

Физиология человека, 2020, T. 46, № 6, стр. 78-91
Использование амплитудно-временнЫ́х параметров, связанных с событиями потенциалов мозга, в качестве индикаторов специфики процессов именования (номинации) и узнавания предмета по имени (денотации)
С. А. Исайчев 1, А. М. Черноризов 1, Т. В. Адамович 1, 4, А. В. Пилечева 1, *, А. А. Скворцов 1, 2, Ю. В. Микадзе 1, 2, 3
1 Московский государственный университет имени М.В. Ломоносова
Москва, Россия
2 ФГБОУ ВО Российский национальный исследовательский медицинский университет
имени Н.И. Пирогова Минздрава РФ
Москва, Россия
3 ФГБУ Федеральный центр мозга и нейротехнологий Федерального медико-биологического агенства РФ
Москва, Россия
4 ФГБНУ Психологический институт РАО
Москва, Россия
* E-mail: adita2010@yandex.ru
Поступила в редакцию 14.02.2020
После доработки 05.05.2020
Принята к публикации 01.08.2020
Аннотация
В статье представлены предварительные результаты экспериментального исследования специфики когнитивных процессов, связанных с именованием изображений (номинация) и узнаванием нужного изображения по слову-именованию (денотация), проведенного с использованием методов нейрофизиологии, психофизиологии и нейропсихологии. Отправной точкой для работы является описываемое в нейропсихологии расстройство называния (номинации) объекта, предъявляемого пациенту, сочетающееся с сохранным опознанием объекта (денотации) по его названию. Представление о сходстве основных компонентов, входящих в системную организацию когнитивных процессов, обеспечивающих номинацию и денотацию, опровергается этим фактом и свидетельствует о различиях в мозговой организации этих процессов, что проявляется в специфике их нарушений при локальных поражениях мозга. Пилотное исследование проводилось на выборке из 20 чел. (10 мужского и 10 женского пола в возрасте 18–23 лет) и включало в себя регистрацию: простой сенсомоторной реакции (СМР), электроэнцефалограммы (ЭЭГ), связанных с событиями потенциалов (ССП) и условной негативной волны (contingent negative variation – CNV) при предъявлении объектов и слов-названий. Статистический анализ динамики амплитудно-временны́х параметров основных компонентов ССП и CNV, регистрируемых в процессах номинации и денотации показал, что имеются достоверные различия в их конфигурации и топографии. Эти отличия могут отражать последовательную активность различных структур мозга в разные временны́е этапы реализации исследуемых функций речи.
С позиций психолингвистики, процессы именования изображения (номинативная функция речи) и чтения слов с последующим узнаванием соответствующего изображения (денотативная функция речи) содержат общие компоненты перцептивного (восприятие) и когнитивного (вербализация) свойства в их системной организации и различаются лишь характером взаимодействия между ними – от изображения объекта к слову или же от слова к изображению объекта [1, 2]. Из психолингвистических представлений о сходстве структурной организации номинации и денотации можно предположить, что локальные поражения мозга должны приводить к повреждению того или иного компонента или связи между ними и, как следствие, одновременному нарушению обеих функций речи. Однако имеются данные о расхождении между психолингвистическими моделями и данными нейропсихологии, согласно которым эти процессы имеют разную мозговую организацию, что проявляется в различиях характера и выраженности нарушений при локальных поражениях мозга. Наиболее яркий пример такого расхождения – явление “аномии”, т.е. трудности называния зрительно предъявляемого объекта в сочетании с сохранностью способности к выбору нужного объекта по его названию [3–6]. Можно предположить, что наряду со сходством когнитивных компонентов существует и определенная специфичность мозговых механизмов, обеспечивающих номинативную и денотативную функции речи. Полезным инструментом для выявления этой специфики могут быть активно используемые в нейролингвистике методы регистрации и анализа вызванных потенциалов (ВП) мозга: когнитивных вызванных потенциалов (КВП), связанных с событиями потенциалов (ССП) и условной негативной волны (CNV), позволяющих оценить особенности пространственно-временнóй организации нейрофизиологических механизмов когнитивных процессов, в том числе, процессов и функций речи [7, 8]. В истории уделяется большое внимание исследованиям ВП в нейролингвистике. В 1980 году M. Kutas и S.A. Hillyard описали “семантический” N400 компонент ВП [9]. В последующие годы были обнаружены дополнительные компоненты ВП, связанные с акустико-фонетическими, фонологическими, орфографическими, просодическими и синтаксическими процессами, а такие как различение фонем или сегментация слов соотносятся с более ранними (100–200 мс), быстрыми и автоматическими компонентами. Компоненты, отражающие процессы интеграции, имеют большие латенции (до 1 с). Например, P600 связывают с синтаксической обработкой [10]. Однако функциональная интерпретация N400 и P600 все еще остается предметом дискуссий. В работе F. Delogu et al. [11] был экспериментально проверен ряд конкурирующих теорий и сделаны выводы о том, что: компонент N400 отражает процесс поиска, а не интеграционные процессы; P600 отражает общие сложные интегративные, а не только синтаксические процессы; имеется пространственно-временнóе наложение компонентов N400 и P600. Некоторые авторы соотносят появление вышеперечисленных компонентов ВП с различными этапами развития речи в детском возрасте [12–15]. Сейчас уже становится очевидным, что большинство “лингвистических” компонентов ВП также могут быть связаны с неязыковыми функциями, т.е. неспецифичны, и вопрос себя далеко не исчерпал [16].
С позиций системной психофизиологии проблема специфичности отдельных компонентов ССП решается достаточно радикально. Эти компоненты могут отражать электрическую активность множества мозговых структур в данном временнóм интервале, а с другой стороны – их амплитудно-временнáя динамика может отражать участие в данный момент времени – разных психических феноменов, функций и когнитивных процессов. В качестве примера можно привести компонент P300. Величина амплитуды, латентного периода и топография этой волны связаны с принятием решения, значимостью стимула, ориентировочной реакцией, отдельными параметрами памяти, внимания, эмоций и часто коррелирует с наличием различных психологических нарушений [17]. То есть проблема специфичности психолингвистических функций и их отражение в параметрах компонентов ССП становится базисной проблемой для выявления и описания нейрофизиологических механизмов различных функций речи. Выходом из этой экспериментальной ситуации (парадигмы) может быть поиск комплекса психофизиологических маркеров, позволяющих дифференцировать собственно психолингвистические феномены от других когнитивных процессов. Одним из таких маркеров может быть латенция отдельных компонентов ССП и CNV. Известно, что латенция компонента в меньшей степени, чем амплитуда, зависит от экспериментальных условий и фоновой активности ЭЭГ. Временны́е аспекты взаимодействия между двумя стимулами, один из которых является предупреждающим, а другой – императивным, хорошо отражает динамика компонентов CNV.
CNV – эндогенный ВП, впервые описанный W.G. Walter et al. [18] и представляющий характерную волну, появление которой обусловлено внутренними процессами [19], происходящими между двумя взаимосвязанными для испытуемого событиями. Традиционно разделяют CNV, возникающий в процессе выполнения задачи, предполагающей моторный ответ [20–22], и задачи на восприятие временны́х интервалов [23–25]. Ряд авторов связывают формирование CNV с работой таламуса, стриатума и SMA (supplementary motor area), но дискуссия по этому вопросу продолжается [26]. Также отмечено влияние на отдельные параметры CNV работы дофаминергической системы мозга [27]. В своей обзорной работе T.W. Kononowicz et al. показали, что CNV отражает в основном общий подготовительный процесс, связанный с оптимизацией систем мозга и других когнитивных механизмов, которые зависят от конкретной задачи [28], а в качестве основной проблемы выделяют смешение побочных сигналов, сопровождающих запись электроэнцефалограммы с непосредственно когнитивными и перцептивными процессами. Там же авторы выделяют ряд вопросов в отношении CNV, которые по-прежнему остаются нерешенными, и, среди прочего, имеющие отношение к задаче: можно ли определить структуры мозга, которые обеспечивают формирование компонентов CNV, и их связь с механизмами номинации и денотации. В данной работе проводились регистрация и анализ основных компонентов ССП и CNV в экспериментах, где в качестве стимульного материала использовался стандартный психолингвистический набор “речевых задач” – называние картинки и подбор картинки в ответ на предъявленное слово. В статье излагаются результаты анализа амплитудно-временны́х параметров и топографического распределения ССП и CNV по скальпу, свидетельствующие о различиях пространственно-временнóй организации механизмов номинативной и денотативной функций речи.
МЕТОДИКА
В эксперименте принимали участие 20 студентов и аспирантов факультета психологии МГУ им. М.В. Ломоносова (10 мужского и 10 женского пола) в возрасте от 18 до 23 лет (n = 20, SD = 0.18). Все участники исследования являлись правшами, имели нормальное зрение, история неврологических заболеваний или травм головного мозга в анамнезе отсутствовала.
Экспериментальная процедура. С каждым участником проводили две серии экспериментов. В первой серии (называние объектов-стимулов – номинация) сначала предъявляли изображение объекта и через секунду – два слова, одно из которых соответствовало названию объекта (рис. 1, А). Во второй серии (выбор предъявляемых объектов-стимулов по названию – денотация) предъявляли слово и через секунду – два объекта, один из которых соответствовал слову (рис. 1, Б). Предъявление стимул-объекта и выбор из двух одновременно предъявляемых объектов нужного (целевого объекта) составляли одну пробу. Каждая экспериментальная серия включала 60 проб. Временнóй промежуток между пробами составлял 2 с. Объекты-стимулы предъявляли на мониторе персонального компьютера (ПК) с использованием программы презентации стимулов PsychoPy (https://www.psychopy.org). Выбор целевых объектов сопровождался нажатием на клавишу ПК.
Рис. 1.
Последовательность предъявления стимулов в двух экспериментальных сериях. а – демонстрация стимула (0.5 с); б – фон (1 с); в – демонстрация двойного стимула; г – выбор стимула; д – фон (2 с); е – повтор серии с другим стимулом; А – от изображения к слову. Б – от слова к изображению.
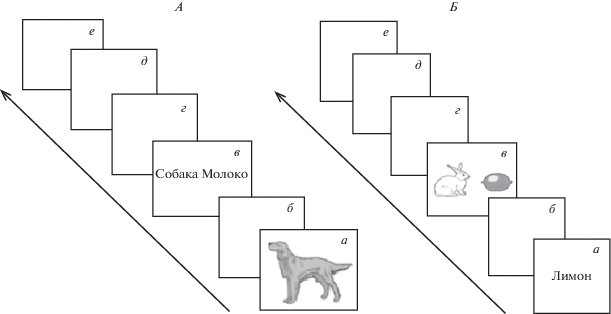
Подбор стимульного материала проводили с учетом норм, представленных в стандартных наборах [29, 30]. Экспериментальная процедура включала ряд этапов, во время которых проводили сбор дополнительной информации и подготовку к выполнению испытуемым основных задач эксперимента. 1 этап – регистрация фоновой активности ЭЭГ – 6 мин, сначала 3 мин с открытыми глазами, затем 3 мин с закрытыми глазами. 2 этап – выполнение задания на измерение латентного периода (ЛП) простой сенсомоторной реакции (СМР). 3 этап – выполнение задания “Изображение и два слова” (рис. 1, А – номинация). 4 этап – отдых 1 мин. 5 этап – выполнение задания “Слово и два изображения” (рис. 1, Б – денотация). 6 этап – регистрация фоновой активности ЭЭГ. Стимулы в условиях номинации и денотации демонстрировали следующим образом: 1) Предъявление первого (целевого) стимула – 0.5 с. 2) Фон – 1 с. 3) Предъявление двойного стимула – 2 с. 4) Фон – 2 с. 5) Повтор серии.
Ответы фиксировали с помощью клавиш “Влево” и “Вправо” на клавиатуре. Ответы давали правой рукой.
Половине респондентов задания предъявляли в порядке “Номинация” – “Денотация”, другой половине – в обратном порядке.
В ходе задания на сенсомоторную реакцию респонденту предъявляли на 0.3 с крест в центре экрана, от респондента требовалось нажать на клавишу “Пробел” как можно быстрее при появлении стимула на экране. Время между стимулами выбирали случайно от 1 до 4 с.
Перед экспериментом участнику давали подробную инструкцию его действий на каждом этапе и проводили тренировочную серию по основным этапам выполнения заданий.
Регистрация ЭЭГ. Параллельно с предъявлением стимульного материала проводили регистрацию ЭЭГ по 64 каналам с использованием оборудования и программного обеспечения фирмы Brain Products GmbH (Германия). Запись сигнала ЭЭГ осуществляли с помощью программы Brain Vision Recorder с использованием шлема BrainVision EasyCap, земляной и референтный электроды располагали на местах AFz и FCz соответственно. Частота дискретизации сигнала ЭЭГ при записи составляла 1000 Гц. Зарегистрированные в процессе экспериментальной процедуры сигналы ЭЭГ были предварительно обработаны с использованием программного обеспечения BrainVision Analyzer (версия 2.0, Brain Products). В частности, проводили фильтрацию в диапазоне 0.5–30 Гц, производили смену референта на усредненный, частоту дискретизации снижали до 250 Гц. Удаление артефактов производили вручную, удаление глазодвигательных артефактов производили с помощью анализа независимых компонент (ICA) После обработки ЭЭГ записи постстимульных эпох усредняли для выделения ССП и CNV для каждого испытуемого и, после усреднения, для экспериментальной группы в целом. Построение топографических карт распределения амплитуды ССП и CNV, зарегистрированных во время реализации номинативной и денотативной функций речи, и оценку значимости амплитудно-временны́х и пространственных различий между этими потенциалами выполняли в программной среде MNE для языка программирования Python при помощи функции mne.viz.plot_topomap [31, 32]. Для выявлений и статистической оценки величины ЛП при выполнении разных экспериментальных заданий использовали дисперсионный анализ ANOVA с использованием апостериорного анализа и критерия Тьюки.
Для сравнения и оценки различий двух ССП на предъявление изображений объектов и слов использовали однофакторный дисперсионный анализ (ANOVA) для повторных измерений.
Для сравнительного анализа распределения электрической активности мозга использовали методы топографического картирования и “кластерный пермутационный Т-тест” [33].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
В двух экспериментальных сериях был обнаружен ряд различий в параметрах поведенческих (ЛП СМР) и нейрофизиологических (отдельных, идентифицированных компонентов ССП и CNV) реакций, регистрируемых во время выполнения участниками задач по номинации и денотации.
Различия по времени реакции на предъявление визуальных простых и императивных стимулов при выполнении заданий в сериях “номинация”/“денотация”. Согласно классическим представлениям, общее время СМР складывается из вариативных и относительно стабильных компонентов, отражающих процессы возбуждения рецепторов, передачи сигнала в сенсомоторные отделы коры, выбора моторной программы и реализации мышечной реакции [34].
Эти процессы в значительной степени генетически детерминированы. Но если ставится задача реагирования на разные типы стимулов специфическим образом, то латентный период СМР увеличивается за счет активации когнитивных процессов (опознания, категоризации стимулов, принятия решения). И чем сложнее когнитивная задача, тем больше ЛП СМР. Таким образом, по увеличению ЛП можно оценить степень когнитивной нагрузки во время выполнения экспериментальных заданий.
Результаты сравнительного анализа ЛП СМР в различных экспериментальных сериях представлены в табл. 1 (усреднение данных для всех участников). Тест показал наличие значимых различий (F (2, 1674) = 477.8, p < 0.001) между ЛП СМР (Среднее = 0.33 с, SD = 0.07 с), денотацией (WtP) (Среднее = 0.43 с, SD = 0.11 с) и номинацией (PtW) (Среднее = 0.5 с, SD = 0.13 с). Усложнение заданий в сериях “денотация” (WtP) и “номинация” (PtW) приводит к увеличению ЛП СМР на 100 и 170 мс, соответственно.
Таблица 1.
Основные статистики времени реакции на предъявление визуальных простых и императивных стимулов
| Тип ВР\Значение | Время | Ст. откл. (SD) |
|---|---|---|
| Время реакции | 0.33 (330) | 0.07 (70) |
| WtP (денотация) | 0.43 (430) | 0.11 (110) |
| PtW (номинация) | 0.50 (500) | 0.13 (130) |
Результаты анализа свидетельствуют о статистической значимости различий (Ptukey < 0.001) между измерениями ЛП в разных экспериментальных сериях. Наиболее выражены различия между временем реакции в серии “номинация” (PtW) и простой СМР. Разница во времени между сериями “номинация” и “денотация” составляет 70 мс. Возможно, она отражает длительность когнитивной обработки на этапе “номинация”, т.е. тот факт, что это задание является более трудным. Эти результаты подтверждаются в работах [35, 36].
Различия амплитудно-временны́х параметров ССП и CNV при выполнении заданий в сериях “номинация”/“денотация”. В момент предъявления изображения объектов у каждого участника регистрировались ССП и CNV, которые затем усреднялись по экспериментальной группе из 20 чел. На рис. 2 представлены групповые усредненные ССП и CNV, характеризующие изменение электрической активности мозга во время выполнения задач на номинацию и денотацию.
Рис. 2.
Групповые усредненные ССП и CNV, характеризующие изменение электрической активности мозга во время прохождения экспериментальных серий “номинация” и “денотация”. 1 – сенсорные и когнитивные компоненты ССП; 2 – компоненты ССП и CNV; 3 – “фон” – ожидание продолжения стимульной серии. Положение электрода регистрации ССП и CNV - вертекс (Cz). Основные компоненты волны CNV и ССП: P1000-N1500-P1800-N2000-P2500 − отражающие активность структур, задействованные в реализации процессов номинации (а) и денотации (б).
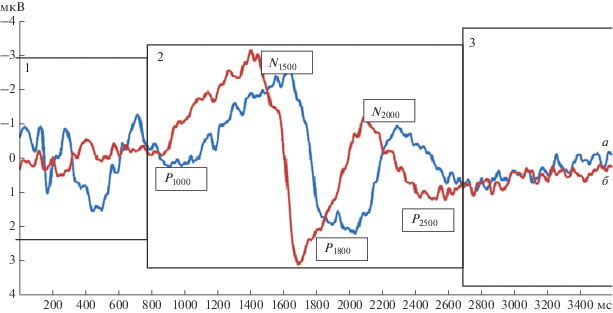
По динамике амплитуд и временны́х параметров, в частности минимальных различий показателей амплитуд отдельных компонентов ССП и CNV, и статистически значимой оценке конфигурации ВП можно выделить три временны́х блока, в которых отражаются основные различия выполнения заданий по реализации номинативной и денотативной функций речи.
Блок 1 (временнóй период от 0 до 800 мс). На этом этапе регистрируются компоненты сенсорных ВП (от 0 до 200 мс) и когнитивных ССП (от 200 до 800 мс) на предъявление первого стимула (“изображение объекта” или “слово”) в соответствующей стимульной серии, происходит его опознание и категоризация. Физические параметры стимула на сенсорном уровне отражаются в конфигурации и амплитудно-временны́х параметрах отдельных компонентов сенсорных ВП. В амплитудно-временны́х параметрах ССП (период от 200 до 800 мс) отражаются процессы опознания, категоризации стимулов объектов или слов, осуществляется их семантическая и синтаксическая обработка.
Блок 2 (временнóй период от 800 до 2700 мс). Период ожидания появления двойного императивного стимула (два слова или два изображения) и выбор соответствующего ответа. Здесь регистрируется сложный эндогенный потенциал – CNV, который отражает комплекс когнитивных процессов, связанных с ожиданием, опознанием, сравнением, категоризацией, выбором нужного стимула, принятием решения, подготовкой и реализацией адекватного моторного ответа или поведенческого акта. Различия в конфигурации ВП в данном блоке связаны с отличием когнитивных механизмов процессов денотации и номинации.
Блок 3 (временнóй период от 2700 до 3600 мс). Это фоновый этап – регистрация усредненной ЭЭГ-активности в отсутствие стимуляции во время пассивного ожидания продолжения экспериментальной серии. Этот этап характеризует фоновую постстимульную активность мозга и не содержит информативных характеристик ССП и CNV.
Стандартный анализ амплитудно-временны́х параметров ССП сводится к фиксации и оценке значимости различий идентифицированных компонентов ССП, которые отражают изменения в качестве или интенсивности используемого стимульного материала. На рис. 3 представлены усредненные по группе сенсорные и ССП, зарегистрированные в отведениях Oz и Pz на предъявление первого стимула в сериях выполнения заданий по номинации и денотации.
Рис. 3.
Сенсорные и когнитивные компоненты ССП на предъявление изображений и написанных слов. По оси Х – время в мс; по оси Y – амплитуда в мкВ; а – процесс номинации; б – процесс денотации; А – ССП, зарегистрированные в отведении Oz. Б – ССП, зарегистрированные в отведении Pz.
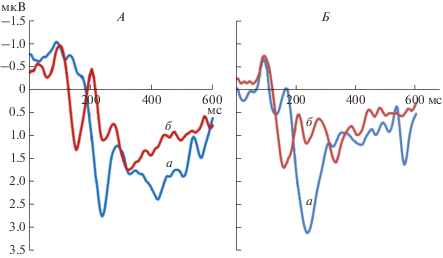
На рис. 3, А, Б видно, что конфигуративные параметры ССП от отведений Oz и Pz явно различаются. Меняется количество, латенция и амплитуда отдельных компонентов ССП. На этом уровне можно выделить два типа различий. Наиболее выражены различия в зависимости от качественных характеристик предъявляемых стимулов – “изображение” объектов или “слов”. Второй тип различий обусловлен топологическим положением отведений ЭЭГ. Чем дальше находятся отведения (электроды) от областей мозга, специфичных по отношению к обработке данного класса стимулов, тем больше проявляются различия в конфигуративных параметрах ССП. Эти два типа различий отражают временнýю и морфологическую специфику процессов реализации номинативной и денотативной функций речи на временнóм этапе от 0 до 600 мс.
Сравнительный анализ ССП в отведениях Oz и Pz во временны́х интервалах от 0–500 мс, и от 1500–2500 мс выявил значимые различия между двумя условиями:
1) для ССП в интервале 0–500 мс: средняя амплитуда PtW = 1.75, SD = 1.27; WtP = 0.88, SD = = 0.75; параметры ANOVA: F = 10.29, df = 219, p < < 0.05;
2) для ССП в интервале 1500–2500 мс: средняя амплитуда PtW = 0.00533, SD = 1.51; WtP = 0.54, SD = 1.3. Параметры ANOVA: F = 17.08, df = 485, p < 0.001.
Аналогичным образом можно сравнивать различные типы компонентов когнитивных ССП. Для этого выделяются устойчивые идентифицированные компоненты ССП, вычисляются их амплитудные и временны́е характеристики и проводится статистическая оценка различий между разными экспериментальными сериями. Выявляемые таким образом различия отражают локальный характер суммарной мозговой активности в отдельных отведениях, и полученные данные отражают активность и работу центров, которые расположены вблизи от регистрирующих электродов. Но если механизм психофизиологической функции представляет собой распределенную систему различных структур мозга, то такой подход не позволяет оценить активность всего комплекса мозговых структур, реализующих системные процессы исследуемых речевых функций. Таким образом, основная проблема анализа амплитудно-временны́х параметров ССП возникает тогда, когда выявляют мозговые структуры, участвующие в генерации основных компонентов ССП. Для решения этой задачи требуется учитывать не только амплитуду и латенцию основных компонентов ССП, но и анализировать динамику их пространственно-временны́х характеристик. Эта динамика может значительно меняться в зависимости от параметров стимуляции, расположения регистрирующих электродов, количества и активности возможных генераторов ЭЭГ в данном временнóм интервале. Иногда эти изменения довольно значительны, даже если регистрация ССП производится от близких по топологии областей поверхности головы. Так, наибольшая амплитуда сенсорных ВП на звуковые стимулы регистрируется в отведении Cz, а не в височных отведениях, соответствующих пространственному положению первичной слуховой коры.
Для выявления структур мозга, связанных с реализацией изучаемых системных психофизиологических функций речи, необходим многомерный анализ потенциальных генераторов электрической активности, который отражает системный характер организации этих функций. Первым шагом такого анализа является оценка временны́х и пространственных различий в организации мозгового обеспечения реализации функций номинации и денотации [37–44]. В данном исследовании для сравнительного анализа распределения электрической активности мозга использовались методы топографического картирования и “кластерный пермутационный Т-тест” [33].
Статистический анализ амплитудных, временны́х и пространственных различий ССП, регистрируемых в задачах на номинацию и денотацию, с помощью кластерного пермутационного Т-теста. В настоящее время кластерные пермутационные тесты получают признание в качестве логических процедур в когнитивной нейробиологии. Они решают проблему множественных сравнений многомерных МЭГ- и ЭЭГ-данных. Кластерный пермутационный тест относится к семейству пермутационных тестов, позволяющих решить проблемы ошибок I рода (число ошибочно отвергнутых гипотез), II рода (число ошибочно принятых гипотез) и множественных сравнений в многомерных данных. При анализе данных ЭЭГ существуют три домена – пространственное расположение, амплитуда и время, по которым могут наблюдаться различия. Использование кластерного пермутационного теста позволяет надежно оценить различия по всем доменам одновременно в различных экспериментальных условиях, решая при этом проблему множественных сравнений. Эта проблема не может быть решена в процессе применения одномерного амплитудного анализа ВП, который ограничен определенными отведениями и качественным описанием компонент. Таким образом, использование пермутационного теста позволяет надежно показать различия во всех трех доменах, что дополняет и подтверждает результаты, полученные с помощью других методов [33, 45].
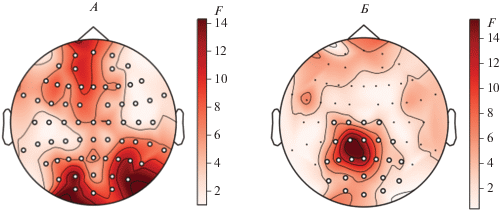
Кластерный пермутационный Т-тест показал наличие статистически значимых различий между условиями выполнения заданий в сериях “номинация” и “денотация”. Различия проявляются во временны́х диапазонах 676–1766 и 1730–2238 мс (рис. 4).
Рис. 4.
Карта кластерных различий между условиями выполнения заданий в сериях “номинация” и “денотация”. Контрастной шкалой обозначена F-статистика, круглыми точками – электроды, для которых различия считаются значимыми (p < 0.05). А – Кластер 1 – диапазон 676–1766 мс; Б – Кластер 2 – диапазон 1730–2238 мс.
Особеннсти топографического распределения активности мозга во время выполнения задач на номинацию и денотацию. Следует рассмотреть топограммы активности мозга во время реализации номинативной и денотативной функций речи (рис. 5). Топографические карты отражают суммарную активность областей мозга в различные временны́е интервалы с шагом 50 мс, что позволяет визуально и статистически выявить и оценить различия в пространственной локализации и временнóй последовательности активации зон мозга при реализации изучаемых функций речи.
Рис. 5.
Топография электрической активности мозга во время реализации номинативной и денотативной функций речи во временнóм диапазоне от 0 до 2950 мс с шагом 50 мс. А – предъявление “изображения” (серия “номинация”) или “слова” (серия “денотация”), временнóй диапазон 0–950 мс; Б – предъявление императивного стимула (ситуация выбора ответа) в виде “двух слов” (серия “номинация”) или двух “картинок” (серия “денотация”), временнóй диапазон 1000–1950 мс. Верхняя строка – денотация, нижняя строка – номинация. Контрастная шкала – микровольты (мкВ (uV)). В рамке выделен временнóй диапазон различий между двумя условиями.
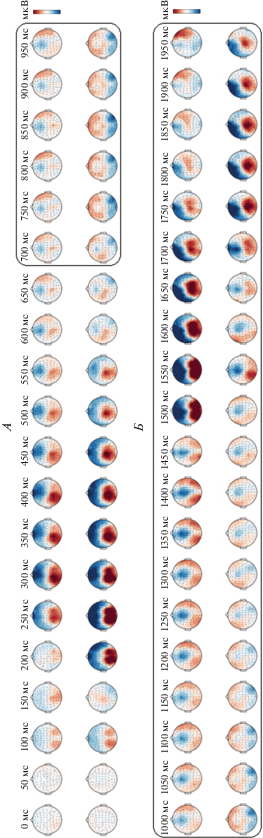
Основные различия активности мозга во время реализации номинативной и денотативной функций речи наблюдаются в двух временны́х диапазонах: 1-й – от 0 до 800 мс; 2-й – от 800 до 2700 мс. В первом диапазоне регистрируются сенсорные (от 0 до 250 мс) и когнитивные ВП (от 250 до 800 мс), которые отражают различия на подачу и оценку первого стимула (S1), а во втором диапазоне регистрируются компоненты CNV на подачу императивного стимула. По сравнению с анализом отдельных компонентов ССП (рис. 3) анализ различий по топографии дает нам дополнительную информацию о суммарной активности разных областей мозга в определенные временны́е периоды. При регистрации сенсорных компонентов ССП на подачу первого стимула на топографических картах (рис. 5, А) начало стимуляции сопровождается активацией окципитальных областей через 100 мс после подачи любого из стимулов (“слово” или “изображение”). Следует отметить, что предъявление первого стимула в серии “номинация” (“изображение”), по сравнению с первым стимулом в серии “денотация” (“слово”), характеризуется большей интенсивностью и величиной амплитуд основных компонентов этого ВП. Затем в затылочной области во временнóм диапазоне от 200 до 300 мс после подачи стимулов формируется фокус активации, который расширяется, охватывая полностью окципитально-париетальные области. Реакция на предъявление первых стимулов в обеих сериях заканчивается в диапазоне 550–600 мс после начала стимуляции. В целом активация на стимул “слово” характеризуется более слабой интенсивностью и более выраженной левосторонней асимметрией, чем активация на стимул “изображение”.
Во временнóм диапазоне от 700 до 2200 мс наблюдаются амплитудные и временны́е различия компонентов CNV, которые отражают специфику организации мозговых механизмов, реализующих собственно номинативную и денотативную функции речи. Один из таких компонентов – это “условная негативная волна” (УНВ, contingent negative variation – CNV) [46]. Использованная нами методика позволяла проводить совместную регистрацию ССП и CNV [47–51]. В качестве предупреждающего стимула (S1) использовался стимул “слово” или “изображение”. В качестве императивного стимула (S2) подавалась стимульная пара (два слова или два изображения), которая является сигналом к выбору ответа и сопровождается нажатием на кнопку клавиатуры. В ряде работ было показано, что, если интервал между S1–S2 достаточно велик (порядка – 1500 мс), то регистрируется мощная терминальная волна tCNV [52, 53], которая представляет собой негативный медленный компонент [28]. Этот компонент связан с интеграцией сенсорной информации и организацией ответной реакции на S2 [54, 55].
На рис. 2 представлены ССП и компоненты CNV, зарегистрированные при выполнении заданий на номинацию и денотацию. Комплекс компонентов с соответствующей латенцией P1000–N1500–P1800–N2000 отражает терминальные компоненты (tCNV) [56, 57]. Эти компоненты являются отражением, собственно активности мозговых структур, обеспечивающих процессы окончательной реализации номинативной и денотативной функций речи. Конфигуративно по количеству основных компонентов tCNV, регистрируемых при выполнении задач на номинацию и денотацию, различий нет (рис. 2, Блок 2).
Различие этих компонент CNV в двух сериях проявляется в латенции и амплитуде (рис. 2, Блок 2). Причем наиболее выражена разница в латенции этих основных компонентов. Отличия между аналогичными компонентами tCNV в сериях “номинация” и “денотация” по их латенции составляют: для N1500 – 212 мс, для P1800 – 352 мс, для N2000 – 220 мс. Различия в амплитуде этих компонентов менее выражены, чем различия в латенции. В серии “денотация” амплитуда (от пика до пика) компонентов N1500–P1800 составляет 6 мкВ. В серии номинация амплитуда аналогичного компонента N1620–P2000 составляет 4.9 мкВ. То есть оказывается, что амплитуда основных компонентов tCNV при денотации выше, чем при номинации. Но как было показано выше, на сенсорном уровне основные компоненты ССП на стимулы “изображение” отличаются большей амплитудой, чем аналогичные компоненты на стимулы “слово” (рис. 2, Блок 1). Увеличение амплитуды от пика до пика компонентов N1500–P1800 можно объяснить тем, что стимул “слово” вызывает более мощную активацию нейрональных структур, объединенных ассоциативными связями. Причем эта активация преимущественно охватывает лобно-теменные и премоторные области [54, 56, 58]. Таким образом, основное различие между компонентами CNV, зарегистрированными при выполнении заданий “номинация” и “денотация”, заключается в сдвиге (уменьшении) латенции основных компонентов CNV в серии “денотация” на 200–350 мс по отношению к серии “номинация”. Важно отметить, что стимульная пара (императивный стимул) появляется на экране монитора через 1500 мс после начала серии, где в качестве предупреждающего стимула (первого в серии) используется стимул “слово”. А максимальная реакция основного компонента P1500 в комплексе CNV наблюдается за 100 мс до подачи стимула S2. Это означает, что наблюдается выраженный эффект антиципации в случае, если в качестве предупреждающего стимула используется стимул “слово” [35, 59, 60].
В ряде исследований речевой деятельности был обнаружен эффект антиципации или прогнозирования при понимании языка на нескольких уровнях, в том числе и на уровне лексики [61]. В работе L. Magyari et al. была установлена связь между десинхронизацией β-ритма и антиципацией при использовании речевых оборотов, имеющих предсказуемые и непредсказуемые окончания [62]. Предсказуемые речевые обороты сопровождались снижением мощности в более медленном β-диапазоне (11–18.5 Гц) уже за 1250 мс до окончания высказывания, что отразило эффект ранней антиципации на основе обработки синтаксической, семантической и временнóй информации. В одном из своих исследований R.S. Gisladottir et al. регистрировали потенциалы, связанные с событиями, в то время, как испытуемые слушали диалоги и выполняли задачу категоризации действий. Анализ результатов определил появление ранней активности уже через 200 мс после начала целевого высказывания. Это указывает на то, что распознавание речевого акта идет с самого начала, когда высказывание обработано лишь частично [63]. В более поздней работе R.S. Gisladottir et al. показали, что предвосхищение в речевом акте может иметь решающее значение для стратегии речевого поведения, позволяя быстро извлекать элементы из лингвистического кода и реагировать в течение 200 мс, характерных для разговорной речи [64].
В исследовании мы получили надежные маркеры амплитудно-временны́х различий между основными компонентами CNV, которые свидетельствуют о специфике пространственно-временнóй организации нейрофизиологических механизмов, реализующих номинативную и денотативную функции речи.
Кроме различий в амплитуде и латенции основных компонентов CNV, регистрируемых в интервале 700–2200 мс, наблюдаются и различия в топографии этих компонентов (рис. 5). В серии “номинация” ожидание императивного стимула сопровождается синхронной симметричной активацией фронтальных и фронто-темпоральных областей, которая наблюдается во временнóм диапазоне от 750 до 1250 мс. Это типичная реакция CNV по интенсивности и по топологии на ожидание подачи императивного стимула [56, 65, 66].
В серии “денотация” (по сравнению с серией “номинация”) в этом же временнóм диапазоне можно наблюдать нарушение симметричности фронто-темпоральных локусов активации. В диапазоне от 700 до 1100 мс активируются сначала правые фронто-темпоральные области, затем в диапазоне от 1200 до 1400 мс фокус активации смещается в правую темпоральную область с одновременной активацией левой и правой окципитальных областей (рис. 5). В момент подачи императивного стимула (1500 мс) происходит максимальная активация в окципитально-париетальных областях. Такое распределение активности не соответствует топографическому распределению активности при номинации, значительно отличается от CNV по основным локусам активации и может интерпретироваться как специфическая реакция антиципации на предъявление стимула “слово”.
Демонстрация двойного стимула и реакции на него в сериях “номинация” и “денотация”, также имеют свою специфику. Предъявление двух слов в серии “номинация” вызывает ряд динамических изменений активации во временнóм диапазоне от 1500 (время подачи императивного стимула) до 1950 мс. Причем структура этих изменений сходна по пространственно-временны́м параметрам предъявлению стимула “слово” при выполнении задания “денотация”. Активация начинается в окципитально-париетальных областях левого полушария, затем ее фокус смещается в центр, где в диапазоне 1750–1800 мс достигает максимума интенсивности.
Специфичность реакции активации в серии “денотация” на предъявление императивного стимула заключается в том, что она начинается еще до реальной подачи этого стимула. За 200 мс до подачи стимула активируются окципитально-париетальные области левого полушария и окципитально–париетальные и темпоральные области правого полушария (рис. 5). В момент подачи стимула (1500 мс) активация достигает максимальных значений и сохраняется такой в течение 150–200 мс после подачи императивного стимула. Затем фокус активности смещается в центральную и правую лобно-височную область. Следует также отметить, что интенсивность активации в серии “денотация” на этом уровне в целом выше, чем в серии “номинация”. При этом на сенсорном уровне наблюдается противоположное соотношение – амплитуда компонентов ВП на предъявление стимулов “изображение” больше, чем на предъявление стимула “слово”.
ЗАКЛЮЧЕНИЕ
Основным результатом работы является установление специфики поведенческих реакций и активности мозга в процессах номинации и денотации. Задания в сериях “денотация” и “номинация” приводят к увеличению ЛП простой СМР на 100 и 170 мс, соответственно. Предположительно, разница во времени СМР между сериями “номинация” и “денотация” (70 мс) связана с увеличением сложности когнитивной обработки на этапе “номинация”. Это согласуется с другим обнаруженным фактом, состоящим в том, что основное различие между ССП, зарегистрированными при выполнении заданий “номинация” и “денотация”, заключается в увеличении ЛП основных компонентов ССП в серии “номинация” на 200–350 мс по отношению к серии “денотация”. Эти расхождения объяснимы. В задачах номинации для доступа к имени объекта требуется целенаправленный поиск ассоциации между его зрительным образом и обозначающим его словом. В работе C.M. Hamame et al. показано, что критически важной структурой, обеспечивающей этот ассоциативный процесс, является гиппокамп [67]. В экспериментах с участием больных эпилепсией с помощью интракраниально имплантированных электродов (по хирургическим показаниям) регистрировали суммарную электрическую активность (iEEG) непосредственно гиппокампа в процессе выполнения пациентом задачи по называнию картинки. В качестве показателей суммарной нейронной активности гиппокампа во время называния использовали значения спектральной мощности в диапазоне γ-ритма (50–150 Гц). Обнаружено, что латентный период активации гиппокампа является надежным предиктором латентного периода речевого ответа. В частности, пониженный уровень активности гиппокампа коррелирует с неудачами в назывании и феноменом “вертится на кончике языка” (“прескевю”). Таким образом, можно заключить, что гиппокамп является важным звеном нейронной сети номинации, и что динамика гиппокампальной активности тесно связана с эффективностью речепроизводства.
Кроме различий в амплитуде и латенции компонентов CNV и когнитивных ССП, регистрируемых в заданиях номинации и денотации в интервале 700–2200 мс, наблюдаются и различия в характере временнóй последовательности активации разных зон мозга при реализации изучаемых функций речи. Различия между ССП в заданиях на номинацию и денотацию в перцептивном диапазоне отражают, в основном, физические параметры стимулов (картинок или слов), а в когнитивном – процессы категоризации и принятия решения.
По данным кластерного пермутационного Т-теста, статистически значимые различия между условиями выполнения заданий в сериях “номинация” и “денотация” проявляются во временны́х диапазонах 676–1766 и 1766–2238 мс.
Результаты исследования свидетельствуют о том, что амплитудно-временны́е свойства ССП и пространственно-временны́е параметры их распределения по поверхности головы могут быть использованы в качестве информативных показателей в нейролингвистических исследованиях системной организации функций речи.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены локальным биоэтическим комитетом Этической комиссии факультета психологии МГУ им. М.В. Ломоносова (Москва).
Информированное согласие. Каждый участник исследования получил подробное разъяснение потенциальных рисков и преимуществ, а также характера предстоящего исследования и представил добровольное согласие.
Финансирование работы. Работа выполнена при поддержке РФФИ (проект № 18-013-00655).
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Indefrey P. The Spatial and Temporal Signatures of Word Production Components: A Critical Update // Front. In Psychology. 2011. V. 2. P. 255.
Васильева М. Ментальный лексикон: где же место морфологии? // Российский журн. когнитивной науки. 2014. Т. 1. № 4. С. 31.
Лурия А.Р. Основные проблемы нейролингвистики. Изд. 2-е. М.: Издательство ЛКИ, 2007. 256 с.
DeLeon I.G., Frank-Crawford M.A., Gregory M. et al. On the correspondence between preference assessment outcomes and progressive-ratio schedule assessments of stimulus value // J. Appl. Behav. Anal. 2009. V. 42. № 3. P. 729.
Бреус Е.В. Теория и практика перевода с английского на русский: учебное пособие. Часть 1. М.: Изд-во УРАО, 2001. 104 с.
Valente A., Pinet S., Alario F-X. et al. “When” Does Picture Naming Take Longer Than Word Reading? // Front. Psychol. 2016. V. 7. P. 31.
Rommers J., Federmeier K. Electrophysiological Methods / Research Methods in Psycholinguistics // Eds. De Groot A.M.B., Hagoort P. A Practical Guide. Hoboken, NJ: Wiley Blackwell, 2017. 392 p.
Микадзе Ю.В., Черноризов А.М., Скворцов А.А. и др. Модели и методы исследования переработки информации в процессах называния предмета и соотнесения названия с предметом // Экспериментальная психология. 2019. Т. 12. № 1. С. 153. Mikadze Yu.V., Chernorizov A.M., Skvortsov A.A. et al. Models and methods for the study of information processing in the processes of naming the subject // Experimental Psychology. 2019. V. 12. № 1. P. 153.
Kutas M., Hillyard S.A. Reading senseless sentences: Brain potentials reflect semantic incongruity // Science. 1980. V. 207. № 4427. P. 203.
Osterhout L., Holcomb P.J. Event-related brain potentials elicited by syntactic anomaly // J. Mem. Lang. 1992. V. 31. P. 785.
Delogu F., Brouwer H., Crocker M.W. Event-related potentials index lexical retrieval (N400) and integration (P600) during language comprehension // Brain Cogn. 2019. V. 135. P. 103569.
Holcomb P., Grainger J., O’Rourke T. An electrophysiological study of the effects of orthographic neighborhood size on printed word perception // J. Cogn. Neurosci. 2002. V. 14. № 6. P. 938.
Hahne A., Friederici A.D. Electrophysiological evidence for two steps in syntactic analysis: Early automatic and late controlled processes // J. Cogn. Neurosci. 1999. V. 11. № 2. P. 194.
Friederici A.D. Neurophysiological markers of early language acquisition: From syllables to sentences // Trends Cogn. Sci. 2006. V. 9. № 10. P. 481.
Mills D.L., Coffey-Corina S., Neville H.J. Language comprehension and cerebral specialization from 13 to 20 months // Dev. Neuropsychol. 1997. V. 13. № 3. P. 397.
Steinhauer K., Connolly J.F. Event-Related Potentials in the Study of Language / Handbook of the Neuroscience of Language // Eds. Stemmer B., Whitaker H.A. Academic Press, 2008. P. 191.
Александров И.О., Максимова Н.Е. Связанные с событиями потенциалы мозга (ССП) в психофизиологическом исследовании. Основы психофизиологии. Изд. 2-е, дополненное. СПб.: Питер, 2001. 496 с.
Walter W.G., Cooper R., Aldridge V.J. et al. Contingent negative variation: an electric sign of sensorimotor association and expectancy in the human brain // Nature. 1964. V. 203. P. 380.
Tecce J.J. Contingent negative variation (CNV) and psychological processes in man // Psychol. Bull. 1972. V. 77. № 2. P. 73.
Macar F., Vidal F., Casini L. The supplementary motor area in motor and sensory timing: evidence from slow brain potential changes // Exp. Brain Res. 1999. V. 125. P. 271.
Kononowicz T.W., Van Rijn H. Slow potentials in time estimation: the role of temporal accumulation and habituation // Front. Integr. Neurosci. 2011. V. 5. P. 48.
Kononowicz T.W., Sander T., Van Rijn H. Neuroelectromagnetic signatures of the reproduction of supra-second durations // Neuropsychologia. 2015. V. 75. P. 201.
Herbst S.K., Chaumon M., Penney T.B., Busch N.A. Flicker-induced time dilation does not modulate EEG correlates of temporal encoding // Brain Topogr. 2014. V. 28. № 4. P. 559.
Wiener M., Kliot D., Turkeltaub P.E. et al. Parietal influence on temporal encoding indexed by simultaneous transcranial magnetic stimulation and electroencephalography // J. Neurosci. 2012. V. 32. P. 12258.
Kononowicz T.W., Van Rijn H. Decoupling interval timing and climbing neural activity: a dissociation between CNV and N1P2 amplitudes // J. Neurosci. 2014. V. 34. P. 2931.
Plichta M.M., Wolf I., Hohmann S. et al. Simultaneous EEG and fMRI reveals a causally connected subcortical–cortical network during reward anticipation // J. Neurosci. 2013. V. 33. P. 14526.
Mallet N., Pogosyan A., Sharott A. et al. Disrupted dopamine transmission and the emergence of exaggerated beta oscillations in subthalamic nucleus and cerebral cortex // J. Neurosci. 2008. V. 28. P. 4795.
Kononowicz T.W., Penney T.B. The contingent negative variation (CNV): timing isn’t everything // Curr. Opin. Behav. Sci. 2016. V. 8. P. 231.
Snodgrass J.G., Vanderwart M. A standardized set of 260 pictures: Norms for name agreement, image agreement, familiarity, and visual complexity // J. Exp. Psychol.: Human Learning and Memory. 1980. V. 6. № 2. P. 174.
Григорьев А.А., Ощепков И.В., Балясникова О.В. и др. Нормативные данные по образности, согласованности наименований, знакомости и соответствию наименования изображению для набора из 286 стимулов // Вопросы психолингвистики. 2009. № 10. С. 128.
Gramfort A., Luessi M., Larson E. et al. MNE software for processing MEG and EEG data // NeuroImage. 2014. V. 1. № 86. P. 446.
Gramfort A., Luessi M., Larson E. et al. MEG and EEG data analysis with MNE-Python // Front. Neurosci. 2013. V. 7. № 267. P. 267.
Maris E., Oostenveld R. Nonparametric statistical testing of EEG- and MEG-data // J. Neurosci. Meth. 2007. V. 164. № 1. P. 177.
Anderson M., Nettelbeck T., Barlow J. Reaction time measures of speed processing: Speed of response selection increases with age but speed of stimulus categorization does not // Br. J. Dev. Psychol. 1997. V. 15. № 2. P. 145.
Riès S., Legou T., Burle B. et al. Why does picture naming take longer than word reading? The contribution of articulatory processes // Psychon. Bull. Rev. 2012. V. 19. P. 955.
Valente A., Pinet S., Alario F.-X., Laganaro M. “When” Does Picture Naming Take Longer Than Word Reading? // Front. Psychol. 2016. V. 7. № 31. P. 31.
Schwarzlose R.F., Swisher J.D., Dang S., Kanwisher N. The distribution of category and location information across object-selective regions in human visual cortex // PNAS. 2008. V. 105. № 11. P. 4447.
Weiner K.S., Grill-Spector K. Neural representations of faces and limbs neighbor in human high-level visual cortex: evidence for a new organization principle // Psychol. Res. 2013. V. 77. № 1. P. 74.
Peelen M.V., Downing P.E. Selectivity for the human body in the fusiform gyrus // J. Neurophysiol. 2005. V. 93. № 1. P. 603.
Kadipasaoglu C.M., Conner C.R., Whaley M.L. et al. Category-Selectivity in Human Visual Cortex Follows Cortical Topology: A Grouped icEEG Study // PLoS One. 2016. V. 11. № 6. P. e0157109.
Bracci S., Caramazza A., Peelen M.V. Representational Similarity of Body Parts in Human Occipitotemporal Cortex // J. Neurosci. 2015. V. 35. № 38. P. 12977.
Nobre A.C., Allison T., McCarthy G. Word recognition in the human inferior temporal lobe // Nature. 1994. V. 372. № 6503. P. 260.
Konkle T., Oliva A. A real-world size organization of object responses in occipitotemporal cortex // Neuron. 2012. V. 74. № 6. P. 1114.
Mollo G., Pulvermüller F., Hauk O. Movement priming of EEG/MEG brain responses for action-words characterizes the link between language and action // Cortex. 2016. V. 74. P. 262.
Maris E. Statistical testing in electrophysiological studies // Psychophysiology. 2012. V. 49. № 4. P. 549.
Walter W.G., Cooper R., Aldridge V.J. et al. Contingent Negative Variation: an electric sign of sensorimotor association and expectancy in the human brain // Nature. 1964. V. 203. № 4943. P. 380.
Rijn H., Kononowicz T.W., Meck W.H. et al. Contingent negative variation and its relation to time estimation: a theoretical evaluation // Front. Integr. Neurosci. 2011. V. 5. P. 91.
Kiroy V.N., Bakhtin O.M., Minyaeva N. et al. [Contingent negative variation during prospective activity] // Zh. Vyssh. Nerv. Deyat. Im. I.P. Pavlova. 2017. V. 67. № 3. P. 299.
Uysal U., Idiman F., Idiman E. et al. Contingent negative variation is associated with cognitive dysfunction and secondary progressive disease course in multiple sclerosis // J. Clin. Neurol. 2014. V. 10. № 4. P. 296.
Oishi M., Mochizuki Y., Du C. et al. Contingent negative variation and movement-related cortical potentials in parkinsonism // Electroencephalogr. Clin. Neurophysiol. 1995. V. 95. P. 346.
Drake M.E., Jr, Weate S.J., Newell S.A. Contingent negative variation in epilepsy // Seizure. 1997. V. 6. P. 297.
Kamijo K., Pontifex M.B., O’Leary K.C. et al. The effects of an afterschool physical activity program on working memory in preadolescent children // Dev. Sci. 2011. V. 14. №. 5. P. 1046.
Ludyga S., Gerber M., Kamijo K. et al. The effects of a school-based exercise program on neurophysiological indices of working memory operations in adolescents // J. Sci. Med. Sport. 2018. V. 21. № 8. P. 833.
Segalowitz S.J., Santesso D.L., Jetha M.K. Electrophysiological changes during adolescence: a review // Brain Cognition. 2010. V. 72. № 1. P. 86.
Flores A.B., Digiacomo M.R., Meneres S. et al. Development of preparatory activity indexed by the contingent negative variation in children // Brain Cognition. 2009. V. 71. № 2. P. 129.
Gomez C.M., Flores A., Ledesma A. Fronto-parietal networks activation during the contingent negative variation period // Brain Res. Bull. 2007. V. 73. № 1–3. P. 40.
Arjona A., Gómez C.M. Sequential Effects in the Central Cue Posner Paradigm: On-line Bayesian Learning / Cognitive Electrophysiology of Attention // Ed. George R. Mangun, 2014. P. 45.
Loveless N.E., Sanford A.J. Slow potential correlates of preparatory set // Biol. Psychol. 1974. V. 1. № 4. P. 303.
Анохин П.К. Системные механизмы высшей нервной деятельности: избранные труды. М.: Наука, 1979. 454 с.
Price C.J., McCrory E., Noppeney U. et al. How reading differs from object naming at the neuronal level // Neuroimage. 2006. V. 29. P. 643.
Kutas M., DeLong K.A., Smith N.J. A look around at what lies ahead: prediction and predictability in language processing. Predictions in the Brain: Using our Past to Generate a Future / Ed. Bar M. Oxford: Oxford University Press, 2011. P. 400.
Magyari L., Bastiaansen M., De Ruiter J.P., Levinson S.C. Early anticipation lies behind the speed of response in conversation // J. Cogn. Neurosci. 2014. V. 26. № 11. P. 2530.
Gisladottir R.S., Chwilla D.J., Levinson S.C. Conversation Electrified: ERP Correlates of Speech Act Recognition in Underspecified Utterances // PLoS One. 2015. V. 10. № 3. P. e0120068.
Levinson S.C. Action formation and ascription. The handbook of conversation analysis / Eds. Stivers T., Sidnell J. Malden, MA: Wiley- Blackwell, 2013. P. 844.
Babiloni C., Brancucci A., Vecchio F. et al. Anticipation of somatosensory and motor events increases centro-parietal functional coupling: An EEG coherence study // Clin. Neurophysiol. 2006. V. 117. № 5. P. 1000.
Bender S., Resch F., Weisbrod M. et al. Specific task anticipation versus unspecific orienting reaction during early contingent negative variation // Clin. Neurophysol. 2004. V. 115. № 8. P. 1836.
Hamame C.M., Vidal J.R., Perrone-Bertolotti M. et al. Functional selectivity in the human occipitotemporal cortex during natural vision: evidence from combined intracranial EEG and eye-tracking // Neuroimage. 2014. V. 95. P. 276.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека