

Физиология человека, 2020, T. 46, № 3, стр. 15-28
Возрастные особенности пространственно-временнóй организации ЭЭГ у испытуемых 8–30 лет мужского и женского пола при восприятии устных и письменных текстов
О. В. Кручинина 1, *, Е. П. Станкова 1, Е. И. Гальперина 1
1 ФГБУН Институт эволюционной физиологии и биохимии имени И.М. Сеченова РАН
Санкт-Петербург, Россия
* E-mail: kruchinina_ol@mail.ru
Поступила в редакцию 07.10.2019
После доработки 17.12.2019
Принята к публикации 03.02.2020
Аннотация
Созревание мозговых структур и связей между ними, а также совершенствование вербальных навыков по мере взросления являются двумя сторонами процессов формирования центральных механизмов вербальной деятельности, и оценить изолированнo вклад каждой составляющей практически невозможно. Помимо возраста существенное влияние на структурные и функциональные особенности мозга в целом и реализацию когнитивной деятельности оказывает пол. Несмотря на то, что влияние пола выражено меньше, чем возраста, его нельзя игнорировать. Задачей данного исследования было оценить отражение возрастных и половых особенностей нейрофизиологических механизмов восприятия текстов, предъявляемых на слух и зрительно, в пространственной организации биопотенциалов мозга. Пространственную синхронизацию биопотенциалов мозга (ПСБП) оценивали у 143 испытуемых (детей 8–11 лет, подростков 12–14 лет и 15–17 лет взрослых) на основе изменений интегрального показателя “объем рассеяния” (VOL) 20 ЭЭГ-векторов и степени тесноты связей ЭЭГ-сигналов. Выявлено, что с возрастом происходит постепенное увеличение пространственной синхронизации биопотенциалов мозга, как в состоянии спокойного бодрствования, так и при вербальной деятельности, при этом уровень ПСБП во всех возрастных группах при выполнении вербальных заданий ниже, чем в фоновом состоянии. Начиная с подросткового возраста проявляются половые отличия изменения уровней ПСБП при вербальной деятельности: при восприятии текста на слух – с 12 лет, при чтении текста – с 15 лет. У испытуемых мужского и женского пола показаны разные траектории формирования дистантных связей ЭЭГ, имеющие зональную специфичность для исследованных видов вербальной деятельности.
Одним из наиболее актуальных вопросов современной когнитивной нейрофизиологии является изучение формирования центральных механизмов вербальной деятельности на протяжении детского и подросткового возраста [1–8]. Структурные и функциональные изменения мозга сочетаются с процессами становления изучаемых когнитивных навыков, при этом дефинитивный возраст формирования различных вербальных навыков сильно отличается, а иногда имеет также половые особенности и темповые характеристики формирования. Невозможность изолировать эти факторы в онтогенезе затрудняет точную оценку вклада каждого из них.
Гетерохронное созревание высших психических функций позволяет предполагать наличие различных мозговых механизмов вербальной деятельности в каждом возрасте. Остается до конца не выясненным вопрос: реализация вербальной деятельности осуществляется за счет специфических для детского, подросткового или взрослого возраста мозговых механизмов, либо по мере возрастного развития происходит исключительно тонкая настройка и совершенствование/оптимизация уже заложенных механизмов. Так, существует мнение, что дети (4–12 лет) задействуют те же базовые корковые области языковой сети, что и взрослые [7], при этом не отрицается влияние возраста, задачи и методологии исследования на нейронную активацию. С другой стороны, современные данные трактографии показали, что процесс миелинизации проводящих путей, образующих основу структурной связности мозга, не заканчивается в подростковом возрасте, как это считалось ранее, а продолжается вплоть до 25-летнего возраста и даже далее [9–11], кроме того, зависит от пола [12] и изменяется благодаря синаптической пластичности в процессе становления навыка [13]. Эти факторы значимо влияют на мозговые механизмы и не исключают появления специфических для каждого возраста вариантов реализации вербальной деятельности, наиболее адекватных для каждого этапа развития.
Центральное обеспечение когнитивной деятельности опосредуется сложно скоординированной работой множества пространственно распределенных мозговых структур. Оценка функциональной связности (functional connectivity) различных зон мозга производится с использованием методов функциональной МРТ [12, 14, 15], МЭГ и ЭЭГ [16, 17]. Один из способов исследования координированного взаимодействия структур в работе мозга – анализ пространственных отношений колебаний биопотенциалов мозга методом оценки кросскорреляционных и когерентных связей ЭЭГ-процессов [18–20].
Степень сформированности интегративных процессов, обеспечивающих целостную деятельность мозга, создает необходимые предпосылки для развития когнитивных навыков в определенном возрасте. В то же время созревание различных мозговых структур отличается у юношей и девушек в среднем на 1–2 года [10, 11, 21]. Кроме того, описаны половые особенности перестройки фоновой ЭЭГ пубертатного периода [20, 22] и наличие гендерных отличий в пространственной организации мозговой активности, связанной с обеспечением разных видов когнитивной деятельности [20, 23, 24]. Различия функциональной связности мозга у мужчин и женщин проявляются в том числе при восприятии речи [25–31]. Критическим для формирования таких различий является период полового созревания [32, 33]. Показано, что именно в это время происходит интенсивное формирование межрегионального взаимодействия корковых и подкорковых структур [34, 35]. Может создаваться впечатление, что половые различия реализации вербальной деятельности пубертатного периода исчерпываются темповыми характеристиками. Однако мальчики и девочки могут иметь различные траектории развития, поскольку есть данные, показывающие, что с возрастом не происходит нивелирования половых отличий [36].
Эволюционно и онтогенетически устная речь появляется у человека раньше, чем письменная. Мозг младенца избирательно преднастроен на обработку слуховой информации, специфичной для речи [36–38]. Тогда как чтение – это навык, которому целенаправленно обучают. Вероятно, нейрональные механизмы восприятия устной и письменной речи различны, при этом механизмы устной речи могут быть более упорядоченными и автоматическими, чем письменной [7, 39]. Физиологические различия восприятия речи на слух и чтения могут быть обусловлены разными сенсорными входами, тогда как процессы понимания могут опираться на модально неспецифичные системы [8]. С одной стороны, чтение может осуществляться на основе устной речи – перевода информации из зрительной модальности в слуховую (графемы в фонему), что приводит к активации planum temporale с прилегающими участками верхней височной борозды и извилины [40], при этом для понимания прочитанного задействуется та же система связей, что и при анализе устной речи. Однако существует как минимум еще один механизм чтения текста, не задействующий слуховую систему [41]: прямой путь обработки прочитанного, в котором участвует область графического образа слова (visual word form area) [42]. В обработке знакомых и коротких слов, а также т.н. словарных слов, написание которых не соответствует их звуковой структуре, участвует в основном область графического образа слова, а длинных и незнакомых слов – planum temporale, поскольку для распознавания слова требуется соотнесение с его фонетическим образом. Возможно, изменение этих механизмов происходит под действием задачи по мере взросления, совершенствования навыка и пр.
Возникает сложная проблема взаимосвязи многих факторов (возраста и пола испытуемых, модальности вербальной задачи и пр.) и их влияния на системное взаимодействие различных отделов коры при выполнении вербальной деятельности. На решение этой проблемы направлено данное исследование, целью которого стало описание траектории становления пространственных взаимодействий биопотенциалов мозга, отражающих нейрофизиологические механизмы чтения и восприятия устного текста у испытуемых мужского и женского пола.
МЕТОДИКА
В исследовании принимали участие 143 испытуемых в возрасте от 8 до 30 лет: дети 8–11 лет (средний возраст 10.1 ± 1.2, n = 21.8 мальчиков), подростки 12–14 лет (средний возраст 13.3 ± 1.1, n = 41.19 юношей), подростки 15–17 лет (средний возраст 16.5 ± 1.3, n = 35.17 юношей) и взрослые испытуемые (средний возраст 23.1 ± 5.1, n = = 46.20 мужчин). Все испытуемые были правши (оценено по модифицированному тесту Annett) [43]. Испытуемые не имели неврологических заболеваний и речевых нарушений в анамнезе, не принимали лекарственных препаратов и не имели жалоб на здоровье на момент обследования.
Стимульный материал. С целью выявления возрастных особенностей системной организации мозга при когнитивной деятельности испытуемым предлагали задания на восприятие устного и письменного текстов. Стимульный материал подбирали, исходя из возраста испытуемых и соответственно определенным критериям: семантически завершенный отрывок литературного текста описательного характера длиной 400–450 слов, преимущественно эмоционально нейтральный, новый для испытуемых. Текст для чтения предъявляли на мониторе компьютера, расположенном в 1 м от испытуемого. Испытуемого не ограничивали во времени прочтения текста, он сам извещал экспериментатора об окончании выполнения задания, после чего монитор выключали. Текст для восприятия на слух предъявляли через наушники. После прочтения/прослушивания отрывка испытуемым необходимо было ответить на 10 вопросов по содержанию текста для оценки понимания прочитанного/прослушанного, процент ошибочных ответов или отсутствия ответа на вопрос также не превышал 10%.
ЭЭГ-исследование. ЭЭГ непрерывно регистрировали в фоне (спокойное бодрствование с закрытыми и открытыми газами), а также при восприятии текста на слух с закрытыми глазами и при чтении текста на 24-канальном компьютерном электроэнцефалографе (Brain Dynamics analyzer, Россия) с полосой пропускания – 0.5–30 Гц, с частотой дискретизации 250 Гц по каждому из каналов. Запись производили от 20 монополярных отведений, из них 16 располагали по международной схеме 10–20 (Fp1, Fp2, F3, F4, F7, F8, C3, C4, T3, T4, T5, T6, Р3, Р4, O1, O2). Дополнительно устанавливали четыре электрода: два – в передневисочных областях каждого из полушарий (T1, T2) и два – в зонах ТРО, т.е. в зонах перекрытия височной, теменной и затылочной областей (Tp1, Tp2). В качестве референтного отведения использовали объединенные электроды на мочках ушей. Для регистрации движений глаз и морганий использовали биполярное отведение с расположением электродов в районе височного угла глаза и верхнего края глаза.
ЭЭГ в диапазоне 1.6–30 Гц после удаления артефактов разделяли на эпохи анализа по 4 с, для каждой эпохи анализа вычисляли матрицы коэффициентов кросскорреляции (КК) ЭЭГ от всех отведений попарно (всего 190 значений). Для определения уровня общей пространственной синхронизации биопотенциалов мозга (ПСБП) и особенностей топической организации ПСБП матрицы КК ЭЭГ анализировали методом “объемов” [44]. В основе метода лежит представление многоканальной ЭЭГ в виде совокупности векторов b1, …, bn единичной длины в многомерном евклидовом пространстве, где каждый из векторов соответствует локальному ЭЭГ-процессу, а косинус угла между двумя векторами есть КК между локальными процессами. Иными словами, матрица КК ЭЭГ A = (aij), 1 ≤ i, j ≤ n представляет собой матрицу попарных скалярных произведений векторов b1, …, bn и aij = (bi, bj). В нашем случае n = 20, что соответствует числу отведений ЭЭГ. Производили расчет объема d-мерного параллелепипеда, натянутого на вектора bi1, …, bid. Для получения более статистически устойчивых оценок параметра VOL, объем параллелепипеда рассчитывали не в полномерном (d = n = 20) пространстве, а при d = 4, таким образом, что величина VOL соответствовала среднему из n!/(d !(n – d)!) объемов. Объем параллелепипеда, соответствующего каждой подматрице матрица КК ЭЭГ A = (aij), 1 ≤ i, j ≤ d, пропорционален квадратному корню значения определителя подматрицы А. В целях оптимизации вычислительной процедуры параметр VOL вычисляли по формуле: VOL = ($\bar {1}$)d– 1TrBd, где Вd – присоединенные матрицы для подматриц Аd, которые рассчитывали по методу Д.К. Фаддеева [45].
Чем больше скоррелированы сигналы, тем меньше углы между векторами, тем меньший объем занимает пучок ЭЭГ-векторов в пространстве. Таким образом, “объем” (VOL), занимаемый пучком ЭЭГ-векторов в пространстве, иначе говоря, показатель линейной зависимости ЭЭГ-сигналов от всех отведений, выступает в качестве меры ПСБП. Если VOL = 0, то процессы максимально линейно зависимы, если VOL = 1, то процессы линейно независимы (векторы пучка попарно ортогональны). Кроме того, вычисляли показатели, характеризующие “объемы” пучков 10 векторов в каждом из полушарий мозга – для правого (VOL-RH) и левого (VOL-LH) полушарий.
Для оценки особенностей топической организации ПСБП для каждого из ЭЭГ-отведений вычисляли показатель V(i) (где i – порядковый номер отведения) – “долю объема” пучка векторов, приходящуюся на i-й вектор. Величины V(i) пропорциональны диагональным элементам матрицы Bd. Величина V(i) характеризует степень отличия i-го ЭЭГ-сигнала от совокупности остальных: чем больше V(i), тем больше отличие.
Картина ЭЭГ, а, следовательно, и показатель VOL, сильно изменяются в зависимости от наличия зрительного сенсорного потока. В данном исследовании при восприятии текста на слух испытуемый находился с закрытыми глазами, а при чтении – с открытыми, поэтому особенности ПСБП мозга при вербальной деятельности, оцениваемые как по интегративному показателю VOL, так и по долям зон V(i), рассматривали по отношению к соответствующему фоновому состоянию: аудирование сравнивали с фоном, зарегистрированным при закрытых глазах, а чтение – с состоянием спокойного бодрствования, зарегистрированным при открытых глазах.
Статистический анализ производили в пакете программ Statistica‑12 for Windows. Межгрупповые сравнения ПСБП проводили с помощью дисперсионного анализа (ANOVA) c учетом факторов “Возраст” (4 возрастные группы), “Задание” (наличие или отсутствие вербальной деятельности, данный фактор оценивали для прослушивания текстов и чтения отдельно) и “Пол”. Уровень значимости принимали равным 0.05. Апостериорный анализ проводили с помощью теста Фишера, применяли поправку Бонферони для корректировки уровня значимости гипотезы на количество сравнений. В тексте приведены средние значения показателей ПСБП и 95% доверительный интервал для ошибки среднего.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Изучение пространственно-временнóй организации биопотенциалов мозга при выполнении вербальных заданий детьми 8–11 лет, подростками 12–14 и 15–17 лет и взрослыми 18–30 лет выявило возрастные, половые и связанные с выполнением заданий особенности перестройки межцентральных взаимодействий биопотенциалов мозга. Рассматривали влияние перечисленных факторов на величину общего “объема рассеяния” ЭЭГ-векторов, “объема рассеяния” ЭЭГ-векторов отдельно для левого и для правого полушарий мозга, а также локального уровня ПСБП, оцениваемого по показателям долей зон V(i).
Выявлено, что фактор “Возраст” (4 градации: 8–11, 12–14, 15–17, 18–30 лет) оказывал достоверное влияние на величину показателя VOL, зарегистрированного в состоянии спокойного бодрствования с закрытыми (F(3, 104) = 46.1, p < < 0.00001) и открытыми глазами (F(3, 127) = 32.2, p < 0.00001), а также при выполнении каждого из вербальных заданий: восприятии текста на слух (F(3, 104) = 28.8, p < 0.00001) или чтении текста (F(3, 127) = 25.3, p < 0.00001). В частности, от детского к взрослому возрасту происходит снижение показателя VOL, характеризующего степень линейной взаимосвязанности ЭЭГ-сигналов от всей совокупности отведений (табл. 1). Поскольку показатель “объем рассеяния” ЭЭГ-векторов обратно пропорционален уровню пространственной синхронизации ЭЭГ, т.е. меньшие значения величины VOL свидетельствуют о большем уровне ПСБП мозга, следовательно с возрастом происходит увеличение пространственной синхронизации биопотенциалов мозга. Выявленная динамика изменения уровня ПСБП наблюдается как в состоянии спокойного бодрствования с закрытыми и открытыми глазами, так и при вербальной деятельности – восприятии текста на слух или чтении.
Таблица 1.
Уровень пространственной синхронизации ЭЭГ, оцениваемый по величине “объема рассеяния” ЭЭГ-векторов, у детей и взрослых при восприятии текстов и в фоне
| VOL | 8–11 лет | 12–14 лет | 15–17 лет | 18–30 лет | ||||
|---|---|---|---|---|---|---|---|---|
| ср. зач. | 95% доверит. для ош. ср. | ср. зач. | 95% доверит. для ош. ср. | ср. зач. | 95% доверит. для ош. ср. | ср. зач. | 95% доверит. для ош. ср. | |
| Закрытые глаза | 0.310 | ±0.003 | 0.280 | ±0.002 | 0.212 | ±0.002 | 0.204 | ±0.003 |
| Восприятие на слух | 0.326 | ±0.005 | 0.294 | ±0.003 | 0.250 | ±0.003 | 0.228 | ±0.005 |
| Открытые глаза | 0.335 | ±0.008 | 0.293 | ±0.003 | 0.280 | ±0.004 | 0.277 | ±0.002 |
| Чтение текста | 0.381 | ±0.007 | 0.308 | ±0.003 | 0.297 | ±0.004 | 0.290 | ±0.003 |
Вторым фактором, оказывающим существенное влияние на величины общего объема рассеяния ЭЭГ-векторов, VOL-LH и VOL-RH (табл. 2, А) и локального уровня ПСБП (оцениваемого по показателям долей зон V(i)), является фактор “Задание”. Поскольку картина ЭЭГ, а, следовательно, и показатель VOL, сильно изменяются в зависимости от наличия зрительного сенсорного потока, а выполнение двух тестовых заданий различалось по этому параметру (при восприятии текста на слух испытуемый находился с закрытыми глазами, а при чтении – с открытыми), то не сравнивали между собой показатели VOL, вычисленные при выполнении разных вербальных заданий, а сравнивали только электрическую активность мозга при выполнении задания с соответствующим этому заданию фоновым состоянием. Таким образом, фактор “Задание” имел две градации: фоновое состояние и вербальную деятельность.
Таблица 2.
Влияние факторов “Задание”, “Пол” и взаимодействие этих факторов на показатели общего “объема рассеяния” ЭЭГ-векторов
| Фактор “Задание” (А) | 8–11 лет | 12–14 лет | 15–17 лет | Взрослые | |||||
|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | ||
| Фон ЗГ
и восприятие на слух |
VOL | 11.56 | 0.00 | 16.47 | 0.00 | 86.12 | 0.00 | 7.28 | 0.01 |
| LH | 20.79 | 0.00 | 18.15 | 0.00 | 76.84 | 0.00 | 16.00 | 0.00 | |
| RH | 11.85 | 0.00 | 8.03 | 0.00 | 58.59 | 0.00 | 4.48 | 0.03 | |
| Фон ОГ и чтение | VOL | 22.96 | 0.00 | 14.93 | 0.00 | 10.23 | 0.00 | 22.06 | 0.00 |
| LH | 23.29 | 0.00 | 12.14 | 0.00 | 4.41 | 0.04 | 16.49 | 0.00 | |
| RH | 15.38 | 0.00 | 3.68 | 0.06 | 20.67 | 0.00 | 22.18 | 0.00 | |
| Фактор “Пол” (Б) | F | p | F | p | F | p | F | p | |
| Фон ЗГ
и восприятие на слух |
VOL | 5.89 | 0.02 | 64.99 | 0.00 | 46.60 | 0.00 | 5.93 | 0.02 |
| LH | 0.17 | 0.68 | 12.55 | 0.00 | 30.11 | 0.00 | 72.81 | 0.00 | |
| RH | 2.11 | 0.15 | 114.84 | 0.00 | 48.17 | 0.00 | 4.32 | 0.04 | |
| Фон ОГ и чтение | VOL | 7.41 | 0.01 | 0.02 | 0.90 | 20.14 | 0.00 | 132.94 | 0.00 |
| LH | 8.52 | 0.00 | 7.63 | 0.01 | 14.85 | 0.00 | 104.53 | 0.00 | |
| RH | 12.10 | 0.00 | 20.61 | 0.00 | 6.27 | 0.01 | 123.61 | 0.00 | |
По данным двухфакторного дисперсионного анализа, был выявлен значимый эффект фактора “Задание” (2 градации: фон – вербальное задание) на величину показателя VOL (при р < 0.01), характеризующего общий уровень ПСБП, во всех возрастных группах (табл. 2, А). Кроме того, при вербальной деятельности (как при восприятии текста на слух, так и при чтении) у всех испытуемых отмечаются более высокие значения показателя VOL, чем в фоновом состоянии (закрытые или открытые глаза), т.е. выполнение вербальных заданий вызывает достоверное снижение общего уровня ПСБП мозга (десинхронизацию) (табл. 1).
При прослушивании текста VOL-LH достоверно (при р < 0.001) отличался от фоновых значений во всех возрастах, а при чтении текста – во всех возрастных группах, кроме подростков 15–17 лет. Статистически значимые отличия (при р < 0.001) ПСБП в правом полушарии (VOL-RH) от фонового уровня при прослушивании выявлены во всех возрастах, кроме взрослого, а при чтении – во всех возрастах, кроме подростков 12–14 лет. Таким образом, результаты дисперсионного анализа показали, что выполнение вербальной деятельности оказывает значимое влияние на интегральные показатели ПСБП мозга.
Если обратиться к анализу тесноты связей отдельных зон мозга с совокупностью остальных, оцениваемых по показателю V(i), можно обнаружить значительные отличия в организации деятельности мозга во время восприятия устной и письменной речи. Специфическая для каждого вида вербальной деятельности картина изменений уровня ПСБП прослеживается у детей, подростков 12–14 и 15–17 лет и взрослых. На рис. 1 показаны изменения величины V(i), характеризующей степень участия корковых областей в процессах пространственной синхронизации при вербальной деятельности. Возрастание величины V(i) свидетельствует об увеличении выраженности локальной активности и снижении доли пространственно-синхронных процессов при выполнении вербальных заданий по сравнению с фоном в данной области коры мозга. И наоборот, уменьшение величины V(i) свидетельствует о снижении выраженности локальной активности и повышении вклада пространственно-синхронных процессов.
Рис. 1.
Топические особенности пространственных взаимосвязей ЭЭГ у детей, подростков 12–14 и 15–17 лет и взрослых при восприятии на слух текста по сравнению со спокойным бодрствованием с закрытыми глазами (А), при чтении текста по сравнению со спокойным бодрствованием с открытыми глазами (Б) (по показателям вклада зон V(i)). Диаметр кружка пропорционален величине критерия Фишера для апостериорных сравнений, серый кружок – увеличение V(i) при деятельности по сравнению с фоном, черный – уменьшение V(i). Уровень значимости различий: * – p < 0.01, p < 0.001.
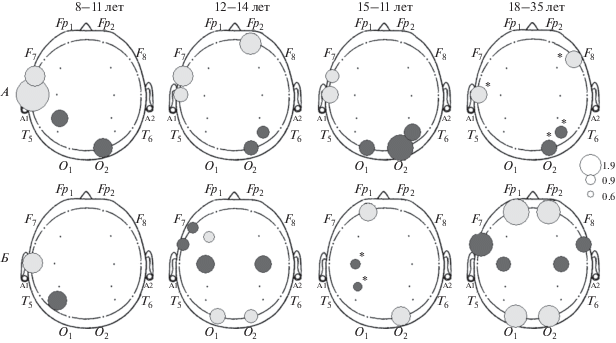
При восприятии текстов на слух, по сравнению с фоновым состоянием спокойного бодрствования с закрытыми глазами, наблюдается увеличение показателя V(i) в зонах Т1, Т3 левого полушария и снижение – в О2, Тpo2 правого полушария (рис. 1, А), т.е. вклад пространственно-асинхронной (локальной) составляющей увеличивается в височных областях левого полушария и снижается в затылочно-теменных областях правого полушария.
При чтении текста, по сравнению с фоновым состоянием спокойного бодрствования с открытыми глазами, отмечено уменьшение степени тесноты связей Fp1, Fp2, O1, O2 зон и увеличение – С3, С4 со всеми остальными (рис. 1, Б), т.е. вклад пространственно-асинхронной (локальной) составляющей увеличивается во фронтальных и затылочных областях и снижается в центральных зонах обоих полушарий.
Кроме того, для каждого вида вербальной деятельности можно отметить свою возрастную динамику межцентральных взаимодействий. По мере взросления уменьшается выраженность изменений биоэлектрической активности мозга относительно фонового уровня при восприятии речи на слух (рис. 1, A), в то же время при чтении, по сравнению с фоном, происходит усиление степени выраженности изменений и возрастание вклада пространственно-асинхронной составляющей лобных (Fp1, Fp2) и затылочных (О1, О2) областей (рис. 1, Б). С возрастом паттерн межрегиональных взаимодействий, характеризующий процесс чтения, проявляется все более отчетливо. Если у детей самой младшей возрастной группы (8–11 лет) характер изменений ПСБП при чтении, по сравнению с фоном, напоминает таковой при восприятии текста на слух, по сравнению с соответствующим фоновым состоянием (рис. 2, А, Б – 8–11 лет), а именно увеличение V(i) в височных и уменьшение – в теменных областях левого полушария, то уже с 12–14 лет начинает формироваться специфический для чтения паттерн активации (увеличение степени тесноты связей (V(i)) затылочных областей и уменьшение – центральных билатерально и передневисочных левого полушария). Возможно, наблюдаемая возрастная динамика отражает становление навыка чтения, переход его на более сложный уровень.
Рис. 2.
Половые различия показателей общего “объема рассеяния” ЭЭГ-векторов (VOL) в фоне (А – закрытые глаза и В – открытые глаза), а также восприятии на слух (Б) и чтении (Г) текстов детьми, подростками двух возрастных групп и взрослыми. а – мальчики и мужчины (n = 64), б – девочки и женщины (n = 79). По оси абсцисс представлены 4 группы испытуемых. По оси ординат – показатель уровня пространственной синхронизации ЭЭГ, рассчитанный по “объемам рассеяния” векторов ЭЭГ в относительных единицах, вертикальные отрезки – 95% доверительные интервалы для ошибки среднего.
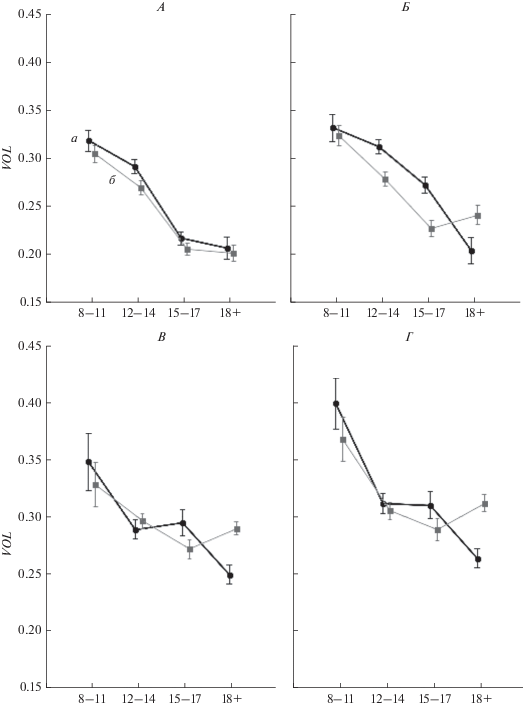
Третьим фактором, оказывающим наряду с “Возрастом” и “Заданием” значимое влияние на организацию электрической активности мозга, является пол испытуемых. По данным дисперсионного анализа, фактор “Пол” оказывал значимое влияние на показатель общего объема рассеяния ЭЭГ-векторов (VOL) (при р < 0.0001), зарегистрированных у подростков от 12 до 17 лет при восприятии текстов на слух (рис. 2, Б), а также на ПСБП во время спокойного бодрствования с открытыми глазами и чтения у испытуемых 15–17 лет и взрослых (рис. 2, В, Г). Несмотря на то, что общий возрастной тренд – снижение уровня VOL от детского к взрослому возрасту – проявляется как у мужчин, так и у женщин во всех функциональных состояниях, с возрастом при выполнении вербальной деятельности начинают появляться половые отличия уровня ПСБП, в большей степени эти отличия выражены при восприятии текста на слух (рис. 2, Б). При этом в фоновом состоянии общий уровень ПСБП мозга у мужчин и у женщин практически одинаков, различия обнаружены только в состоянии спокойного бодрствования с открытыми глазами у взрослых (рис. 2, В). Таким образом, по мере возрастного развития половые отличия изменения уровня ПСБП проявляются в большей степени при вербальной деятельности – при восприятии текста на слух (рис. 2, Б), начиная с подросткового возраста (с 12 лет), при чтении текста, начиная с 15 лет (рис. 2, Г). В фоновом состоянии с открытыми глазами половые отличия проявляются больше (рис. 2, В), чем с закрытыми глазами, при котором возрастная динамика изменения ПСБП практически идентична у мужчин и женщин (рис. 2, А).
Следует также отметить, что траектория развития (developmental trajectory) функциональной связности мозга, оцениваемая по показателям ПСБП мозга у мужчин и женщин, различается. У испытуемых женского пола, по мере возрастного развития, происходит постепенное увеличение уровня ПСБП (снижение показателя VOL) от детского к подростковому возрасту 15–17 лет и небольшое снижение уровня ПСБП во взрослом возрасте (рис. 2) во всех исследованных состояниях. В отличие от женского пола, у испытуемых мужского пола возрастная динамика изменения уровня ПСБП различается для состояний с открытыми и закрытыми глазами. В состояниях с закрытыми глазами уровень ПСБП мозга постепенно увеличивается от одной возрастной группы к другой (монотонное снижение кривой VOL). В состояниях с открытыми глазами (как в фоне, так и при чтении) уровень ПСБП мозга нарастает от детского к подростковому возрасту, не изменяется на протяжении подросткового возраста, а потом опять увеличивается у взрослых. Таким образом, в подростковом возрасте половые отличия ПСБП проявляются наиболее отчетливо, т.е. можно предположить, что подростковый возраст является определяющим для формирования половых отличий.
Помимо возрастной динамики изменений уровня ПСБП мозга, отличия у испытуемых разного пола проявляются еще и в степени, т.е. абсолютном значении изменений. Более существенные изменения уровня ПСБП от детского к взрослому возрасту наблюдаются у мужчин, чем у женщин. Это характерно для состояний с открытыми глазами, например, при чтении у мужчин показатель общего “объема рассеяния” ЭЭГ-векторов VOL изменяется от 0.4 в детском до 0.26 во взрослом возрасте, а у женщин только – от 0.37 до 0.31 (рис. 2, Г).
Поскольку при выполнении вербальных заданий были выявлены половые различия возрастной динамики изменения общего “объема рассеяния” ЭЭГ-векторов, т.е. уровня ПСБП (рис. 2, табл. 2, Б), решили более детально рассмотреть топические особенности становления речевой функции у испытуемых разного пола по мере взросления. По данным апостериорного дисперсионного анализа с использованием поправки Бонферрони, было выявлено значимое влияние фактора “Пол” на показатели вклада зон V(i), позволяющих оценить уровень взаимосвязанности колебаний биопотенциалов в каждом отведении ЭЭГ со всеми остальными. Достоверные отличия при p < 0.001 обнаружены в 14 отведениях ЭЭГ при восприятии текста на слух (Fp1, Fp2, F4, F8, T1, T2, T3, T4, T5, T6, C3, C4, O1, O2) и в 8 – при чтении (Fp2, F8, T1, T3, T5, C4, Tpo2, O1) (рис. 3). Отведения ЭЭГ, в которых были выявлены половые отличия, схожи для обоих видов вербальных заданий, но при восприятии текстов на слух этих отведений значительно больше.
Рис. 3.
Возрастная динамика изменений величины V(i), характеризующей степень участия корковых областей в процессах пространственной синхронизации при восприятии на слух (А) и чтении текстов (Б) испытуемыми мужского (а) и женского (б) пола. а – мужчины (n = 64), б – женщины (n = 79). I – 8–11 лет, II – 12–14 лет, III – 15–17 лет, IV – 18–30 лет. Вертикальные отрезки – 95% доверительные интервалы для ошибки среднего. Представлены отведения, для которых был выявлен значимый эффект фактора “Пол” по результатам апостериорного анализа при р < 0.001 (Бонферрони).
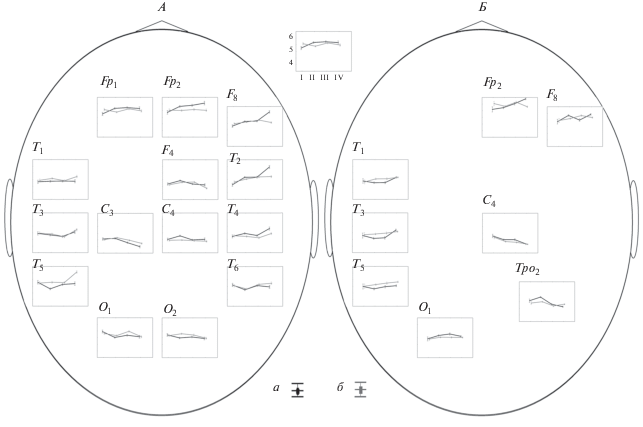
Половые отличия наиболее ярко проявляются у взрослых при восприятии текстов на слух (F8, T1, T2, T4, T5, T6) (рис. 3, А) и у подростков от 12 до 17 лет при чтении (Т1, Т3, Т5, F8, О1) (рис. 3, Б). В целом, при выполнении вербальных заданий показатель V(i) выше у испытуемых женского пола, чем мужского, преимущественно в отведениях левого полушария (в височных – Т1, Т3, Т5, Т6, центральной – С3 и затылочных – О1, О2 областях мозга), а у испытуемых мужского пола – преимущественно правого (фронтальных – Fp1, Fp2, F4, F8, центральной – С4 и височной – Т2, Т4 областей мозга).
На рис. 3 представлена возрастная динамика изменения показателя тесноты связей зон V(i) с совокупностью остальных у испытуемых разного пола при восприятии текстов на слух и чтении. Для одних зон мозга характер изменений с возрастом одинаков у испытуемых обоих полов, при этом в каждом возрасте наблюдаются достоверные половые различия по значениям показателя V(i) (например, фронтальные и затылочные области мозга). В других зонах выявлена различная возрастная динамика изменения значения показателей V(i). В височных областях левого полушария (Т1, Т3, Т5) при восприятии текстов на слух у испытуемых мужского пола значение показателя V(i) остается стабильным (V(i) = 4.9 ± 0.2) во всех возрастных группах, в то время как у испытуемых женского пола этот показатель практически не изменяется на протяжении детского и подросткового возраста (V(i) = 5 ± 0.1), а затем значительно возрастает у взрослых особенно в Т5, где V(i) = 6 (рис. 3, А). Возрастание величины V(i) свидетельствует об увеличении выраженности локальной активности и снижении доли пространственно-синхронных процессов. При этом, при чтении в тех же височных областях левого полушария у испытуемых женского пола наблюдается незначительное монотонное нарастание значения V(i) по мере взросления, а у испытуемых мужского пола – резкое изменение показателей V(i) при переходе от подросткового к взрослому возрасту (рис. 3, Б). В височных зонах правого полушария (T2, T4) и нижнелобной области (F8) справа при восприятии на слух выявлена следующая возрастная динамика изменения показателей V(i): у испытуемых женского пола наблюдается незначительное монотонное нарастание значения V(i), а у испытуемых мужского пола – резкое изменение показателей V(i) от детского к подростковому возрасту (12–14 лет), а затем от подросткового (15–17 лет) к взрослому (рис. 3, А).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Исследование пространственно-временнóй организации биопотенциалов мозга при выполнении вербальных заданий детьми, подростками 12–14 и 15–17 лет и взрослыми выявило зависимость перестройки межцентральных взаимодействий от таких факторов как возраст, пол испытуемых, а также специфика выполняемого вербального задания.
Возраст. В данном исследовании показано постепенное увеличение по мере возрастного развития ПСБП, наблюдаемое как в состоянии спокойного бодрствования с закрытыми и открытыми глазами, так и при двух видах вербальной деятельности: восприятии текста на слух и при чтении. Изменения уровня ПСБП оценивали по показателю “объема” рассеяния ЭЭГ-векторов (VOL) (рис. 1 и 2) у испытуемых от 8 до 30 лет (n = = 143, 64 мужского пола). Показатель “объем рассеяния” ЭЭГ-векторов (VOL) характеризует степень линейной взаимосвязанности колебаний потенциалов во всей совокупности отведений ЭЭГ, его величина обратно пропорциональна уровню пространственной синхронизации ЭЭГ, т.е. меньшие значения величины VOL свидетельствуют о большем уровне ПСБП мозга. Полученные результаты подтверждают данные литературы о становлении пространственной синхронизации ЭЭГ с возрастом, которая оценивалась с использованием расчета когерентности ЭЭГ-сигнала [34], коэффициентов корреляции ЭЭГ-процессов [19, 24] и “объема” рассеяния ЭЭГ-векторов [22, 46, 47]. Ранее было показано, что по мере возрастного развития происходит увеличение уровня пространственной синхронизации биопотенциалов в состоянии спокойного бодрствования с закрытыми глазами (от 4 до 30 лет), а также при восприятии на слух текстов на русском и английском языках [24, 46]. Выполнение вербальной нагрузки с открытыми глазами – чтение текста (рис. 1) согласуется с этими данными. Поэтому можно предположить наличие более общих, не зависящих от специфики задания, мозговых механизмов, обеспечивающих интеграцию нервных процессов как в фоне, так и при деятельности, и действующих одинаково эффективно при нагрузках, выполняемых с закрытыми и открытыми глазами.
Задания. Выявлена специфика перестройки межрегиональных взаимодействий биопотенциалов мозга для каждого вида вербальной деятельности, по сравнению с соответствующим фоновым состоянием.
Во-первых, выполнение вербальных заданий вызывает снижение общего уровня пространственной синхронизации биоэлектрической активности мозга по сравнению с фоновым состоянием во всех исследованных группах испытуемых (увеличение показателя VOL) (рис. 2). Снижение уровня ПСБП мозга характеризует увеличение доли локальной активности и, следовательно, возрастание степени дифференцированного участия различных зон коры в обеспечении когнитивных процессов. Когнитивная деятельность требует соотношения процессов регулируемой локальной активации, создающих структурную основу объединений [48, 49], а также процессов межцентральной интеграции, обеспечивающих взаимодействие в этой распределенной системе [24, 50]. Несмотря на кажущееся противоречие между процессами локализации и системной интеграции, большинство исследователей соглашаются с тем, что целенаправленная деятельность обеспечивается одновременно селективным вовлечением в работу различных систем мозга, повышением активности одних и снижением других зон коры, а также избирательной организацией синергических и реципрокных связей между ними [22, 34].
Во-вторых, наблюдаются специфические для каждого задания (восприятие на слух или чтение текстов по сравнению с соответствующим фоном) паттерны изменений структуры пространственного взаимодействия ЭЭГ (рис. 1). Так при восприятии текстов на слух, по сравнению с фоновым состоянием, увеличивается вклад пространственно-асинхронной (локальной) составляющей в височных областях левого полушария и снижается в затылочно-теменных областях правого полушария (рис. 1, А). Специфическая активация верхней височной извилины левого полушария показана также методами нейровизуализации (фМРТ) при обработке воспринимаемых на слух речевых стимулов [42, 51].
По мере возрастного развития специфический паттерн активации мозга при восприятии текста на слух, по сравнению с фоном, сохраняется, но степень его выраженности с возрастом падает, что подтверждает ранее высказанную гипотезу о возрастной минимизации функций [52]. Было показано, что у детей при когнитивной деятельности (прослушивание и запоминание стихотворений, корректурные пробы и др.) по сравнению с фоном, наблюдается повышенная, по сравнению со взрослыми генерализация изменений исходного паттерна межцентральных отношений [52]. Кроме того, у детей, по сравнению со взрослыми, крупномасштабные нейронные сети (large-scale networks) более диффузны и имеют менее жесткие границы [53, 54] и, вероятно, при переходе к состояниям, обусловленным внешними воздействиями, в работу вовлекается больше областей, и, следовательно, изменения являются более генерализованными [55].
Выявленные различия в степени изменения вклада долей зон при восприятии текстов на слух и при чтении, по сравнению с фоном, могут быть обусловлены тем, что для чтения показано привлечение более широких нейроанатомических путей, что отражает более активную обработку и больший набор когнитивных требований, необходимых для чтения, по сравнению с прослушиванием [7].
При чтении текста, по сравнению с фоновым состоянием, увеличивается локальная активация фронтальных и затылочных областей и снижается – центральных зон обоих полушарий, причем по мере возрастного развития паттерн межрегиональных взаимодействий, характеризующий процесс чтения на уровне текста, проявляется все более отчетливо (рис. 1, Б). Так называемая “сеть чтения” обычно включает три основных региона: левую затылочно-височную, височно-теменную и нижнелобную области [56]. Кроме того, при чтении по сравнению с покоем описана активация затылочных и задних вентральных височных областей обоих полушарий, а также лобных и теменных областей [7, 57, 58]. Ранее полагали, что вовлечение гомологичных областей обоих полушарий мозга во время чтения является необходимым условием для повышения производительности в процессе становления навыка [7], в нашем исследовании при чтении текстов по сравнению с фоном задействованы гомологичные отведения правого и левого полушария мозга у взрослых (рис. 1, Б). Вероятно, подобный паттерн активации характерен для чтения вообще, а не является отражением степени сформированности навыка. Это согласуется с данными о привлечении более широких нейроанатомических путей при чтении по сравнению с прослушиванием [58].
У детей младшей возрастной группы 8–11 лет характер межцентральных взаимодействий при восприятии и на слух и при чтении по сравнению с фоном имеют сходную структуру, а именно отмечается увеличение вклада височных отделов левого полушария и снижение теменно-височных. Такого сходства в более старших возрастных группах не наблюдается (рис. 1), поэтому допустимо предположить изменение механизма чтения по мере совершенствования этого навыка, в частности, может снижаться опора на фонологический образ прочитанного слова, преимущественно использоваться прямой доступ к хранилищу зрительных образов слов и происходить прямое соотнесение с семантикой. Это предположение согласуется с теорией двух потоков обработки письменной речи [59]. Кроме того, языковая латерализация в слуховой модальности происходит относительно рано, еще у детей дошкольного возраста, в то время как латерализация лингвистических процессов, участвующих в чтении, развивается медленнее, параллельно с обучением чтению [60].
Пол. Половые различия существенно выше при деятельности, чем в состоянии спокойного бодрствования. Вероятно, у мужчин и женщин существует специфика реализации вербальной деятельности, которую признают даже сторонники теории “гендерного равенства” (Gender Similarities) [29], четко оговаривая условия, в которых были получены гендерные различия [61]. Различия показаны в целом ряде вербальных заданий: беглость речи (verbal fluency), правописании (spelling) и грамотности (written expression) [30, 62]. По результатам метаанализа, включавшего работы последних 40 лет, авторы приходят к выводу, что различия есть, они устойчивы, но невелики по значениям [62]. Данные нейрофизиологических исследований показывают, что при различных видах вербальной деятельности функциональная активность мозга у мужчин более латерализована, чем у женщин, для которых более характерно вовлечение в деятельность примерно в равной мере как левого, так и правого полушария мозга [23, 27, 28].
Вопрос о возрасте проявления половых отличий до конца не решен, в связи с недостатком лонгитюдных и онтогенетических исследований. В нашем исследовании, охватывающем возрастной диапазон от 8 до 30 лет, показано, что половые отличия изменения уровня ПСБП начинают проявляться при восприятии текста на слух с 12 лет, а при чтении текста – с 15 лет (рис. 2). Таким образом, учитывая, что при деятельности половые отличия проявляются более явно, чем в состоянии спокойного бодрствования, можно предположить, что на возраст начала проявления половых отличий оказывает влияние вид когнитивной деятельности и степень сформированности навыка. Таким образом, половые отличия выявляются, начиная с подросткового возраста, а сам подростковый возраст является определяющим для формирования половых отличий.
У испытуемых мужского и женского пола выявлена различная возрастная динамика изменения значения показателей степени тесноты связей височных отведений ЭЭГ с совокупностью остальных в зависимости от типа задания. Так, снижение пространственно-синхронных процессов (увеличение локальной активности) височных областей левого полушария (Т1, Т3, Т5) при переходе от подросткового к взрослому возрасту выявлено у испытуемых женского пола при восприятии текстов на слух, а у испытуемых мужского пола – при чтении (рис. 3). В то же время в височных зонах правого полушария (T2, T4) и нижнелобной области (F8) справа при восприятии на слух у испытуемых мужского пола выявлено резкое изменение показателей V(i) от детского к подростковому возрасту (12–14 лет), а затем от подросткового (15–17 лет) к взрослому (рис. 3). Ранее мы получили половые различия ПСБП височных областей левого полушария мозга при чтении текстов у подростков [47, 63]. Это согласуется с данными как о структурных [64], так и функциональных [23, 65] половых различиях в связях между отделами мозга: у мужчин связи более локальные, обособленные [66], больше выражены внутриполушарные взаимодействия, а у женщин – межполушарные [64, 66]. Кроме того, показано, что половое созревание оказывает влияние на миелинизацию [67], функциональная анизотропия (FA) проводящих путей superior frontal gyrus, precentral gyrus и anterior corona radiata выше у мальчиков подростков [68]. Можно предположить, что эти структурные половые особенности отражаются в пространственной организации мозга при вербальной деятельности.
ВЫВОДЫ
1. Выявлено влияние возраста, пола и вида вербальной деятельности на структуру биопотенциального поля мозга у испытуемых от 8 до 30 лет. По мере возрастного развития происходит постепенное увеличение пространственной синхронизации биопотенциалов мозга ПСБП, наблюдаемое как в состоянии спокойного бодрствования с закрытыми и открытыми глазами, так и при двух видах вербальной деятельности: при восприятии текста на слух и при чтении.
2. Уровень пространственной синхронизации биоэлектрической активности мозга во всех возрастных группах (8–11, 12–14, 15–17 лет, взрослые) ниже при выполнении вербальных заданий, чем в фоновом состоянии. Выявлена специфика перестройки межрегиональных взаимодействий биопотенциалов мозга для каждого вида вербальной деятельности по сравнению с соответствующим фоновым состоянием.
3. Половые отличия изменения уровней ПСБП проявляются в большей степени при вербальной деятельности начиная с подросткового возраста: при восприятии текста на слух – с 12 лет, при чтении текста – с 15 лет. Траектория развития функциональной связности мозга, оцениваемая по показателям ПСБП мозга, у мужчин и женщин отличается. У женщин происходит постепенное увеличение уровня ПСБП от детского к подростковому возрасту 15–17 лет, с небольшим последующим его снижением у взрослых. У мужчин возрастная динамика изменения уровня ПСБП различается для состояний с открытыми и закрытыми глазами: в состояниях с закрытыми глазами уровень ПСБП мозга постепенно увеличивается; в состояниях с открытыми глазами – нарастает от детского к подростковому возрасту, не изменяется на протяжении подросткового возраста, а затем опять увеличивается у взрослых.
4. Выявлены топические особенности по показателям тесноты связей долей зон головного мозга у мужчин и женщин при выполнении вербальных заданий: начиная с подросткового возраста вклад височных областей левого полушария выше у испытуемых женского пола, а нижнелобной и передневосочной области правого полушария – у мужского.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Комиссией по этике биомедицинских исследований Института эволюционной физиологии и биохимии им. И.М. Сеченова РАН (Санкт-Петербург).
Информированное согласие. Родители (законные представители) несовершеннолетних участников исследования представили добровольное письменное информированное согласие, подписанное ими после разъяснения им потенциальных рисков и преимуществ, а также характера предстоящего исследования. Все испытуемые участвовали в исследованиях добровольно.
Финансирование работы. Часть исследований в группе подростков 12–17 лет выполнена в рамках РФФИ (грант № 18-313-00169), группы детей 4–11 лет и взрослых исследованы в рамках государственного задания № 075-00776-19-02.
Благодарности. Авторский коллектив выражает благодарность к. б. н. В.П. Рожкову за методическую поддержку.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Фарбер Д.А., Мачинская Р.И., Курганский А., Петренко Н.Е. Функциональная организация коры больших полушарий при подготовке к опознанию неполных изображений у детей 7–8 лет и взрослых // Физиология человека. 2014. Т. 40. № 5. С. 5. Farber D.A., Machinskaya R.I., Kurgansky A.V., Petrenko N.E. Functional organization of the brain in the period of preparation for recognizing fragmented images in seven- to eight-year-old children and adults // Human Physiology. 2014. V. 40. № 5. P. 475.
Семенова О.А., Мачинская Р.И., Ломакин Д.И. Влияние функционального состояния регуляторных систем мозга на эффективность программирования, избирательной регуляции и контроля когнитивной деятельности у детей. Сообщение I. Нейропсихологический и электроэнцефалографический анализ возрастных преобразований регуляторных функций мозга в период от 9 до 12 лет // Физиология человека. 2015. Т. 41. № 4. С. 5. Semenova O.A., Machinskaya R.I., Lomakin D.I. The influence of the functional state of brain regulatory systems on the programming, selective regulation and control of cognitive activity in children: I. Neuropsychological and EEG analysis of age-related changes in brain regulatory functions in children aged 9–12 years // Human Physiology. 2015. V. 41. № 4. P. 345.
Соколова Л.В. ЭЭГ-корреляты направленного внимания у школьников с разной успешностью в чтении // Физиология человека. 2003. Т. 29. № 3. С. 136. Sokolova L.V. EEG Correlates of Selective Attention in Schoolchildren with Different Reading Progress // Human Physiology. 2003. V. 29. № 3. P. 384.
Grigorenko E.L., Kornev A.N., Rakhlin N. et al. Reading-related skills, reading achievement, and inattention: a correlational study // J. Cogn. Education Psychol. 2011. V. 10. № 2. P. 140.
Корнев А.Н., Столярова Э.И., Гальперина Е.И., Гийемар Д.М. Формирование сенсомоторных механизмов продукции слога на начальном этапе усвоения чтения // Педиатр. 2014. Т. 5. № 4. С. 85.
Григорьев А.С., Ляксо Е.Е. Слуховое восприятие слов детей 5–8 лет // Сенсорные системы. 2014. Т. 28. № 3. С. 28.
Berl M.M., Duke E.S., Mayo J. et al. Functional anatomy of listening and reading comprehension during development // Brain Lang. 2010. V. 114. № 2. P. 115.
Deniz F., Nunez-Elizalde A.O., Huth A.G. et al. The Representation of Semantic Information Across Human Cerebral Cortex During Listening Versus Reading Is Invariant to Stimulus Modality // J. Neurosci. 2019. V. 39. № 39. P. 7722.
Sowell E.R., Thompson P.M., Toga A.W. Mapping changes in the human cortex throughout the span of life // Neuroscientist. 2004. V. 10. № 4. P. 372.
Tiemeier H., Lenroot R.K., Greenstein D.K. et al. Cerebellum development during childhood and adolescence: a longitudinal morphometric MRI study // Neuroimage. 2010. V. 49. № 1. P. 63.
Giedd J.N., Raznahan A., Alexander-Bloch A. et al. Child Psychiatry Branch of the National Institute of Mental Health Longitudinal Structural Magnetic Resonance Imaging Study of Human Brain Development // Neuropsychopharmacol. 2015. V. 40. № 3. P. 43.
Vijayakumar N., Op de Macks Z., Shirtcliff E.A., Pfeifer J.H. Puberty and the human brain: Insights into adolescent development // Neurosci Biobehav Rev. 2018. V. 92. P. 417.
Mizuguchi N., Maudrich T., Kenville R. et al. Structural connectivity prior to whole-body sensorimotor skill learning associates with changes in resting state functional connectivity // Neuroimage. 2019. V. 197. P. 191.
Aboud K.S., Bailey S.K., Petrill S.A. et al. Comprehending text versus reading words in young readers with varying reading ability: distinct patterns of functional connectivity from common processing hubs // Dev. Sci. 2016. V. 19. № 4. P. 632.
Stevens M.C. The contributions of resting state and taskbased functional connectivity studies to our understanding of adolescent brain network maturation // Neurosci Biobehav Rev. 2016. V. 70. P. 13.
O'Neill G.C., Tewarie P., Vidaurre D. et al. Dynamics of large-scale electrophysiological networks: A technical review // Neuroimage. 2018. V. 180. Pt. B. P. 559.
Wang S.H., Lobier M., Siebenhühner F. et al. Hyperedge bundling: A practical solution to spurious interactions in MEG/EEG source connectivity analyses // Neuroimage. 2018. V. 173. P. 610.
Schaller F., Weiss S., Müller H.M. EEG beta-power changes reflect motor involvement in abstract action language processing // Brain Lang. 2017. V. 168. P. 95.
Цицерошин М.Н., Шеповальников А.Н. Становление интегративной функции мозга. СПб.: Наука, 2009. 209 с.
Мачинская Р.И., Курганский А.В. Фронтальные билатерально-синхронные тета-волны и когерентность фоновой ЭЭГ у детей 7–8 и 9–10 лет с трудностями обучения // Физиология человека. 2013. Т. 39. № 1. С. 71.
Lenroot R.K., Giedd J.N. Sex differences in the adolescent brain // Brain Cogn. 2010. V. 72. № 1. P. 46.
Сороко С.И., Бекшаев С.С., Рожков В.П. ЭЭГ корреляты генофенотипических особенностей возрастного развития мозга у детей аборигенного и пришлого населения северо-востока России // Российский физиологический журн. им. И.М. Сеченова. 2012. Т. 98. № 1. С. 3.
Панасевич Е.А., Цицерошин М.Н. Отражение в топологических особенностях пространственной организации межкортикальных взаимодействий способности к успешному выполнению детьми 5–6 лет различных видов когнитивной деятельности (гендерные различия) // Физиология человека. 2015. Т. 41. № 5. С. 39. Panasevich E.A., Tsitseroshin M.N. The ability to successfully perform different kinds of cognitive activity is reflected in the topological features of intercortical interactions: sex-related differences between boys and girls aged five to six years // Human Physiology. 2015. V. 41. № 5. P. 487.
Guillemard (Tsaparina) D.M., Tsitseroshin M.N., Shepovalnikov A.N. et al. Ontogenetic development of Neurophysiological Mechanisms Underlying Language Processing / Licensee InTech. Evolutionary Physiology and Biochemistry – Advances and Perspectives. City. 2018. P. 75.
Etchell A., Adhikari A., Weinberg L.S. et al. A systematic literature review of sex differences in childhood language and brain development // Neuropsychologia. 2018. V. 114. P. 19.
Benninger C., Matthis P., Scheffner D. EEG development of healthy boys and girls. Results of a longitudinal study // EEG Clin. Neurophysiol. 1984. V. 57. № 1. P. 1.
Горбачевская Н.Л., Якупова Л.П., Кожушко Л.Ф. и др. Нейробиологические причины школьной дезадаптации // Физиология человека. 1991. Т. 17. № 5. С. 72.
Разумникова О.М. Мышление и функциональная асимметрия мозга. Новосибирск: Изд-во СО РАМН, 2004. 272 с.
Hyde J.S. The gender similarities hypothesis // Am. Psychol. 2005. V. 60. № 6. P. 581.
Reynolds M.R., Scheiber C., Hajovsky D.B. et al. Gender Differences in Academic Achievement: Is Writing an Exception to the Gender Similarities Hypothesis? // J. Genet. Psychol. 2015. V. 176. № 3–4. P. 211.
Burman D.D., Minas T., Bolger D.J., Booth J.R. Age, sex, and verbal abilities affect location of linguistic connectivity in ventral visual pathway // Brain Lang. 2013. V. 124. № 2. P. 184.
Dennis E.L., Thompson P.M. Reprint of: Mapping connectivity in the developing brain // Int. J. Dev. Neurosci. 2014. V. 32. P. 41.
Goddings A.L., Mills K.L., Clasen L.S. et al. The influence of puberty on subcortical brain development // Neuroimage. 2014. V. 88. P. 242.
Развитие мозга и формирование познавательной деятельности ребенка / Под ред. Фарбер Д.А., Безруких М.М. М.: Изд-во Моск. психол.-соц. ин-та; Воронеж: МОДЭК, 2009. 432 с.
Dennison M., Whittle S., Yücel M. et al. Mapping subcortical brain maturation during adolescence: evidence of hemisphere- and sex-specific longitudinal changes // Dev. Sci. 2013. V. 16. № 5. P. 772.
Kansaku K., Kitazawa S. Imaging studies on sex differences in the lateralization of language // Neuroscience Research. 2001. V. 41. № 4. P. 333.
Dehaene-Lambertz G., Hertz-Pannier L., Dubois J. Nature and nurture in language acquisition: anatomical and functional brain-imaging studies in infants // Trends Neurosci. 2006. V. 29. № 7. P. 367.
Telkemeyer S., Rossi S., Koch S.P. et al. Sensitivity of newborn auditory cortex to the temporal structure of sounds // J. Neurosci. 2009. V. 29. № 47. P. 14726.
Indefrey P., Hellwig F., Herzog H. et al. Neural responses to the production and comprehension of syntax in identical utterances // Brain Lang. 2004. V. 89. № 2. P. 312.
Richlan F. The Functional Neuroanatomy of Letter-Speech Sound Integration and Its Relation to Brain Abnormalities in Developmental Dyslexia // Front. Hum. Neurosci. 2019. V. 13. P. 21.
Pugh K.R., Mencl W.E., Jenner A.R. et al. Functional neuroimaging studies of reading and reading disability (developmental dyslexia) // Ment. Retard. Dev. Disabil. Res. Rev. 2000. V. 6. № 3. P. 207.
Cohen L., Lehéricy S., Chochon F. et al. Language-specific tuning of visual cortex? Functional properties of the Visual Word Form Area // Brain. 2002. V. 125. № 5. P. 1054.
Annett M. The binomial distribution of right, mixed and left handedness // Q. J. Exp. Psychol. 1967. V. 19. № 4. P. 327.
Барвинок А.И., Рожков В.П. Особенности межцентральной координации корковых электрических процессов при умственной деятельности // Физиология человека. 1992. Т. 18. № 3. С. 5.
Гантмахер Ф.Р. Теория матриц. М.: Наука, 1967. 576 с.
Кручинина О.В., Гальперина Е.И., Шеповальников А.Н. Особенности пространственной организации колебаний биопотенциалов мозга у подростков // Физиология человека. 2014. Т. 40. № 5. С. 14. Kruchinina O.V., Galperina E.I., Shepovalnikov A.N. Characteristics of the spatial organization of oscillations of brain bioelectric potentials in adolescents // Human Physiology. 2014. V. 40. № 5. P. 483.
Кручинина О.В., Гальперина Е.И. Пространственно-временнáя организация ЭЭГ у подростков 12–13 и 15–17 лет при чтении текстов // Новые исследования. 2018. № 3. С. 5.
Павлова Л.П., Романенко А.Ф. Системный подход к психофизиологическому исследованию мозга человека. Л.: Наука, 1988. 213 с.
Дубровинская Н.В. Развитие речи и организация вербальной деятельности / Развитие мозга и формирование познавательной деятельности ребенка // Под ред. Фарбер Д.А., Безруких М.М. М.: Изд-во Моск. психолого-социального ин-та; Воронеж: Издательство НПО “МОДЭК”, 2009. С. 327.
Цицерошин М.Н., Гальперина Е.И. Коррелятивное формирование функций как один из механизмов функциональной эволюции (на примере становления в онтогенезе ребенка центрального обеспечения стереогноза и функции речи) // Журн. эволюционной биохимии и физиологии. 2012. Т. 48. № 3. С. 296. Tsitseroshin M.N., Galperina E.I. Correlative formation of functions as one of mechanisms of functional evolution (by example of development in childs ontogenesis of central maintenance of stereognosis and speech function) // J. Evol. Biochem. Phys. 2012. V. 48. № 3. P. 355.
Michael E.B., Keller T.A., Carpenter P.A., Just M.A. fMRI investigation of sentence comprehension by eye and by ear: modality fingerprints on cognitive processes // Hum. Brain Mapp. 2001. V. 13. № 4. P. 239.
Шеповальников А.Н., Цицерошин М.Н., Левинченко Н.В. “ Возрастная минимизация” областей мозга, участвующих в системном обеспечении психических функций: аргументы за и против // Физиология человека. 1991. Т. 17. № 5. С. 28.
Thomason M.E., Chang C.E., Glover G.H. et al. Default-mode function and task-induced deactivation have overlapping brain substrates in children // NeuroImage. 2008. V. 41. № 4. P. 1493.
Fair D.A., Cohen A.L., Power J.D. et al. Functional brain networks develop from a “local to distributed” organization // PLoS Comput Biol. 2009. V. 5. № 5. P. e1000381.
Szostakiwskyj J.M.H., Willatt S.E., Cortese F. et al. The modulation of EEG variability between internally- and externally-driven cognitive states varies with maturation and task performance // PLoS ONE. 2017. V. 12. № 7. P. e0181894.
Norton E.S., Beach S.D., Gabrieli J.D. Neurobiology of dyslexia // Curr. Opin. Neurobiol. 2015. V. 30. P. 73.
Wilson S.M., Bautista A., McCarron A. Convergence of spoken and written language processing in the superior temporal sulcus // Neuroimage. 2018. V. 171. P. 62.
Bookheimer S. Functional MRI of language: new approaches to understanding the cortical organization of semantic processing // Annu. Rev. Neurosci. 2002. V. 25. P. 151.
Coltheart M. Modeling Reading: The Dual-Route Approach / Eds. Snowling M.J., Hulme C. The science of reading: A handbook. Oxford: Blackwell, 2005. P. 6.
Spironelli C., Penolazzi B., Angrilli A. Gender Differences in Reading in School-Aged Children: An Early ERP Study // Dev. Neuropsychol. 2010. V. 35. № 4. P. 357.
Hyde J.S. Sex and cognition: gender and cognitive functions // Curr. Opin. Neurobiol. 2016. V. 38. P. 53.
Hirnstein M., Hugdahl K., Hausmann M. Cognitive sex differences and hemispheric asymmetry: A critical review of 40 years of research // Laterality. 2019. V. 24. № 2. P. 204.
Гальперина Е.И., Кручинина О.В., Рожков В.П. Пространственная синхронизация биопотенциалов мозга отличается у юношей и девушек 12–13 лет при чтении повествовательных текстов // Физиология человека. 2018. Т. 44. № 2. С. 31. Galperina E.I., Kruchinina O.V., Rozhkov V.P. Spatial synchronization of brain bioelectric potentials differs in boys and girls aged 12–13 years reading narrative texts // Human Physiology. 2018. V. 44. № 2. P. 143.
Ingalhalikar M., Smith A., Parker D. et al. Sex differences in the structural connectome of the human brain // PNAS. 2014. V. 111. № 2. P. 823.
Marosi E., Harmony T., Becker J. et al. Sex differences in EEG coherence in normal children // Int. J. Neurosci. 1993. V. 72. № 1–2. P. 115.
Tyan Y.S., Liao J.R., Shen C.Y. et al. Gender differences in the structural connectome of the teenage brain revealed by generalized q-sampling MRI // Neuroimage Clin. 2017. V. 15. P. 376.
Perrin J.S., Herve P.-Y., Leonard G. et al. Growth of white matter in the adolescent brain: Role of testosterone and androgen receptor // J. Neurosci. 2008. V. 28. № 38. P. 9519.
Herting M.M., Kim R., Uban K.A. et al. Longitudinal changes in pubertal maturation and white matter microstructure // Psychoneuroendocrinology. 2017. V. 81. P. 70.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека