

Физиология человека, 2020, T. 46, № 4, стр. 5-21
Влияние положения траектории движущихся звуковых стимулов на латерализацию вызванных ответов
Е. А. Петропавловская 1, *, Л. А. Канахина 2, Л. Б. Шестопалова 1
1 ФГБУН Институт физиологии имени И.П. Павлова РАН
Санкт-Петербург, Россия
2 Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
* E-mail: petropavlovskaiae@infran.ru
Поступила в редакцию 28.03.2019
После доработки 06.06.2019
Принята к публикации 10.11.2019
Аннотация
Обработка пространственной акустической информации в слуховой системе человека неразрывно связана с взаимодействием левого и правого полушарий мозга. В данной работе исследовали влияние положения и направления движения звуковых стимулов на амплитуду и латентность компонентов N1, P2, MMN и P3a слуховых вызванных потенциалов в левом и правом полушариях головного мозга. Полученные результаты позволяют охарактеризовать функциональную асимметрию комплекса N1P2 и разностных потенциалов как неустойчивое контралатеральное доминирование. Девианты, движущиеся от латеральных позиций, вызывали более поздние компоненты P3a, чем движущиеся от средней линии. Это согласуется с концепцией более точной локализации звука во фронтальной части акустического пространства. Направление движения влияло на латентности волн MMN и Р3а: центробежные девианты вызывали более ранние реакции MMN и Р3а, чем центростремительные. Это может быть связано с более сильной ориентировочной реакцией, которую вызывают стимулы, удаляющиеся от центра к периферии, по сравнению с движущимися в обратном направлении.
Билатеральная симметрия является одним из основных принципов строения центральной нервной системы (ЦНС) человека. Органы чувств, проводящие пути и центральные структуры являются парными, зеркально симметричными. При этом функциональной организации ЦНС в значительной степени свойственно контралатеральное доминирование. Это значит, что каждое полушарие связано главным образом с моторными функциями противоположной стороны тела и с обработкой сенсорных сигналов, поступающих из противоположной половины окружающего пространства. Однако контралатеральное доминирование в разных сенсорных системах проявляется в разной степени, так как другим основным принципом организации ЦНС является параллельная распределенная обработка информации, что подразумевает многоканальность проведения информации и многоуровневость ее обработки. Информация от рецепторов до коры больших полушарий передается по множеству относительно самостоятельных каналов, а горизонтальные связи обеспечивают взаимодействие между симметричными структурами мозга на разных уровнях сенсорных систем. Такое строение обеспечивает надежность передачи информации, позволяет осуществлять одновременную обработку разных признаков сигнала на разных уровнях и создает условия для тонкого регулирования процессов восприятия [1, 2]. Параллельная распределенная обработка информации также является предпосылкой для появления функциональной межполушарной асимметрии, когда правое и левое полушария в разной степени вносят вклад в анализ сложных признаков сенсорных сигналов или реализацию других функций. Наиболее известным (но далеко не единственным) проявлением функциональной межполушарной асимметрии является доминирование левого полушария при обработке семантической информации. Определение положения источника звука в пространстве и анализ динамически меняющейся информации при движении источника звука также являются сложными процессами, при реализации которых неоднократно была описана асимметрия в активности правого и левого полушарий.
Наиболее информативными признаками, по которым слуховая система определяет положение источника звука, являются различия по времени и по интенсивности между звуковыми стимулами, поступающими на правое и левое ухо. Таким образом, при определении положения источника звука используется информация, получаемая от обоих ушей, и при этом уже на начальных этапах обработки пространственных характеристик входящей сенсорной информации задействованы оба полушария.
В течение долгих лет рассматривались две основные гипотезы относительно обработки бинауральных признаков слуховой системой человека: преимущественная обработка в полушарии, контралатеральном стороне звуковой стимуляции (“контралатеральное доминирование”), и преобладающая роль правой слуховой коры в анализе пространственных признаков звука (“правостороннее доминирование”). Гипотеза о контралатеральном доминировании полушарий мозга при обработке пространственных признаков звуковых стимулов основывается преимущественно на данных о нейронной активности у животных [3, 4], а также на клинических данных [5, 6]. Ряд исследований, проведенных с помощью методов фМРТ, ЭЭГ и МЭГ, показали более сильные или ранние ответы в полушарии, контралатеральном стороне, с которой предъявлялись звуковые стимулы (неподвижные звуки [7–9], движущиеся звуки [10–12]). В то же время существуют данные, свидетельствующие о доминировании правого полушария при анализе пространственной слуховой информации, как основанные на клинических исследованиях [6, 13], так и полученные при регистрации фМРТ или МЭГ-ответов на неподвижные или движущиеся звуковые стимулы (неподвижные звуки [8, 14–16], движущиеся звуки [17, 18]).
Однако к настоящему времени накоплено множество экспериментальных свидетельств того, что данные гипотезы не являются полностью взаимоисключающими. Было установлено, что реакции левого полушария активируются преимущественно правосторонним движением стимула, тогда как правое полушарие одинаково реагирует на движение звуковых стимулов с обеих сторон. Иными словами, движение вправо активирует оба полушария, а движение влево – в основном правую кору [10, 19–21]. Это явление получило название феномена “левостороннего игнорирования” (в англоязычной литературе – neglect model), под которым подразумевают преимущественную обработку контралатеральной пространственной информации в левом полушарии и обработку информации по всему акустическому пространству в правом полушарии мозга [22–26].
Базовые аспекты асимметрии слуховой активности мозга могут быть связаны с первичной (“автоматической”) обработкой слуховой информации на уровне, предшествующем вниманию (в англоязычной литературе – preattentive, preconscious), показателем которой является негативность рассогласования (mismatchnegativity, MMN). В общем виде MMN представляет собой негативную волну в составе вызванного потенциала (ВП), возникающую в ответ на изменение параметров стимуляции [27]. Имеются определенные свидетельства того, что процесс автоматического распознавания различий вызывает более выраженную полушарную специализацию соответствующих нейронных структур, чем независимое предъявление стимулов вне процедуры регистрации MMN [28]. Нарушение закономерности в последовательности звуковых стимулов также может быть обнаружено и на сознательном уровне. Предъявление девианта может приводить к переключению внимания, что сопровождается генерацией компонента P3a в составе ВП [29]. Поэтому негативность рассогласования и компонент P3a интенсивно используются наряду с ВП при изучении функциональной роли левого и правого полушарий в пространственном слуховом анализе.
Основной объем данных относительно межполушарной асимметрии MMN, вызванной пространственными различиями, был получен либо в условиях предъявления неподвижных стандартов и девиантов различной латерализации [24, 25, 28, 30–33], либо для неподвижных стандартов, расположенных в области средней линии головы, и движущихся от них девиантов [34–36], либо при движении стандартов и девиантов от центра (средней линии головы) влево или вправо [37, 38]. Большинство авторов приходят к выводу о преимущественно контралатеральной первичной обработке пространственных звуковых различий. В то же время в ряде работ отмечены проявления феномена “левостороннего игнорирования” [24, 25, 38].
Однако данные литературы свидетельствуют о том, что параметры ВП и MMN (особенно ее латентность) проявляют чувствительность к положению начала траектории стимулов (MMN [31], ВП [12, 20, 39]). Смещение стимулов по направлению к средней линии или от нее, т.е. в центростремительном или центробежном направлении, может отражаться на степени контралатеральной асимметрии реакции. Так, в работе N. Richter et al. [31] показано, что степень межполушарной асимметрии MMN зависит от направления смещения девиантных стимулов: контралатеральное преобладание MMN значительно более выражено при центробежном смещении стимулов (от неподвижных латерализованных стандартов к периферии), чем при центростремительном смещении (к средней линии головы).
Данное исследование было посвящено влиянию положения звукового стимула (центральное или латеральное, слева или справа) и направления движения (влево или вправо) на компоненты N1, P2, MMN и P3a, зарегистрированные в левом и правом полушариях, а также сравнению параметров центробежной и центростремительной MMN, вызванной плавным движением звукового стимула. Для этого был применен режим традиционной oddball-стимуляции, включавшей помимо центральных неподвижных стандартов также и латерализованные. Девианты перемещались от стандартов к средней линии либо в обратном направлении. На основании данных литературы были выдвинуты следующие предположения, относящиеся как к компонентам N1 и P 2, так и к разностным потенциалам: 1) реакция на центробежное движение стимула больше по амплитуде, чем на центростремительное движение; 2) контралатеральность реакции нарастает при движении к периферии и падает при движении к центру. Задача проведенного эксперимента состояла в проверке этих предположений.
МЕТОДИКА
В эксперименте участвовали 12 испытуемых (2 мужчин, 10 женщин, все праворукие) в возрасте от 21 до 42 лет. До начала опытов все испытуемые проходили стандартную процедуру аудиометрии на частотах 125, 250, 500, 750, 1000, 1500, 2000, 3000, 4000, 6000 и 8000 Гц. Отклонения индивидуальных пороговых величин от нормального уровня слуха не превышали 10 дБ. Разница в порогах, измеренных для правого и левого уха, не превышала 5 дБ в диапазоне частот 750–1500 Гц и 10 дБ в диапазонах 125–500 и 2000–8000 Гц. Слушателей располагали в комфортном кресле в экранированной звукоизолированной камере. В ходе регистрации ЭЭГ им предлагали читать книгу по своему выбору. Каждый испытуемый проходил экспериментальный цикл полностью.
Стимулы. Звуковыми стимулами служили дихотически предъявленные отрезки белого шума полосой 100–1300 Гц и длительностью 200 мс, синтезированные в цифровом виде. Фронты нарастания и спада длительностью 10 мс были сглажены косинусоидальной функцией. Дихотическое предъявление всех применявшихся звуковых стимулов вызывало у испытуемых ощущение слитных звуковых образов, локализованных внутри головы.
В ходе данного эксперимента использовали два типа пространственных звуковых стимулов – неподвижные и движущиеся. Неподвижные стимулы были локализованы вблизи средней линии головы (в дальнейшем для краткости – по центру), либо приблизительно в середине правой или левой части субъективного акустического пространства. Движущиеся стимулы плавно смещались от неподвижных влево или вправо. Эффект плавного движения стимулов создавали за счет динамических изменений межушной задержки ΔT в сигнале в пределах от 0 до 400 мкс. Ранее, в нашей [40], выполненной с использованием стимулов с аналогичными частотными и временны́ми характеристиками, для неподвижных и плавно движущихся стимулов была получена зависимость воспринимаемого положения стимула от величины ΔT в диапазоне задержек от 0 до 800 мс. Значение ΔT = 200 мкс соответствовало воспринимаемому угловому положению звукового образа 33 град, а ΔT = 400 мкс – положению 57 град относительно средней линии головы.
При записи ЭЭГ в режиме oddball-парадигмы неподвижные стимулы использовали в качестве стандартов, а движущиеся в качестве девиантов. По расположению стандарта стимулы объединяли в 3 типа серий: Центр, Левая и Правая (рис. 1, А).
Рис. 1.
Схематическое изображение звуковых сигналов, расположения электродов для регистрации ЭЭГ, группировки данных для статистического анализа. А – неподвижные и движущиеся звуковые стимулы в сериях Левая, Центр и Правая. I – схематическое изображение положения стандартного стимула и траекторий перемещения девиантов. Пояснения условных названий стимулов даны в тексте. II – изменения межушной задержки (ΔT) в стандартных и девиантных стимулах. По оси абсцисс – длительность стимула, мс. По оси ординат – межушная задержка, мкс. Б – симметричные электродные кластеры. В – схематическое изображение группировки данных по выборкам для проведения статистического анализа, пояснения в тексте.
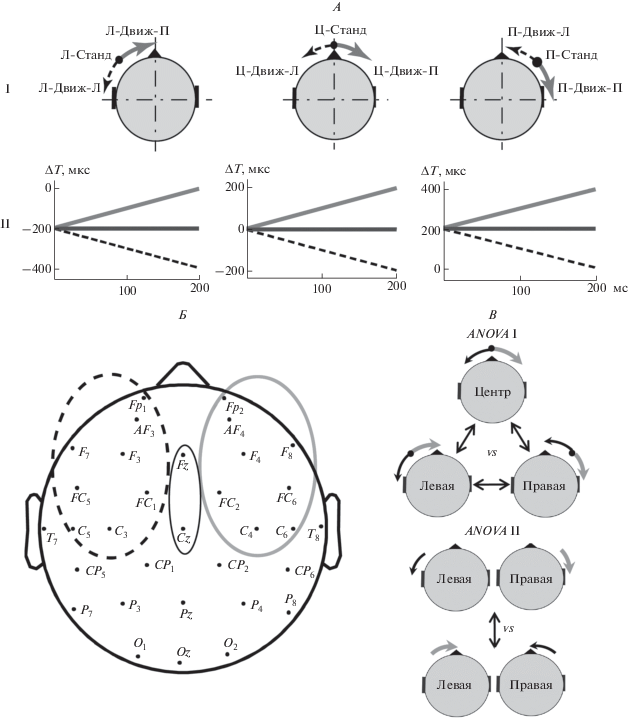
Серия Центр: Стандарт (Ц-Станд) – стимул с постоянным значением межушной задержки ΔT = 0 мкс, вызывающий ощущение неподвижного звукового образа, расположенного по центру головы. Девианты Ц-Движ-Л и Ц-Движ-П моделировали плавное движение вправо и влево от центра за счет линейного изменения ΔT от 0 до ±200 мкс.
Серия Правая: Стандарт (П-Станд) – стимул с постоянным значением ΔT = 200 мкс. Такой стимул испытуемые воспринимали как неподвижный, расположенный в правой половине акустического пространства. Девиант П-Движ-П – плавное движение от стандарта к уху (вправо) за счет линейного нарастания ΔT от 200 до 400 мкс. Девиант П-Движ-Л – плавное движение от стандарта к центру (влево). Величина ΔT линейно уменьшается от 200 мкс до 0.
Серия Левая: Стандарт (Л-Станд) – стимул с постоянным значением ΔT = –200 мкс, которому соответствовал неподвижный звуковой образ в левой половине акустического пространства. Девиант ЛHДвиж-П – плавное движение от стандарта к центру (вправо) за счет линейного уменьшения ΔT от –200 мкс до 0. Девиант Л-Движ-Л – плавное движение от стандарта к уху (влево). Величина ΔT линейно нарастала от –200 до –400 мкс.
Синтезированные сигналы преобразовывали в аналоговую форму при помощи многоканальной аудиоплаты Gina24 (EchoAudio, США). Для дихотического предъявления сигналов непосредственно к барабанной перепонке применяли звукоизлучатели ER-2 (Etymotic Research Inc., США) с амплитудно-частотными характеристиками, имеющими в диапазоне 0.1–10 кГц неравномерность ±3 дБ. Наконечники звукоизлучателей обеспечивали дополнительное подавление внешних шумов на 30 дБ. Интенсивность сигналов устанавливали на уровне 50 дБ над порогом слышимости на каждом ухе испытуемого.
Процедура эксперимента. В ходе записи ЭЭГ слушателям предъявляли по 8 серий каждого вида (Левая, Правая и Центр), причем порядок серий был сбалансирован псевдослучайным образом для каждого испытуемого в отдельности. На каждом испытуемом записывали по 24 серии ЭЭГ (3 раза по 8 серий в разные дни). Каждая серия продолжалась около 8 мин, состояла из 500 стимулов с межстимульным интервалом от начала стимула до начала следующего l с, и содержала 420 стандартов и по 40 девиантов, движущихся влево и вправо от стандарта (итого 80 девиантов в одной серии). Вероятность предъявления составляла 84% для стандарта и по 8% для каждого типа девиантов.
Регистрация и обработка результатов ЭЭГ. Регистрацию ЭЭГ проводили при помощи хлорсеребряных электродов, в 32 точках по международной системе 10–20, с заменой точек отведения PO3/PO4 на C5/C6. Дополнительные электроды располагали на мочках ушей и на носу. Горизонтальную и вертикальную электроокулограммы записывали при помощи электродов, установленных над правым глазом и под левым глазом. Регистрацию ЭЭГ осуществляли цифровым энцефалографом (ActiveTwo, BIOSEMI, Голландия), при помощи специально разработанного программного обеспечения фильтровали и записывали на диск компьютера. В процессе эксперимента ЭЭГ оцифровывали в полосе 0–102 Гц с частотой дискретизации 2048 Гц, а затем подвергали децимации до частоты 512 Гц. По окончании эксперимента из ЭЭГ выделяли эпохи анализа длиной 1000 мс, включая предстимульный интервал 300 мс, принимавшийся за базовую линию. Эпохи, содержащие артефакты амплитудой более 500 мкВ, удаляли из анализа на основе визуального просмотра. Количество удаленных таким образом эпох не превышало 2%. Удаление оставшихся артефактов производили на основе разложения по независимым компонентам (independent component analysis, ICA). Данное преобразование позволяет разложить сигнал ЭЭГ на составляющие компоненты, выделить артефакты связанные с движениями глаз, морганиями и разрывом контакта, а затем восстановить ЭЭГ без артефактов [41]. В соответствии с методическими рекомендациями для разложения по независимым компонентам, проводили вычисление потенциала на каждом канале относительно усредненного потенциала всех электродов, и фильтровали в диапазоне 0.5–45 Гц. После удаления артефактов и обратного преобразования ЭЭГ фильтровали в полосе 2–30 Гц, в качестве референта принимали усредненный потенциал электродов на мочках ушей. Затем проводили коррекцию ЭЭГ относительно базовой линии по участку 300 мс перед началом стимула и сортировали эпохи анализа по видам сигналов.
Анализ данных. Для анализа межполушарной асимметрии вызванной активности выделяли симметричные электродные кластеры в левом и в правом полушарии: FC1, Fp1, AF3, F3, C3, FC5, C5, F7 в левом и FC2, Fp2, AF4, F4, C4, FC6, C6, F8 – в правом, а также медиальный кластер (Fz, Cz) (рис. 1, Б).
Слуховые ВП вычисляли для каждого кластера путем усреднения данных ЭЭГ по включенным в кластер каналам, для каждого типа стимула и для каждого испытуемого отдельно. Разностную волну получали вычитанием ответа на стандарт из ответа на девиант. Достоверность отличия амплитуд разностных потенциалов (MMN и P3a) от нуля проверяли методом скользящего среднего. Амплитуды индивидуальных разностных потенциалов усредняли во временнóм окне шириной 50 мс, сдвигавшемся последовательно на протяжении всей эпохи анализа с шагом 2 мс. Отличие каждой полученной таким образом выборки от нуля проверяли при помощи двустороннего t-теста Стьюдента с уровнем значимости α = 0.1.
После получения для каждого кластера индивидуальных слуховых ВП на стандартные и девиантные стимулы, в них оценивали амплитуды компонентов N1, P2, MMN и P3a. Амплитуды всех компонентов измеряли как средние величины потенциала индивидуально для каждого испытуемого во временнóм окне ±25 мс по обе стороны от соответствующего пика гранд-усредненной кривой в медиальном кластере. Индивидуальные пиковые латентности измеряли в том же временнóм окне относительно пика на гранд-усредненной кривой.
Статистический анализ. Измеренные по индивидуальным данным средние амплитуды и пиковые латентности компонентов N1, P2, MMN и P3a подвергали пилотному трехфакторному дисперсионному анализу с повторными измерениями (repeated measures ANOVA) с факторами Серия (левая, центр, правая), Стимул (влево от стандарта, стандарт, вправо от стандарта), Кластер (левый, медиальный, правый). Пилотный статистический анализ выявил высокую значимость фактора Кластер для большинства исследуемых компонентов. Последующие попарные сравнения показали, что ответы в медиальном кластере имели большую амплитуду и меньшую латентность, чем в двух других кластерах. Поскольку задача исследования состояла в сравнении активности двух полушарий, ответы медиального кластера не включали в основной и дополнительный дисперсионный анализ.
Для того чтобы оценить влияние положения неподвижных звуковых стимулов на асимметрию распределения основных компонентов ВП по поверхности головы, средние амплитуды и пиковые латентности компонентов N1 и P2 в ответах на стандартные стимулы подвергали двухфакторному дисперсионному анализу с повторными измерениями с факторами Стандарт (левый, центральный, правый) и Кластер (левый, правый). Для средних амплитуд и пиковых латентностей компонентов N1, P2, MMN и P3a ответов на движущиеся девиантные стимулы проводили трехфакторный дисперсионный анализ с повторными измерениями с факторами Стандарт (левый, центральный, правый), Девиант (движение влево и вправо от стандарта) и Кластер (левый, правый). Так как девиантные стимулы в каждой серии двигались влево или вправо от стандартных, фактор Стандарт в данном анализе соответствовал положению начала траектории движения, а также сектору субъективного акустического пространства, в котором располагалась траектория движения стимула (рис. 1, В, ANOVA I).
Для ответов на девиантные стимулы, движение которых начиналось от правого и левого стандартов, дополнительно оценивали влияние направления движения относительно средней линии субъективного акустического пространства (к центру или к периферии). Ответы на девиантные стимулы, движущиеся от латеральных стандартов, а также соответствующие разностные потенциалы, перегруппировали в две выборки: центростремительные (Л-Движ-П и П-Движ-Л) и центробежные (Л-Движ-Л и П-Движ-П). Ответы на девианты, движущиеся от центрального стандарта, из этого анализа были исключены, так как в сериях с центральным стандартом центростремительное движение невозможно. Индивидуальные средние амплитуды и пиковые латентности N1, P 2, MMN и P3a подвергали трехфакторному дисперсионному анализу с повторными измерениями с факторами Сторона звучания (левая, правая), Направление (центростремительное, центробежное), Кластер (левый, правый) (рис. 1, В, ANOVA II).
Влияние направления движения (к центру или к периферии) на амплитуду и латентность MMN и P3a также исследовали для реакций медиального кластера при помощи двустороннего парного t-теста. Реакции на лево- и правосторонние стимулы в этом анализе были усреднены.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Все стандартные и девиантные стимулы вызывали отчетливые ВП во всех трех кластерах. Вызванные ответы на стандартные и девиантные стимулы и разностные потенциалы, зарегистрированные в медиальном кластере и усредненные по всем испытуемым, а также топограммы распределения активности по поверхности головы, представлены на рис. 2–4. Различия вызванных ответов на девиантные стимулы и ответов на стандартные стимулы позволили выделить в разностных волнах компоненты MMN с латентностью 200–300 мс и P3a с латентностью 350–400 мс. Достоверность отличия разностных компонентов от нуля проверяли методом скользящего среднего. Соответствующие временны́е интервалы выделены на рис. 2–4 серым цветом.
Рис. 2.
Вызванная активность медиального кластера при предъявлении Левой серии. А – гранд-усредненные ВП на стандартные и девиантные стимулы, а также соответствующие разностные потенциалы. По оси абсцисс – время, мс. По оси ординат – разность потенциалов, мкВ. Тонкие линии – реакции на стандарт, жирные линии – реакции на девиант, пунктирные линии – разностный потенциал. Серым цветом выделены интервалы, в которых различия между ответами на стандарт и девиант были достоверны. Б – топограммы амплитуд компонентов ВП и разностных потенциалов, полученные в моменты времени, указанные рядом с каждой топограммой. Негативность показана черным цветом, позитивность – белым. Типы сигналов см. рис. 1, А.
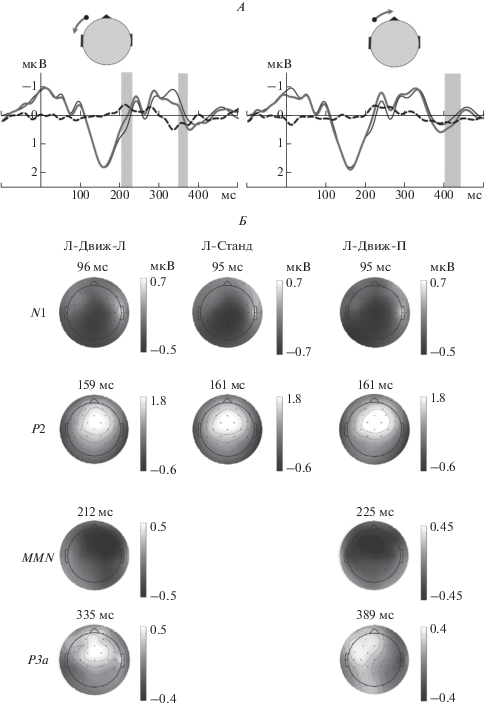
Рис. 3.
Вызванная активность медиального кластера при предъявлении Центральной серии. Обозначения см. рис. 2.

Рис. 4.
Вызванная активность медиального кластера при предъявлении Правой серии. Обозначения см. рис. 2.

Влияние положения звуковых стимулов на средние амплитуды и пиковые латентности компонентов N1 и P2 ВП на неподвижные стандарты оценивали с помощью двухфакторного дисперсионного анализа (Стандарт (левый, центральный, правый) * Кластер (левый, правый)). Для амплитуды N1 была получена высокая значимость фактора Кластер (F(1, 11) = 7.13, p < 0.05) и двухфакторного взаимодействия Стандарт * Кластер (F(1.99, 21.91) = 7.35, p < 0.01). В качестве иллюстрации различий на рис. 5, А, Б, В приведены ВП, полученные в левом и правом кластерах, в ответ на стандартные стимулы из серий Левая, Центр и Правая. Согласно попарным сравнениям, амплитуда N1 была больше в левом кластере, чем в правом, при центральном и правом положениях стандартов (p < 0.05) (рис. 5, А, В). В ответах на стандарты серии Левая достоверных отличий в амплитуде компонента N1 обнаружено не было (p > 0.05) (рис. 5, Б). Для амплитуды компонента P2, а также для латентностей N1 и P2 значимых эффектов и взаимодействий не обнаружено (p > 0.05).
Рис. 5.
ВП в левом и правом кластерах в ответ на неподвижные стандартные (А, Б, В) и движущиеся девиантные (Г, Д) стимулы. А, Б, В – эффект Кластера и взаимодействие Стандарт * Кластер для компонентов N1 и P2, полученных при предъявлении неподвижных стандартных стимулов. Г, Д – эффект Кластера и взаимодействие Сторона * Кластер для компонентов N1 и P2, полученных при предъявлении центробежных и центростремительных девиантов в Левой и Правой сериях. Черные пунктирные линии – ВП левого кластера, серые сплошные линии – ВП правого кластера. По оси абсцисс – время, мс. По оси ординат – разность потенциалов, мкВ. Значимые различия амплитуд показаны в рамках вокруг названия компонента: прямоугольная рамка – результаты ANOVA I по факторам Стандарт (левый, центральный, правый) * Направление (влево, вправо) * Кластер (левый, правый); круглая рамка – результаты ANOVA II по факторам Направление (центростремительное, центробежное) * сторона звучания (левая, правая) * Кластер (левый, правый). Типы стимулов обозначены на схемах головы.

Для компонентов N1 и P2 ВП на движущиеся девиантные стимулы трехфакторный дисперсионный анализ (ANOVA I: Стандарт (левый, центральный, правый) * Девиант (движение влево и вправо от стандарта) * Кластер (левый, правый)) дал сходные результаты. Для амплитуды N1 была получена высокая значимость фактора Кластер (F(1, 11) = 8.25, p < 0.05) при пограничной значимости двухфакторного взаимодействия Стандарт * Кластер (F(1.44, 15.83) = 3.55, p = 0.066). Попарные сравнения показали, что в ответах на девианты амплитуда N1 также была больше в левом кластере, чем в правом, при центральном и правом положениях стандартов (p < 0.01). Направление движения девианта (вправо или влево от стандарта) на амплитуду N1 не влияло. В качестве иллюстрации различий на рис. 5, Г и Д приведены ВП, полученные в левом и правом кластерах в ответ на движущиеся девиантные стимулы. Ответы на стимулы, двигавшиеся влево и вправо от стандарта, усреднены между собой. Достоверные различия в амплитуде, полученные в попарных сравнениях, отмечены стрелками с названием компонента в прямоугольной рамке. Анализ амплитуды компонента P2 не выявил значимых эффектов и взаимодействий, несмотря на существующие достоверные различия при попарных сравнениях, отмеченные на рис. 5, Г. Эти различия проявили себя в дополнительном дисперсионном анализе, результаты которого описаны ниже. Анализ латентностей N1 и P2 значимых эффектов и взаимодействий не выявил (p > 0.05).
Аналогичный дисперсионный анализ не выявил значимых факторов и взаимодействий для амплитуды компонентов MMN и P3a, однако перечисленные факторы оказывали влияние на их латентности. Для латентности компонента MMN анализ показал высокую значимость факторов Стандарт (F (1.75, 19.21) = 32.22, p < 0.001), Девиант (F(1.00, 11.00) = 24.05, p < 0.001) и двухфакторного взаимодействия Стандарт * Девиант (F(1.88, 20.67) = 27.45, p < 0.001). Попарные сравнения показали, что латентность компонента MMN была больше при движении влево от правого стандарта, чем от левого и центрального, а также больше, чем при движении вправо от правого стандарта (p < 0.01) (рис. 6, А). Для латентности компонента P3a анализ показал только высокую значимость фактора Стандарт (F(1.78, 19.62) = 75.84, p < 0.001) при отсутствии других значимых факторов и взаимодействий. При движении девиантов от центрального стандарта латентность P3a была достоверно меньше, чем при движении от левого или правого (p < 0.01) (рис. 6, Б). Этот эффект не зависел от направления движения и проявлялся одинаково в левом и правом кластерах.
Рис. 6.
Эффекты положения стандарта, направления движения и кластера для гранд-усредненных разностных потенциалов. А – латентность MMN при движении от левого, центрального и правого стандарта. 1 – ответ на движение влево, 2 – ответ на движение вправо. Латентности ответов левого и правого кластера усреднены между собой. Б – латентность компонента P3a при движении от левого, центрального и правого стандарта. Латентности ответов на правые и левые девианты, а также ответов левого и правого кластеров усреднены между собой для каждой серии стимулов. Звездочками показаны достоверные различия (*** – p < 0.001). В – эффект направления движения. Слева – разностные потенциалы серии Левая; справа – серии Правая. Ответы левого и правого кластеров усреднены между собой. а – разностный потенциал при центробежном направлении движения девианта, б – при центростремительном. Г – эффект кластера при движении девиантов от латерализованных стандартов. Слева – разностные потенциалы серии Левая; справа – серии Правая. Ответы на центробежные и центростремительные девианты усреднены между собой. Достоверные различия показаны стрелками рядом с названиями компонентов. По оси абсцисс – время от начала действия стимула, мс. По оси ординат – амплитуда реакции, мкВ. в – разностный потенциал в левом кластере электродов, г – в правом кластере.

Влияние центростремительного или центробежного направления движения (от латеральных стандартов к центру или к периферии) на средние амплитуды и пиковые латентности компонентов N1, P2, MMN и P3a оценивали с помощью трехфакторного дисперсионного анализа Направление (центростремительное, центробежное) * Сторона звучания (левая, правая) * Кластер (левый, правый) (ANOVA II). Фактор Направление не оказал значимого влияния ни на амплитуду, ни на латентность компонентов N1 и P2 (p > 0.05). Был обнаружен главный эффект Кластера для амплитуд N1 и P2 (F(1, 11) = 6.26, и F(1, 11) = 5.95, p < < 0.05), который сопровождался значимым взаимодействием Сторона * Кластер (F(1, 11) = 4.31, p = 0.062 для N1 и F(1, 11) = 5.67, p < 0.05 для P2). Согласно попарным сравнениям, при движении с правой стороны амплитуда N1 в левом кластере (контралатеральном) была больше, чем в правом (p < 0.05) (рис. 5, Д). Амплитуда компонента P2 в правом кластере была больше, чем в левом, но эти различия достигали уровня достоверности только при движении с левой стороны (контралатерально) (рис. 5, Г). Других значимых эффектов и взаимодействий не обнаружено (p > 0.05). Достоверные различия в амплитуде, полученные в попарных сравнениях в этом анализе, на рис. 5, Г и Д отмечены стрелками с названием компонента в круглой рамке.
Для амплитуды компонентов MMN и P3a значимых эффектов и взаимодействий также не обнаружено (p > 0.05). Для латентности MMN анализ показал высокую значимость фактора Сторона звучания (F(1, 11) = 40.76, p < 0.01) и двухфакторного взаимодействия Сторона * Направление (F(1, 11) = 30.79, p < 0.01). Попарные сравнения показали, что при движении в правой части слухового пространства латентность центростремительной MMN больше, чем центробежной (рис. 6, В, правая панель). Для латентности P3a также обнаружено значимое двухфакторное взаимодействие Сторона * Направление (F(1, 11) = = 7.54, p < 0.05). Попарные сравнения показали, что латентность компонента P3a также была больше в случае центростремительного движения по сравнению с центробежным, но эти различия были выявлены только при движении в левой части слухового пространства (рис. 6, В, левая панель) и имели пограничный уровень значимости (p = 0.053). Кроме того, имело место значимое двухфакторное взаимодействие Сторона * Кластер (F(1, 11) = 4.94, p < 0.05). Согласно попарным сравнениям, движение с левой стороны вызывало более ранний компонент P3a в правом кластере (контралатеральном), чем в левом (рис. 6, Г).
Влияние направления движения (к центру или к периферии) на амплитуду и латентность MMN и P3a было также исследовано для реакций медиального кластера при помощи двустороннего парного t-теста. Реакции на лево- и правосторонние стимулы в этом анализе были усреднены (рис. 7). Для амплитуд обоих компонентов значимых различий не обнаружено. Латентность центростремительной MMN оказалась больше, чем центробежной, как при движении от центрального стандарта (t = 6.33, p < 0.001), так и при движении от латеральных (t = 5.82, p < 0.001). Латентность P3a была меньше при движении от центрального стандарта, чем от латеральных: t = 10.12, p < 0.001 при сравнении с центростремительным P3a, и t = = 14.92, p < 0.001 при сравнении с центробежным P3a. Аналогичный результат был описан выше для реакций левого и правого полушария (рис. 6, Б, В).
Рис. 7.
Эффекты положения стандарта и направления движения для гранд-усредненных разностных потенциалов медиального кластера. а – разностный потенциал на центробежные стимулы, движущиеся от центрального стандарта; б – разностный потенциал на центробежные стимулы, движущиеся от латеральных стандартов, в – разностный потенциал на центростремительные стимулы, движущиеся от латерализованных стандартов. По оси абсцисс – время от начала действия стимула, мс. По оси ординат – амплитуда реакции, мкВ. Реакции на симметричные стимулы в правой и левой стороне акустического пространства усреднены между собой. Стрелками отмечены компоненты, для которых были получены достоверные различия пиковых латентностей. Типы стимулов обозначены на схемах головы.

ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В данном эксперименте все стандартные стимулы были неподвижными и совпадали с положением начальной точки траектории движения девиантов. Исследовалось влияние положения начала траектории движения стимула (центральное или латеральное, слева или справа) и направления его движения (влево или вправо, к центру или к периферии) на компоненты N1, P2, MMN и P3a, зарегистрированные в левом и правом полушариях.
Как показало исследование, компоненты N1 и P2 проявляли в целом слабую чувствительность к исследуемым факторам. Амплитуда компонента N1 продемонстрировала доминантность левого полушария для неподвижных стандартных стимулов, расположенных в правом полуполе акустического пространства и вблизи средней линии головы, а неподвижные стимулы левого акустического пространства вызывали в левом и правом полушариях мозга реакции одинаковой амплитуды (рис. 5, А, Б, В). Девианты, расположенные справа (независимо от направления движения), вызывали больший компонент N1 в левом полушарии, чем в правом. Девианты, расположенные слева (также независимо от направления) вызывали больший компонент P2 в правом полушарии, чем в левом. Оба результата указывают на контралатеральное преобладание ранних компонентов ВП на движущиеся звуковые стимулы, при том, что центральное или латеральное положение начала траектории и направление движения стимулов на эти компоненты не влияли. Однако, тот факт, что право- и левостороннее расположение движущихся стимулов отражалось на разных компонентах ВП, трудно объяснить на основании имеющихся представлений о природе суммарного ответа. Если рассматривать межполушарную асимметрию с точки зрения ее временнóй динамики, то согласно полученным результатам, контралатеральность реакции на правосторонние стимулы развивалась раньше, чем в случае левосторонних стимулов (на временны́х интервалах волн N1 и P2, соответственно). Если же рассматривать компоненты N1 и P2 как единый комплекс N1P2, который считается более устойчивой характеристикой ВП, чем его составляющие, то амплитуда комплекса N1P2 от пика до пика оказывается меньше для ипсилатеральных ответов, чем для контралатеральных. В правом кластере это различие достигается за счет меньшей амплитуды N1 и P2 в реакциях на правосторонние стимулы, а в левом кластере – за счет уменьшения амплитуды P2 в реакциях на левосторонние стимулы.
В предыдущих исследованиях асимметрии реакций на движение от центра [34, 37] для всех звуковых стимулов было выявлено устойчивое правостороннее доминирование в диапазоне латентностей 110–160 мс после начала сигнала, соответствующем нисходящей ветви компонента N1. Изменение картины асимметрии компонентов N1 и P2 в данном исследовании по сравнению с предыдущими может быть связано с более широким набором положений и траекторий звуковых стимулов, что потребовало более активного взаимодействия между полушариями при анализе пространственной информации в целом.
Положение начала траектории (центральное или латеральное) оказывало наиболее заметный эффект на латентность волны P3a, регистрируемой во всех трех кластерах (рис. 6, Б, рис. 7). Самый ранний компонент P3a формировался при движении от центра. Как известно, этот позитивный компонент разностного потенциала считается индикатором переключения внимания на девиантный стимул. Он может отражать как сенсорные, так и когнитивные аспекты сенсорного события без выраженной модальной специфичности [42, 43]. В настоящее время связь между процессами, формирующими N1 и MMN на первом этапе обработки информации и вызывающими P3a на втором этапе этой обработки, остается до конца не выясненной [44, 45]. Полученные нами результаты также не позволяют установить, почему влияние положения начала траектории слабее проявлялось на этапе формирования MMN. В разностном потенциале, полученном при движении от центра, увеличение амплитуды MMN по сравнению с движением от латеральных стандартов (рис. 7) не достигло уровня достоверности. Единственная имеющаяся в литературе работа, в которой сравниваются разностные потенциалы при разной степени латерализации стандарта, свидетельствует о большей латентности MMN при смещении стандартов к периферии, по сравнению с центральным стандартом [31]. Согласно нашим данным, такой эффект достоверно наблюдался только при движении от правого стандарта к центру (рис. 6, А). В нашем исследовании латентность волны P3a оказалась более чувствительна к положению начала траектории (рис. 6, Б). В целом увеличение амплитуды MMN и формирование более раннего P3a при движении от центра соответствует представлениям о наибольшей чувствительности слуховой системы к смещению источника звука вблизи средней линии головы [46, 47]. Высокая пространственная разрешающая способность позволяет раньше обнаружить небольшое изменение положения звукового образа, что приводит к формированию более раннего и сильного ответа.
Направление движения от латеральных стандартов к центру или к периферии также отражалось в большей степени на времени формирования MMN и P3a, чем на их величине. При движении девиантов в правой части слухового пространства центробежная MMN формировалась раньше, чем центростремительная (рис. 6, Г). Это находится в соответствии с результатами, которые получили N. Richter et al. [31]. Согласно их исследованию MMN на стимулы, двигавшиеся к центру, имели большую латентность, чем, двигавшиеся к периферии [31]. В нашем исследовании при движении слева центробежный компонент P3a также формировался раньше, чем центростремительный. Компонент P3a связывают с непроизвольным переключением внимания на неожиданный стимул и рассматривают как коррелят ориентировочной реакции [48, 49]. Можно предположить, что стимулы, удаляющиеся от центра к периферии, вызывают более раннюю ориентировочную реакцию, чем движущиеся в обратном направлении. Ориентировочная реакция включает в себя движения головы, глаз и ушей в направлении источника раздражения. Это позволяет поместить новый стимул в область максимальной разрешающей способности сенсорных систем, что создает условия для лучшего восприятия нового раздражителя. Если новый стимул появляется в центре зрительного или слухового поля, либо движется к центру, он уже находится в области максимальной разрешающей способности или движется к ней. В этом случае ответная двигательная реакция имеет меньшую амплитуду и большую латентность. В частности, для зрительных стимулов, движущихся в горизонтальной плоскости, было показано, что движение от точки фиксации взгляда вызывало более быстрые реакции, чем движение к точке фиксации, а разница между движением к точке фиксации и движением от нее увеличивалась с эксцентриситетом стимула [50]. Это совпадает с наблюдаемым соотношением латентностей компонента P3a на центробежные и центростремительные стимулы.
Кроме того, при движении с левой стороны компонент P3a возникал раньше с контралатеральной стороны (в правом кластере), чем с ипсилатеральной (рис. 6, Д); при этом на этапе формирования MMN наблюдалась только слабая тенденция к контралатеральности амплитуды разностного потенциала. Таким образом, левосторонние стимулы вызывали более поздние проявления эффекта направления, чем правосторонние (временны́е диапазоны MMN и P3a соответственно), и такие же поздние проявления контралатеральности. В уже упоминавшейся работе N. Richter et al. [31] отмечена зависимость полушарной асимметрии MMN от направления смещения девиантов: при смещении к периферии от центральных или латеральных стандартов наблюдалось контралатеральное доминирование амплитуд MMN, а при смещении от латеральных стандартов к центру асимметрия отсутствовала. Одной из возможных причин несоответствия этих результатов нашим данным могли быть низкие амплитуды полученных разностных потенциалов. Для сравнения, в работе N. Richter et al. [31], амплитуды MMN левого и правого полушарий составляли от 1.5 до 2.8 мкВ в разных условиях. Поскольку амплитуда MMN отражает степень различий между стандартом и девиантом, следующим шагом в исследовании асимметрии ВП и MMN может стать использование стимулов с более длинными траекториями движения, вызывающими высокоамплитудную MMN.
Исследования ВП на центробежные и центростремительные изменения ∆T показали большую величину ответа при смещении к периферии [20, 39]. Результаты данного эксперимента не подтвердили эти выводы и не выявили связи между полушарной асимметрией ВП или разностных потенциалов и направлением смещения к центру или к периферии. Вероятно, различия связаны с тем, что в упомянутых работах исследовались ответы на резкое перемещение звукового стимула как на отдельное событие. В данном исследовании величина межушных различий стимуляции изменялась плавно, а начало изменений совпадало с включением звукового стимула.
ЗАКЛЮЧЕНИЕ
Подводя краткий итог проведенного исследования, можно охарактеризовать функциональную асимметрию комплекса N1P2 и разностных потенциалов как неустойчивое контралатеральное доминирование. Положение траектории движущихся девиантов отражалось не на величине разностных потенциалов, а на времени их формирования. Направление движения было связано с латентностями волн MMN и Р3а: центробежные девианты вызывали более ранние реакции MMN и Р3а, чем центростремительные. Это явление можно рассматривать как электрофизиологический коррелят ориентировочного ответа на звуковые объекты, удаляющиеся от средней линии. Девианты, движущиеся от латеральных позиций, вызывали более поздние компоненты P3a, чем те, которые двигались от средней линии. Это согласуется с концепцией более точной локализации звука во фронтальной части акустического пространства.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Этическим комитетом Санкт-Петербургского государственного университета (Санкт-Петербург).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа выполнена при финансовой поддержке ПФНИ государственных академий на 2013–2020 гг. (ГП-14, раздел 63.3).
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Kandel E.R. The Neurobiology of Behavior / Principles of neural sciences. 4th Edition. Ed. Kandel E.R., Schwartz J.H., Jessel T.M. N.Y.: McGraw-Hill, 2000. 1414 p.
Vulliemoz S., Raineteau O., Jabaudon D. Reaching beyond the midline: why are human brains cross wired? // Lancet Neurol. 2005. V. 4. № 2. P. 87.
Phillips D.P., Brugge J.F. Progress in neurophysiology of sound localization // Annu. Rev. Psychol. 1985. V. 36. P. 245.
McAlpine D., Jiang D., Palmer A.R. A neural code for low frequency sound localization in mammals // Nat. Neurosci. 2001. V. 4. № 4. P. 396
Hart H.C., Palmer A.R., Hall D.A. Different areas of human non-primary auditory cortex are activated by sounds with spatial and nonspatial properties // Hum. Brain Mapp. 2004. V. 21. № 3. P. 178.
Spierer L., Bellmann-Thiran A., Maeder Ph. et al. Hemispheric competence for auditory spatial representation // Brain. 2009. V. 132. Pt. 7. P. 1953.
Fujiki N., Riederer K.A.J., Jousmäki V. et al. Human cortical representation of virtual auditory space: differences between sound azimuth and elevation // Eur. J. Neurosci. 2002. V. 16. № 11. P. 2207.
Palomäki K., Alku P., Mäkinen V. et al. Sound localization in the human brain: neuromagnetic observations // Neuroreport. 2000. V. 11. № 7. P. 1535.
Palomäki K.J., Tiitinen H., Mäkinen V. et al. Spatial processing in human auditory cortex: the effects of 3D, ITD, and ILD stimulation techniques // Cogn. Brain Res. 2005. V. 24. № 3. P. 364.
Krumbholz K., Schonwiesner M., von Cramon D.Y. et al. Representation of interaural temporal information from left and right auditory space in the human planum temporale and inferior parietal lobe // Cereb. Cortex. 2005. V. 15. № 3. P. 317.
Getzmann S. Effect of auditory motion velocity on reaction time and cortical processes // Neuropsychologia. 2009. V. 47. № 12. P. 2625.
Getzmann S. Auditory motion perception: onset position and motion direction are encoded in discrete processing stages // Eur. J. Neurosci. 2011. V. 33. № 7. P. 1339.
Tanaka H., Hachisuka K., Ogata H. Sound lateralisation in patients with left or right cerebral hemispheric lesions: relation with unilateral visuospatial neglect // J. Neurol. Neurosurg. Psychiatry. 1999. V. 67. № 4. P. 481.
Zatorre R.J., Mondor T.A., Evans A.C. Auditory attention to space and frequency activates similar cerebral systems // NeuroImage. 1999. V. 10. № 5. P. 544.
Brunetti M., Belardinelli P., Caulo M. et al. Human brain activation during passive listening to sounds from different locations: an fMRI and MEG study // Hum. Brain Mapp. 2005. V. 26. № 4. P. 251.
Tiitinen H., Salminen N.H., Palomaki K.J. et al. Neuromagnetic recordings reveal the temporal dynamics of auditory spatial processing in the human cortex // Neurosci. Lett. 2006. V. 396. № 1. P. 17.
Baumgart F., Gaschler-Markefski B., Woldorff M.G. et al. A movement-sensitive area in auditory cortex // Nature. 1999. V. 400. № 6746. P. 724.
Griffiths T.D., Rees G., Rees A. et al. Right parietal cortex is involved in the perception of sound movement in humans // Nat. Neurosci. 1998. V. 1. № 1. P. 74.
Krumbholz K., Hewson-Stoate N., Schönwiesner M. Cortical response to auditory motion suggests an asymmetry in the reliance on inter-hemispheric connections between the left and right auditory cortices // J. Neurophysiol. 2007. V. 97. № 2. P. 1649.
Briley P.M., Kitterick P.T., Summerfield A.Q. Evidence for Opponent Process Analysis of Sound Source Location in Humans // J. Assoc. Res. Otolaryngol. 2013. V. 14. № 1. P. 83.
Ahveninen J., Kopco N., Jääskeläinen I.P. Psychophysics and neuronal bases of sound localization in humans // Hear. Res. 2014. V. 307. P. 86.
Mesulam M.M. Spatial attention and neglect: parietal, frontal and cingulate contributions to the mental representation and attentional targeting of salient extrapersonal events // Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1999. V. 354. № 1387. P. 1325.
Teshiba T.M., Ling J., Ruhl D.A. et al. Evoked and intrinsic asymmetries during auditory attention: implications for the contralateral and neglect models of functioning // Cereb. Cortex. 2013. V. 23. № 3. P. 560.
Deouell L.Y., Bentin S., Giard M.H. Mismatch negativity in dichotic listening: Evidence for interhemispheric differences and multiple generators // Psychophysiol. 1998. V. 35. № 4. P. 355.
Kaiser J., Lutzenberger W., Preissl H. et al. Right-hemisphere dominance for the processing of sound-source lateralization // J. Neurosci. 2000. V. 20. № 17. P. 6631.
Schonwiesner M., Krumbholz K., Rubsamen R. et al. Hemispheric asymmetry for auditory processing in the human auditory brainstem, thalamus, and cortex // Cereb. Cortex. 2007. V. 17. № 2. P. 492.
Näätänen R. The role of attention in auditory information processing as revealed by event-related potentials and other brain measures of cognitive function // Behav. Brain Sci. 1990. V. 13. № 2. P. 201.
Kaiser J., Lutzenberger W. Location changes enhance hemispheric asymmetry of magnetic fields evoked by lateralized sounds in humans // Neurosci. Lett. 2001. V. 314. № 1–2. P. 17.
Näätänen R., Paavilainen P., Rinne T. et al. The mismatch negativity (MMN) in basic research of central auditory processing: A review // Clin. Neurophysiol. 2007. V. 118. № 12. P. 2544.
Nager W., Kohlmetz C., Joppich G. et al. Tracking of multiple sound sources defined by interaural time differences: brain potential evidence in humans // Neurosci. Lett. 2003. V. 344. № 3. P. 181.
Richter N., Schröger E., Rübsamen R. Hemispheric specialization during discrimination of sound sources reflected by MMN // Neuropsychologia. 2009. V. 47. № 12. P. 2652.
Takegata R., Huotilainen M., Rinne T. et al. Changes in acoustic features and their conjunctions are processed by separate neuronal populations // Neuroreport. 2001. V. 12. № 3. P. 525.
Tata M.S., Ward L.M. Early phase of spatial mismatch negativity is localized to a posterior “where” auditory pathway // Exp. Brain Res. 2005. V. 167. № 3. P. 481.
Вайтулевич С.Ф., Шестопалова Л.Б. Межполушарная асимметрия слуховых вызванных потенциалов человека и негативностей рассогласования при локализации источника звука // Журн. высшей нерв. деят. 2009. Т. 59. № 3. С. 269.
Вайтулевич С.Ф., Петропавловская Е.А., Шестопалова Л.Б. и др. Межполушарная асимметрия суммарной активности мозга человека при локализации источника звука // Сенсорные системы. 2015. Т. 29. № 2. С. 148.
Altman J.A., Vaitulevich S.Ph., Shestopalova L.B. et al. How does mismatch negativity reflect auditory motion? // Hear. Res. 2010. V. 268. № 1−2. P. 194.
Шестопалова Л.Б., Петропавловская Е.А., Вайтулевич С.Ф. и др. Топография вызванной активности мозга человека при различении движущихся звуковых стимулов // Журн. высшей нерв. деят. 2015. Т. 65. № 5. С. 577.
Shestopalova L.B., Petropavlovskaia E.A., Vaitulevich S.Ph. et al. Hemispheric asymmetry of ERPs and MMNs evoked by slow, fast and abrupt auditory motion // Neuropsychologia. 2016. V. 91. P. 465.
Magezi D.A., Krumbholz K. Evidence for opponent-channel coding of interaural time differences in human auditory cortex // J. Neurophysiol. 2010. V. 104. № 4. P. 1997.
Петропавловская Е.А., Шестопалова Л.Б., Вайтулевич С.Ф. Предсказательная способность слуховой системы при плавном движении и скачкообразном перемещении звуковых образов малой длительности // Журн. высшей нерв. деят. 2011. Т. 61. № 3. С. 293.
Delorme A., Sejnowski T., Makeig S. Enhanced detection of artifacts in EEG data using higher-order statistics and independent component analysis // NeuroImage. 2007. V. 34. № 4. P. 1443.
Barcelo F., Escera C., Corral M.J. et al. Task switching and novelty processing activate a common neural network for cognitive control // J. Cogn. Neurosci. 2006. V. 18. № 10. P. 1734.
Friedman D., Cycowicz Y.M., Gaeta H. The novelty P3: an event-related brain potential (ERP) sign of the brain’s evaluation of novelty // Neurosci. Biobehav. Res. 2001. V. 25. № 4. P. 355.
Rinne T., Särkkä A., Degerman A. et al. Two separate mechanisms underlie auditory change detection and involuntary control of attention // Brain Res. 2006. V. 1077. № 1. P. 135.
Horváth J., Winkler I. Distraction in a continuous-stimulation detection task // Biol. Psychol. 2010. V. 83. № 3. P. 229.
Альтман Я.А. Локализация движущегося источника звука. Л.: Наука, 1983. 176 с.
Альтман Я.А. Пространственный слух. СПб.: Институт физиологии им. И.П. Павлова РАН, 2011. 311 с.
Coull J.T. Neural correlates of attention and arousal: insights from electrophysiology, functional neuroimaging and psychopharmacology // Prog. Neurobiol. 1998. V. 55. P. 343.
Polich J. Updating P300: an integrative theory of P3a and P3b // Clin. Neurophysiol. 2007. V. 118. № 10. P. 2128.
Ball K., Sekuler R. Human Vision Favors Centrifugal Motion // Perception. 1980. V. 9. № 3. P. 317.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека