длину. Шаг изменения ориентации тестовой линии 1°–2°.
Время предъявления линий в экспериментах 1 с. В первом и третьем экспериментах наблюдатель смотрел в центр экрана, на котором сначала появлялся референтный, а затем через 1 с тестовый стимул. Он должен был указать, куда повернута тестовая линия по отношению к референтной – по часовой или против часовой стрелки. Использовали метод вынужденного выбора. Ритм предъявления изображений на экране задавал сам наблюдатель, но после последующего предъявления проходило не менее 1 с.
Изображения синтезировали на стандартном дисплее CTX PR711T. Характеристики дисплея: 17 дюймов по диагонали, шаг апертуры решетки 0.27 мм. Дисплей использовали при разрешении экрана 1024 × 768 пикселов и частоте смены кадров 100 Гц. Яркость экрана равна 80 кд/м2, стимулов –5 кд/м2. Стимулы предъявляли бинокулярно с расстояния до экрана монитора 7.3 м в первом эксперименте и 2.3 м во втором и третьем экспериментах. В первом эксперименте проводили три серии, отличающиеся длиной линий. В работе приведены суммарные данные по десяти опытам в каждой серии, которым соответствует 100 предъявлений стимулов с каждой ориентацией дополнительной и тестовой линий. В нем принимали участие трое наблюдателей с нормальной остротой зрения. Во втором и третьем экспериментах участвовали наблюдатели в возрасте 7–25 лет с остротой зрения, большей 1 ед.: во втором 21 чел. в возрасте 7–8 лет, 26 в возрасте 9 лет, 19 десятилетних и 15 взрослых; в третьем – 16 чел. в возрасте 9 лет, 21 в возрасте 10 лет, 18 в возрасте 11–13 лет, 24 в возрасте 14–16 лет и 17 взрослых. В третьем эксперименте проводили две серии с разной ориентацией референтных линий. Каждому наблюдателю во втором эксперименте референтный стимул с любым тестовым стимулом предъявлялся по 20 раз, в третьем – 40 раз. Кроме того, второй эксперимент с предъявлением стимулов более 100 раз и с меньшим шагом изменения длины проводили на тех же наблюдателях, что и первый эксперимент с расстояния 7.3 м. Результаты второго и третьего экспериментов частично представлены в [15].
Для определения отклонений в восприятии и порогов во всех экспериментах строили психометрические функции. С помощью метода наименьших квадратов психометрические функции аппроксимировали функциями нормального распределения. Величины средних значений у нормальных распределений взяты для оценки иллюзий: они соответствуют тем значениям параметров, при котором наблюдатели считают тестовый и референтный стимул совпадающими. Величина иллюзии – разность между средним и параметром стимула. Пороги вычисляли как стандартные отклонения, что соответствует 84% уровню ответов на психометрической функции.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
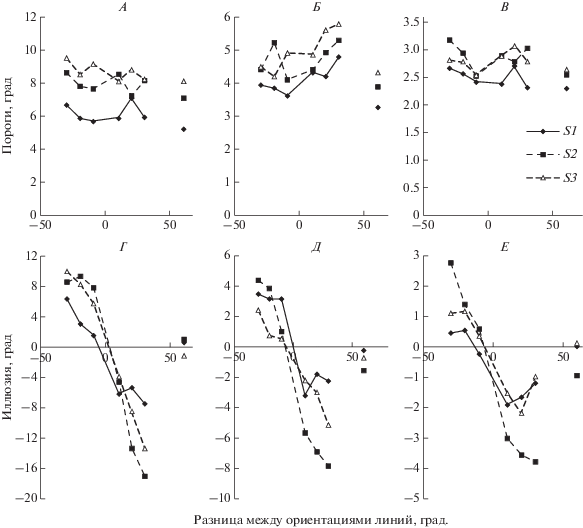
На рис. 2, А–В показаны пороги различения ориентации для линий с длиной 6, 12 и 24 угл. мин в зависимости от разницы в ориентации между короткой и дополнительной линиями. Видна тенденция увеличения порогов в присутствии дополнительных линий по сравнению с порогами различения ориентации одиночных линий. Пороги возрастают по мере увеличения разности в ориентациях линий и уменьшении их длины. На рис. 2, Г–Е приведены кривые зависимостей иллюзий от разницы в ориентациях линий. Кривые идут в противофазе к изменению ориентации дополнительных линий. При повороте дополнительной линии против часовой стрелки линия кажется повернутой по часовой стрелке, т.е. наблюдается эффект отталкивания. Иллюзия проявляется сильнее у наблюдателей, имеющих более высокие пороги различения ориентации. У наблюдателя S2 значения для одиночных линий смещены в область отрицательных значений, за счет этого все кривые опущены вниз. Иллюзии возрастают при уменьшении длины линий.
Рис. 2.
Различение ориентации в иллюзии наклона. А, Б, В – пороги различения ориентации линий с длиной 6, 12 и 24 угл. мин соответственно. Данные наблюдателей S1, S2 и S3. Ось абсцисс – разница между ориентациями референтной и дополнительной линий, град. Ось ординат – пороги, град. Крайние точки справа – пороги различения ориентации одиночных линий без добавочных. Г, Д, Е – иллюзии наклона для линий в 6, 12 и 24 угл. мин. Ось ординат – иллюзии, град. Крайние точки справа – искажение в оценке ориентаций одиночных линий.
Получив искажение в восприятии ориентации даже в случае отсутствия четкого видения двух линий (при длине линий 6 и 7 угл. мин и угле 10° наблюдатель видел асимметричное пятно), возникло предположение, что иллюзия не обязательно связана с взаимодействием между ориентационными каналами (в этом случае трудно вычленить два активизированных канала), а может определяться формой стимула.
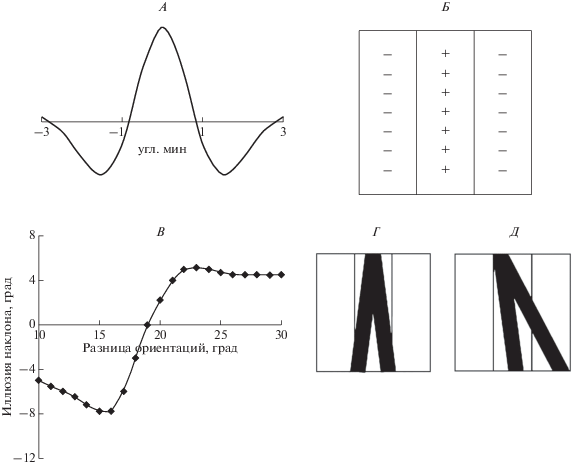
Для проверки этой гипотезы рассчитали интегралы от произведений двумерных весовых функций (ВФ) РП и яркостных функций стимулов. В качестве ВФ была взята самая высокочастотная, приведенная в исследовании H.R. Wilson и D.J. Gelb [14]. Сечение двумерной ВФ в перпендикулярной к оптимальной ориентации РП показано на рис. 3, А. В оптимальной ориентации функцию считали постоянной, что соответствует электрофизиологическим данным [16, 17]. На рис. 3, Б изображена схема РП с такой ВФ, предложенная D.H. Hubel и T.N. Wiesel [16]. Это РП простого нейрона – детектора полосы, которое имеет по бокам две тормозные зоны, а в центре возбудительную. Ширина зон около 2 угл. мин, длина 6 угл. мин. Оптимальная ориентация у РП вертикальная. Вычисляли интегралы при изменении стимулом ориентации и положения в РП. Ширина линий при расчетах взята равной 1 угл. мин. Находили максимальные значения интегралов. На рис. 3, В приведены расчетные значения: показано, при каких ориентациях основной линии достигаются максимальные значения. При малых углах между линиями оптимальная ориентация отклонена от вертикали по часовой стрелке на 5°–9°. При угле больше 20° оптимальной становится ориентация, повернутая по отношению к вертикали против часовой стрелки. На рис. 3, Г, Д показано, при каком положении стимула в РП достигается максимум. Для стимула с малой разницей в ориентации линий (рис. 3, Г) оптимальным в РП является положение, когда стимул симметричен относительно оптимальной ориентации РП. Таким образом при малой разнице в ориентации по расчетам должен наблюдаться эффект притягивания – максимальный ответ дает РП с ориентацией, средней между ориентациями основной и дополнительной линий. При разнице в ориентациях большей 20° наблюдается эффект отталкивания: стимул повернут таким образом, что основная линия (левая) наклонена относительно оптимальной ориентации РП против часовой стрелки (рис. 3, Д). Дополнительная линия (правая) меньше захватывает тормозную зону, тем самым слабее уменьшая ответ возбудительной зоны. Полученный результат моделирования соответствует экспериментальным данным исследователей, которые показали наличие эффекта притягивания при разнице в ориентации в 10° [8, 9]. Можно заметить, что с уменьшением размера РП в модели две линии будут попадать в возбудительную зону при меньшем между ними угле, т.е. при меньшем угле произойдет переход от эффекта притягивания к эффекту отталкивания.
Рис. 3.
Модельное рецептивное поле и его реакция на стимулы. А – сечение двумерной весовой функция в перпендикулярной к оптимальной ориентации РП. Б – карта РП с оптимальной вертикальной ориентацией. В – зависимость ориентации основной линии от разницы в ориентации с дополнительной линией. Ось абсцисс – разница между ориентациями, град. Ось ординат – отклонение основной линии от вертикали, град. Г, Д – положение стимула в РП при максимальном ответе РП с разницей в ориентации линий меньшей 16° или большей 20° соответственно.
Проверку соответствия ВФ модельного РП экспериментальным данным провели следующим образом: определили минимальную длину линий, необходимую для различения вертикальной ориентации от горизонтальной. Нейрон с РП, приведенным на рис. 3, Б, по-разному отвечает на такие линии при выходе линий за пределы возбудительной зоны. Поэтому таким способом можно найти примерную ширину зон минимальных РП. На рис. 4, А показана зависимость процента правильного ответа от длины линии, полученная при различении вертикальной линии от горизонтальной наблюдателями S1, S2 и S3. Процент правильных ответов постепенно увеличивается от уровня случайного гадания (50%) до 81–97% уровня. Выход на плато осуществляется при длине линии 1.8–2.2 угл. мин, что согласуются с размерами возбудительной зоны модельного РП.
Рис. 4.
Различение вертикальной и горизонтальной ориентации в зависимости от длины линий. А, Б – процент правильного ответа. Ось абсцисс – длина линий, угл. мин. А – данные наблюдателей S1, S2 и S3, Б – данные взрослых из группового обследования. В – пороговые длины линий. Ось абсцисс – возраст наблюдателей. Ось ординат – пороги, угл. мин. Вертикальные линии – стандартные ошибки.
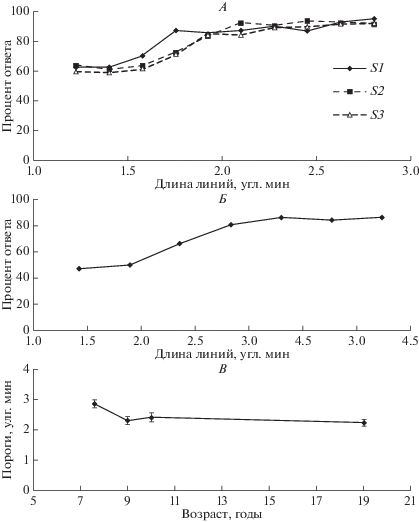
Аналогичные данные получены на группе наблюдателей различного возраста. На рис. 4, Б приведен усредненный процент правильных ответов у взрослых. Кривая выходит на плато позднее, чем на рис. 4, А. Скорее это связано с обучением, чем с более низкой остротой зрения. Наблюдатели S1, S2 и S3 были опытными, и к тому же им предъявляли стимулы большее число раз. На рис. 4, В показаны пороговые значения длин линий, при которых происходит различение вертикали от горизонтали в зависимости от возраста наблюдателей. Пороги для взрослых наблюдателей равны 2.4 угл. мин. Попарное сравнение средних величин по критерию Стьюдента выявило значимые различия порогов (на уровне 5%) у наблюдателей 7–8 лет по сравнению с остальными возрастными группами.
Какова роль взаимодействий между РП, если для коротких линий иллюзия возникает вследствие реакции отдельных РП. Для ответа – в следующем эксперименте для оценки ориентации использовали длинные линии, поскольку на них реагируют нейроны с РП различных размеров (на короткие линии с разной ориентацией нейроны с большими РП отвечают практически одинаково).
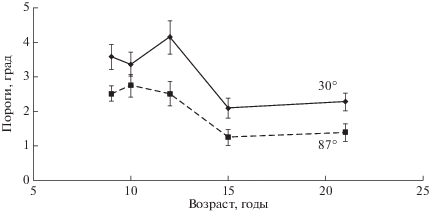
На рис. 5 приведены пороги различения ориентации в зависимости от возраста наблюдателей. Показаны кривые, соответствующие ориентациям 30° и 87°. Пороги выше для ориентации в 30° по сравнению с ориентацией 87°. Попарное сравнение средних величин выявило достоверное увеличение порогов у школьников 8–13-летнего возраста по сравнению с 14–16-летними и взрослыми по критерию Стьюдента (р < 0.05). Ориентационные пороги для взрослых наблюдателей согласованы с данными, полученными в длительных экспериментах [18]. Они составляют примерно 0.8° и 0.5° для ориентации стимулов в 30° и 90° [19]. В нашем исследовании пороги также выше для ориентации в 30°, чем для 87°, и равны 2.2° и 1.3° у взрослых наблюдателей. Расхождения с данными работы [19] могут быть связаны с обучением задаче различения ориентации, при котором пороги уменьшаются в несколько раз [20].
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Выявлены искажения в оценке ориентации линий в присутствии дополнительных линий, зависящие от длины изображений. Величина иллюзии колеблется в пределах 1°–9°, что соответствует литературным данным [3–6]. Линии казались повернутыми в противоположную по отношению к дополнительным линиям сторону, т.е. наблюдался эффект отталкивания. Иллюзия была больше для коротких линий, как и пороги различения ориентации. Ранее было показано, что пороги различения ориентации одиночных линий увеличиваются при уменьшении длин линий [21]. Иллюзию наклона с пропорциональным изменением длин основных и дополнительных линий вплоть до таких малых размеров и углов, что становилось невозможно различать две линии, ранее не исследовали. P. Wenderoth et al. [11] меняли относительные длины примыкающей и основной линий. Для разницы в ориентации линий 15° они показали, что иллюзия уменьшается при фиксированной длине дополнительной линии 2 угл. град. с увеличением длины основной линии от 0.5 до 2.5 угл. град. Эти данные не противоречат приведенным здесь результатам. В данной работе показано, что пороги различения ориентации увеличиваются при уменьшении длин как одиночных линий, так и линий с добавочными контурами. Пороги оказались выше в присутствии примыкающих линий. Это повышение достоверно по совокупности всех данных и критерию знаков (р > 0.95). Повышение порогов в случае присутствия добавочных изображений с другой ориентацией отмечено J.A. Solomon и M.J. Morgan [5], которые в качестве стимулов использовали элементы Габора.
Попытки объяснить иллюзию наклона привели к возникновению гипотезы о латеральном торможении между ориентационными каналами. На схеме, предложенной C. Blakemore et al. [10], представлены профили ориентационной чувствительности двух ориентационных каналов. Они имеют центральные зоны возбуждения, окруженные тормозными зонами. При малой разнице в ориентациях возбудительная зона одного канала совмещается с возбудительной зоной другого ориентационного канала, а при большей разнице – с тормозной зоной другого ориентационного канала. В результате для малой разницы в ориентациях к ориентации тестируемой линии частично добавляется ориентация добавочной линии, возникает эффект отталкивания.
В более поздних исследованиях аппроксимировали экспериментальные зависимости в моделях [9, 12]. Двухслойная модель дала хорошее приближение к экспериментальным данным в работе M.J. Morgan et al. [9]. Сначала в ней изображения фильтруются ориентационно избирательными РП. Затем происходит выпрямление выходного сигнала: отсечение отрицательных значений и замена их нулями, так как зрительная система не передает напрямую торможение. На заключительном этапе происходит нахождение центра тяжести у полученных распределений возбужденных нейронов на следующем уровне зрительной системы. Авторы не вводят дополнительного торможения между РП разной ориентации.
В данной работе иллюзия наклона получена и в условиях отсутствия четкой видимости двух линий при их длине в 6 и 7 угл. мин и разнице в ориентации 10°. Для коротких линий иллюзия промоделирована с использованием полученных H.R. Wilson и D.J. Gelb [14] ВФ РП наименьшего размера. В результате при разнице в ориентации между линиями от 10° до 19° в модели выявлен эффект притягивания, а при дальнейшем увеличении разницы в ориентациях – эффект отталкивания. Этот результат показывает, что нет необходимости вводить взаимодействия между РП для объяснения иллюзии при разнице в ориентациях большей 19°. На две прилегающие линии лучше реагирует РП, повернутое в противоположную к дополнительной линии сторону, поскольку дополнительная линия, попадая в тормозную зону, в этом случае незначительнее уменьшает ответ возбудительной зоны (рис. 3, Д). Именно ориентация РП нейронов, дающих максимальный ответ, видимо, учитывается зрительной системой на следующем уровне при принятии решения об ориентации основной линии. Правда, в описанных экспериментах не выявлен эффект притягивания при малой разнице в ориентациях, полученный в модели, хотя некоторые исследователи отмечают его присутствие [8, 9]. С чем может быть связано расхождение в экспериментальных и модельных результатах. Разница в экспериментальном материале может определяться индивидуальными различиями наблюдателей. P. Wenderoth et al. [11] предъявляли линии разной длины десяти наблюдателям. У двух из десяти они получили эффект притягивания при разнице в ориентациях 10° и длине линий 3 угл. град. Индивидуальные различия могут быть обусловлены наличием отличающихся по размеру наименьших РП у конкретных наблюдателей: при уменьшении размера РП в модели (рис. 3, Г) эффект притягивания будет проявляться для меньшей разницы в ориентации.
Что может происходить с линиями большей длины? В исследовании D.J. Tolhurst и P.G. Thompson [6] показано, что располагающиеся в удаленном на расстояние 0.27 диаметра круга кольце окружающие решетки не искажают ориентацию тестовых решеток, находящихся в круге. Искажения исчезали на расстояниях, превышающих длину линий в 1.4 раза, и в иллюзии жезла-рамки [22]. Можно заключить, что на иллюзию наклона оказывает влияние только близко расположенное окружение, при небольших размерах стимулов попадающее в те же РП, что и тестовое изображение. То есть ключевыми для возникновения иллюзии являются точки пересечения линий с близлежащими отрезками – признаки, используемые и для опознания изображений [23]. Маловероятно, что ориентацию тонких линий оценивают РП больших размеров, поскольку они имеют широкую ориентационную настройку на такие стимулы. Скорее всего, при определении ориентации тонких длинных линий проявляется взаимодействие между небольшими РП, покрывающими линию. При этом дополнительная линия попадает в одни и те же РП с тестируемой только в несколько соседних РП, искажая оценку ее ориентации. Удаленные от точки соприкосновения линий РП, чья реакция нейронов учитывается, обрабатывают только тестируемую линию. Их правильная оценка ориентации уменьшает силу иллюзии.
О взаимодействии ориентационно настроенных РП могут говорить данные, полученные при изучении различения ориентации в онтогенезе. Показано, что величины порогов для минимальных длин линий, при которых происходит грубое различение ориентации (вертикаль отличается от горизонтали), стабилизируются к 9–10 годам. Ориентационная избирательность, исследованная с помощью длинных линий разной ориентации, формируется позднее. Только в возрасте 14–16 лет она становится подобной взрослому уровню. Пороговые значения минимальных длин линий согласуются с шириной зон минимальных РП в модели, а также с шириной оптической функции рассеяния [24]. Минимальные по размеру РП находятся в стриарной коре [18], которая созревает к 8–10 годам. Приведенные здесь данные не противоречат этим срокам. Возрастные зависимости по оценке ориентационной избирательности, скорее всего, отражают взаимодействие между ориентационно настроенными РП. Эти взаимодействия реализуются на более высоком уровне зрительной системы. Поэтому стабилизация ориентационной избирательности происходит в более позднем периоде. Непосредственно сравнить развитие ориентационной чувствительности с иллюзией наклона в онтогенезе не представляется возможным из-за отсутствия таких данных. Результаты по исследованию иллюзии Поггендорфа, которую связывают с иллюзией наклона, согласуются с приведенными здесь данными. В работе [25] получено уменьшение иллюзии Поггендорфа с возрастом до 22 лет. Формирование зрительной системы носит гетерохронный характер и заканчивается примерно к 18–20 годам [26, 27]. Результаты, представленные в данном исследовании, подтверждают этот вывод.
Нейрофизиологические исследования показали, что контекст оказывает влияние на предпочитаемую ориентацию нейронов первичной зрительной коры [28]. Поэтому иллюзия наклона может происходить не из-за латерального торможения между РП нейронов или образованными ими ориентационными каналами. Результаты моделирования, проведанного в исследовании, подтверждают это предположение. Максимальный ответ дает нейрон, ориентация РП которого отличается от ориентации тестируемой части изображения. Взаимодействие же между РП может способствовать уменьшению иллюзии. В модели H.R. Wilson [29] ориентационная чувствительность, максимальная для РП с минимальным размером, повышается за счет механизма вероятностной суммации, который учитывает возбудительные взаимодействия между РП разного размера. Можно предположить, что этим же механизмом обеспечивается и уменьшение иллюзии наклона при увеличении длины линий.
ЗАКЛЮЧЕНИЕ
Уменьшение иллюзии наклона с увеличением длины, а также сопоставление возрастных зависимостей по ориентационной избирательности и порогам для минимальных длин линий с параметрами модели позволяет высказать предположение, что взаимодействия между РП способствуют уменьшению иллюзии наклона. Ее возникновение, скорее, связано с активностью нейронов с ориентационно избирательными минимальными по размеру РП.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Этическим комитетом Санкт-Петербургского государственного университета.
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа выполнена при финансовой поддержке Программы фундаментальных научных исследований государственных академий на 2013–2020 гг. (ГП-14, раздел 63).
Благодарности. Автор выражает благодарность за помощь в проведении экспериментов Л.А. Семенову.
Конфликт интересов. Автор декларирует отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Gibson J.J. Adaptation, after-effect, and contrast in the perception of tilted lines. II. Simultaneous contrast and the real restriction of the after-effect // J. Exp. Psychology. 1937. V. 20. № 6. P. 553.
Gibson J.J., Radner M. Adaptation, after-effect and contrast in the perception of tilted lines // J. Exp. Psychology. 1937. V. 20. № 5. P. 453.
Calvert J.E., Harris J.P. Spatial frequency and duration effect on the tilt illusion and orientation acuity // Vis. Res. 1988. V. 28. № 9. P. 1051.
Corbett J.E., Handy T.C., Enns J.T. When do we know which way is up? The time course of orientation perception // Vision Res. 2009. V. 49. № 1. P. 28.
Solomon J.A., Morgan M.J. Strong tilt illusions always reduce orientation acuity // Vision Res. 2009. V. 49. № 8. P. 819.
Tolhurst D.J., Thompson P.G. Orientation illusions and aftereffects: Inhibition between channels // Vision Res. 1975. V. 15. № 8–9. P. 967.
Wenderoth P., Johnson S. The different mechanisms of the direct and indirect tilt illusions // Vision Res. 1988. V. 28. № 2. P. 301.
Bouma H., Andriessen J.J. Induced changes in the perceived orientation of line segments // Vision Res. 1970. V. 10. № 2. P. 333.
Morgan M.J., Mason A.J.S., Baldassi S. Are there separate first-order and second-order mechanisms for orientation discrimination? // Vision Res. 2000. V. 40. № 13. P. 1751.
Blakemore C., Carpenter R.H.S., Georgeson M.A. Lateral inhibition between orientation detectors in the human visual system // Nature. 1970. V. 228. № 5266. P. 37.
Wenderoth P., O’Connor T., Johnson S. The tilt illusion as a function of the relative and absolute lengths of test and inducing lines // Percept. Psychophys. 1986. V. 39. № 5. P. 339.
Dakin S.C., Williams C.B., Hess R.F. The interaction of first-and second-order cues to orientation // Vision Res. 1999. V. 39. № 17. P. 2867.
Keemink S.W., van Rossum M.C.W. A unified account of tilt illusions, association fields, and contour detection based on elastica // Vision Res. 2016. V. 126. № 2. P. 164.
Wilson H.R., Gelb D.J. Modified line element theory for spatial frequency and width discrimination // J. Opt. Soc. Amer. A. 1984. V. 1. № 1. P. 124.
Бондарко В.М., Семенов Л.А. Ориентационная избирательность и острота зрения у школьников и взрослых // Физиология человека. 2017. Т. 43. № 3. С. 25. Bondarko V.M., Semenov L.A. Orientation selectivity and visual acuity in schoolchildren and adults // Human Physiology. 2017. V. 43. № 3. P. 259.
Hubel D.H., Wiesel T.N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex // J. Physiol. 1962. V. 160. № 1. P. 251.
Глезер В.Д. Зрение и мышление. Л.: Наука, 1985. 300 с.
Campbell F.W., Kulikowski J.J. Orientational selectivity of the human visual system // J. Physiology. 1966. V. 187. № 2. P. 437.
Vogels R., Orban G.A. The effect of practice on the oblique effect in line orientation judgments // Vision res. 1985. V. 25. № 11. P. 1679.
Fahle M. Specificity of learning curvature, orientation, and vernier discriminations // Vision Res. 1997. V. 37. № 14. P. 1885.
Andrews D.P. Perception of contour orientation in the central fovea // Vision Res. 1967. V. 7. № 11–12. P. 975.
Бондарко В.М., Семенов Л.А. Иллюзия жезла и рамки в зависимости от расстояния между ними // Сенс. сис. 2012. Т. 26. № 2. С. 124.
Каменкович В.М., Шевелев И.А. Латентные периоды опознания человеком геометрических фигур при разной степени маскировки их сторон и углов // Физиология человека. 2006. Т. 32. № 2. С. 5. Kamenkovich V.M., Shevelev I.A. Latency of recognition of geometrical figures with sides or corners masked to a different degree in humans // Human Physiology. 2006. V. 32. № 2. P. 127.
Шелепин Ю.Е., Бондарко В.М. Разрешающая способность и дискретизация изображений в зрительной системе // Рос. физиол. журн. им. И.М. Сеченова. 2002. Т. 88. № 9. С. 1116.
Girgus J.S., Coren S. The interaction between stimulus variations and age trends in the Poggendorff illusion // Percept. Psychoph. 1987. V. 41. № 1. P. 60.
Фарбер Д.А., Бетелева Т.Г. Формирование системы зрительного восприятия в онтогенезе // Физиология человека. 2005. Т. 31. № 5. С. 26. Farber D.A., Beteleva T.G. Formation of the System of Visual Perception in Ontogeny // Human Physiology. 2005. V. 31. № 5. P. 515.
Цехмистренко Т.А., Васильева В.А. Структурные преобразования ассоциативных зон коры больших полушарий как морфологическая основа формирования когнитивных функций мозга человека от рождения до 20 лет // Физиология человека. 2001. Т. 27. № 5. С. 41. Tsekhmistrenko T.A., Vasil’eva V.A. Structural Transformations of the Associative Cortex As the Morphological Base of the Development of Human Cognitive Functions from Birth to 20 Years of Age // Human Physiology. 2001. № 5. V. 27. P. 544.
Gilbert C.D., Wiesel T.N. The influence of contextual stimuli on the orientation selectivity of cells in primary visual cortex of the cat // Vision Rec. 1990. V. 30. № 11. P. 1689.
Wilson H.R. Responses of spatial mechanisms can explain hyperacuity // Vision Res. 1986. V. 26. № 3. P. 453.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека