

Физиология человека, 2020, T. 46, № 6, стр. 5-15
Действительно ли инструкция “быть оригинальным и придумывать” влияет на ЭЭГ-корреляты выполнения творческих задач?
Н. В. Шемякина 1, *, Ж. В. Нагорнова 1, **
1 ФГБУН Институт эволюционной физиологии и биохимии
имени И.М. Сеченова РАН
Санкт-Петербург, Россия
* E-mail: natalia.shemyakina@iephb.ru
** E-mail: nagornova.zhanna@iephb.ru
Поступила в редакцию 29.04.2020
После доработки 10.06.2020
Принята к публикации 01.08.2020
Аннотация
В работе приводятся данные связанной с событиями синхронизации/десинхронизации при выполнении вербальных творческих (придумать окончания пословиц) и нетворческих (предложить синоним окончания известной пословицы; вспомнить известное окончание пословиц) задач, а также данные классификации ЭЭГ-паттернов при выполнении испытуемыми заданных инструкций. В исследовании принимали участие 24 добровольца (18–22 года: 20 женщин, 4 мужчин). При придумывании оригинальных окончаний в сравнении с контрольной задачей на память были выявлены бóльшие значения мощности ЭЭГ в лобной области правого полушария в диапазоне частот 8–9 Гц в интервале 400–730 мс. В теменной области левого полушария, в этом же диапазоне частот, были получены бóльшие значения мощности электроэнцефалограммы (ЭЭГ) при создании и оригинальных, и синонимичных окончаний по сравнению с контрольной задачей “вспомнить”. При создании оригинальных и синонимичных окончаний мощность ЭЭГ была выше в центральных лобных областях коры для диапазона частот 14–15 Гц, a также в отведениях правого полушария F4, P4 для задач с синонимами по сравнению с задачей на память (850–950 мс). В более высокочастотном диапазоне 17–21 Гц различий между творческой задачей и задачей на синонимы не выявлено. При этом творческая задача, по сравнению с задачей на память, характеризовалась отличиями в теменных отведениях билатерально, а в центральной лобной зоне только для частот 17–18 Гц. Задача на синонимы, по сравнению с контролем, отличалась в указанных зонах для более широкого диапазона частот: 17–21 Гц. Классификация ЭЭГ-сигналов проводилась с использованием программного пакета classifier learning в среде matlab. Рассматривались результаты классификации на основе линейного дискриминантного анализа, метода опорных векторов и метода, давшего лучший результат классификации. Для классификации использовали преобразованный к плотности источника тока (CSD) сигнал ЭЭГ от лобных (F3, Fz, F4) и теменных отведений (P3, Pz, P4), расположенных на поверхности головы по системе 10–20. Средняя точность классификации по единичным пробам для 3-х типов заданий у всех испытуемых составила – 48.7 ± 5%[SD] для лучших классификаторов, лучший результат отдельного испытуемого достигал – 58.5% c использованием линейного дискриминантного анализа при теоретическом пороге случайной классификации 33%.
Творческое мышление достаточно давно исследуется психологами при изучении интеллекта и способностей человека [1, 2]. В современном обществе творческий потенциал является экономической ценностью, а реализованный творческий потенциал определяет качество жизни человека. Понимание физиологических основ нахождения творческих решений – актуальная задача для возможности самостоятельного управления этим процессом, например, на основе регуляции человеком своего состояния с использованием биологической обратной связи [3]. Нейрофизиологические корреляты творческой деятельности изучены сравнительно мало, даже при условии различных методических возможностей – электроэнцефалограммы (ЭЭГ), позитронно-эмиссионной томографии (ПЭТ), функциональной магнитно-резонансной томографии (фМРТ) и др. [4–6 и др.]. Основные направления исследования нейрофизиологии творческой деятельности – это изучение творческих состояний [7, 8], ассоциативных процессов [9, 10], этапов принятия творческих решений и инсайта [11–14].
В некоторых исследованиях предпринимаются попытки улучшать результаты когнитивной и творческой деятельности посредством различных видов воздействий, например, электрической стимуляции. Анодная tDCS повышает возбудимость коры, тогда как катодная – действует наоборот [15, 16]. В частности, было показано, что анодная tDCS стимуляция повышает точность рабочей памяти в течение 30 мин после стимуляции [17, 18]. В работе [19] обозначено, что выполнение творческих задач (тест отдаленных ассоциаций, тест альтернативного использования и др.) улучшалось при деактивации левого (катодная tDCS) и активации (анодная tDCS) правого полушария. Ранее было показано, что эмоциональный фон при творческой деятельности существенно влияет на ее корреляты [20–23], даже если речь идет просто об использовании эмоционально-значимых стимулов [24, 25].
Вместе с тем, чтобы модулировать состояние нахождения творческих решений, необходимо иметь представления о характеристиках ЭЭГ-сигнала, которые соответствуют тому или иному решению, что мы отчасти попытались выявить в нынешней работе, осуществляя анализ физиологических данных и их классификацию. Данное исследование является логическим продолжением комплексного изучения высших функций человеческого мозга, начатого академиком Н.П. Бехтеревой [26–28]. Предыдущие исследования показали зависимость физиологических коррелят высших функций от когнитивно-специфических характеристик заданий [29, 30], стратегий испытуемых [31, 32], включения дополнительных условий, например таких, как детекция ошибок [33].
Рис. 1.
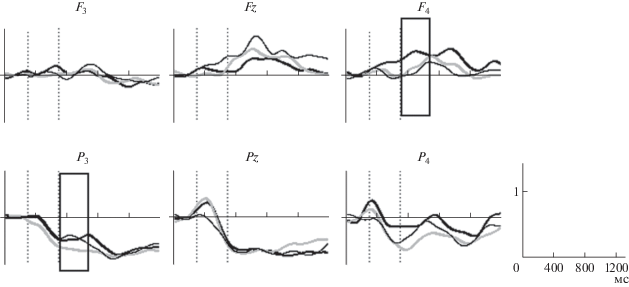
Различия мощности ЭЭГ в высокочастотных диапазонах (17–21 Гц) при выполнении заданий. Тв. – придумать оригинальное окончание пословицы (черная жирная линия), Син. – предложить синоним окончания (тонкая черная линия) и К. – вспомнить общеупотребимое окончание пословицы (серая линия). По оси х – время (мс), по оси у – мощность ЭЭГ-сигнала. Вертикальные линии – начало и окончание предъявления стимула (пословицы). Рамками обозначены временны́е интервалы значимых различий.
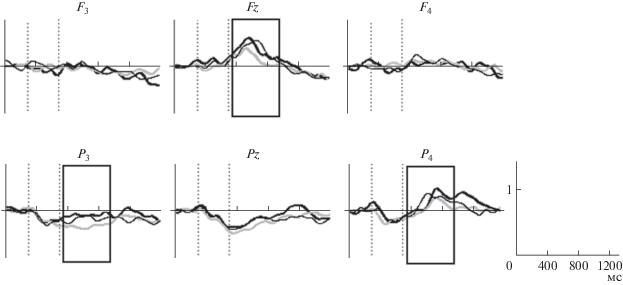
Рис. 2.
Различия мощности ЭЭГ в диапазоне 8–9 Гц при выполнении заданий. Тв. – придумать оригинальное окончание пословицы (черная жирная линия), Син. – предложить синоним окончания (тонкая черная линия) и К. – вспомнить общеупотребимое окончание пословицы (серая линия). По оси х – время (мс), по оси у – мощность ЭЭГ-сигнала. Вертикальные линии – начало и окончание предъявления стимула (пословицы). Рамками обозначены временны́е интервалы значимых различий.
Изучение “кратких” состояний принятия творческих или нахождения оригинальных решений часто выполняется в парадигме исследования вызванных потенциалов [12]. Исследования предполагают регистрацию и накопление определенного количества проб и, как правило, анализ ЭЭГ/ВП [12], или связанной с событиями синхронизации и десинхронизации (ERS/ERD) [34]. Мотивация на нахождение творческих решений может быть задана инструкцией – “быть творческим и оригинальным”, или за счет специфики стимульного материала [35]. В нашем исследовании испытуемые работали с известными пословицами и поговорками, к которым было необходимо придумать оригинальные завершения, отличающиеся от общепринятых так, чтобы изменить известный смысл пословиц на какой-то другой [29]. То есть сама инструкция предполагала ориентацию на преодоление стереотипа памяти и нахождение творческих решений. Использование коротких проб в исследованиях нахождения творческих решений, а также попытки найти информативные характеристики для классификации ЭЭГ-сигналов приближают нас к изучению real-life процессов, и таких исследований исключительно мало [36].
Информативным показателем изменений биоэлектрической активности мозга, дающим возможность получить дополнительную информацию о времени возникновения возможных различий, является частотно-временнóе преобразование сигнала ЭЭГ для анализа связанной с событием синхронизации/десинхронизации ЭЭГ [37, 38]. В настоящей работе изучали изменения связанной с событием синхронизации/десинхронизации ЭЭГ при придумывании человеком оригинальных окончаний к общеизвестным пословицам в сравнении с контрольными задачами нахождения синонимов и воспроизведения из памяти. Дополнительная задача исследования состояла в проверке возможности классификации ЭЭГ-проб при выполнении творческого и нетворческих заданий и в выявлении наиболее информативных признаков ЭЭГ-сигнала.
Сложность классификации ЭЭГ-сигналов связана с низким пространственным разрешением метода. Наибольшее количество работ по классификации ЭЭГ-паттернов приходится на работы, связанные с разработкой ИМК [39] и выделением паттернов реальных и воображаемых движений [40–42]. Количество работ по классификации ментальных состояний исключительно мало. Иногда ментальные состояния используют в сочетании с более понятными классифицируемыми сигналами. Например, для симуляции управления устройствами можно применять классификацию состояний решения ментальных задач (счет в уме) и моторного воображения [43] – т.е. состояний, вовлекающих разную “модальность” ментальных операций. Работы с классификацией электрофизиологических паттернов ментальных состояний часто связаны с попытками оценить степень усталости или когнитивной нагрузки либо классифицировать эмоциональные состояния по данным ЭЭГ [44, 45]. Отдельные исследования посвящены поиску возможности классификации ЭЭГ-сигналов при деятельности, вовлекающей одну модальность (вербальная, пространственно-образная, (в том числе, музыкальная)) в зависимости от творческого характера задания. В частности, в работе [46] классифицировались ЭЭГ-паттерны состояний музыкальной импровизации и “нетворческого” проигрывания гамм. При использовании общей пространственной фильтрации (CSP) в семи частотных диапазонах ЭЭГ и классификации двух состояний, точность классификации у 11 чел. из 14 превысила случайный порог и составила в среднем 75 ± 3.4%. В работе [47] была предпринята попытка выделения информативных признаков ЭЭГ-сигнала для классификации состояний дивергентного мышления и математического счета с использованием классификатора на основе метода опорных векторов (с различными базисными функциями).
Таким образом, работ по классификации ЭЭГ-паттернов, связанных с нахождением творческих решений, практически нет. Вместе с тем, поиск информативных признаков для выделения паттернов ЭЭГ, связанных с выполнением творческой задачи, крайне важен.
Цель данного исследования состояла в сопоставлении и совместном анализе физиологических данных связанной с событием синхронизации/десинхронизации и оценки применимости средств классификации для распознавания ЭЭГ-паттернов при выполнении вербальных заданий, различающихся уровнем вовлечения творческого мышления для преодоления стереотипа. Для реализации данной цели было предложено оригинальное задание, предполагающее три уровня преодоления стереотипа долговременной памяти: вспомнить окончание пословицы (нет преодоления стереотипа), предложить синоним окончания (без изменения семантической составляющей стереотипа) и создание оригинального окончания с задачей – изменить первоначальный смысл пословицы (преодоление стереотипа долговременной памяти).
МЕТОДИКА
Всего в ЭЭГ-исследовании принимали участие 24 испытуемых (в возратсе 18−22 лет, 20 женщин, 4 мужчин) с образованием выше среднего, родным языком являлся русский. На момент исследования испытуемые были здоровы и не проходили курсов медикаментозного лечения, декларировали отсутствие хронических заболеваний.
Задания. Испытуемым предлагали творческое задание (Тв.) – придумать оригинальные окончания общеизвестных пословиц и поговорок, изменяя их известный смысл на какой-то другой [29]. В качестве контрольных задач испытуемым предлагали назвать (вспомнить) общеизвестный вариант окончания предъявленной неоконченной пословицы или поговорки (К.) или назвать синоним окончания пословицы (Син.). Во всех предложенных испытуемым заданиях присутствовало извлечение информации из долговременной памяти. Отличие творческой задачи от нетворческой состояло в преодолении ранее сформированного стереотипа и поиске (придумывании), в соответствии с заданием, нового оригинального окончания пословицы или поговорки, существенно меняющего ее смысл.
Во время всего исследования испытуемые располагались в удобном кресле, на расстоянии 1.5 м от стандартного 17-дюймового LCD монитора, на котором предъявляли пословицы без последнего слова (средние угловые размеры: 4.9′ × 2.1′), написанные черным жирным шрифтом Times New Roman (цвет экрана – белый).
Испытуемые должны были нажимать на кнопку, если находили ответ, и озвучивать его после предъявления знака вопроса в конце пробы. С помощью установленного в камере микрофона исследователь слышал ответы испытуемого и фиксировал их для дальнейшей оценки качества найденных решений. Длительность проб составила 5400 мс с рандомизированными межпробными интервалами (1500–2500 мс).
Внутри одного блока задания испытуемым предъявляли 104 известные пословицы. Порядок выполнения блоков задания, как и порядок проб в заданиях, были рандомизированы среди испытуемых. Для анализа использовали только пробы без миографических артефактов.
Для каждого типа задач оценивали субъективную сложность (по 10-бальной шкале, где 1 – минимальная сложность, а 10 – максимальная) и эмоции по шкале (–3; +3).
Процедура регистрации ЭЭГ. Регистрацию ЭЭГ проводили с помощью 32-х канального цифрового электроэнцефалографа “Мицар” (ООО “Мицар”, Россия) посредством программного пакета WinEEG (Пономарев В.А., Кропотов Ю.Д., № государственной регистрации 2 001 610 516 от 08.05.2001) от 31 AgCl электродов, расположенных по системе 10–10. Референтный электрод располагался на мочках обоих ушей, заземляющий электрод – в передне-центральном отведении на поверхности головы. Сопротивление электродов не превышало 5 кОм, частота квантования сигналов составила 500 Гц. ЭЭГ анализировали в полосе 0.53–30 Гц.
Предобработка сигнала. В ЭЭГ записях испытуемых помечали и исключали из дальнейшего анализа пробы с артефактами движения глаз, с медленными волнами (0–1 Гц с амплитудой больше 50 мкВ), быстрыми волнами (20–35 Гц с амплитудой выше 35 мкВ), фрагменты ЭЭГ с амплитудой сигнала больше 100 мкВ [48]. Артефакты, связанные с движением глаз, удаляли при помощи обнуления соответствующих независимых компонент [49, 50]. Далее данные преобразовывались к так называемой плотности источника тока (current source density (CSD) по [51]), [52–54]) для последующего анализа связанной с событием синхронизации/десинхронизации и классификации проб во время выполнения творческих и нетворческих задач. Алгоритм CSD преобразования был реализован в программном пакете регистрации и обработки данных ЭЭГ (WinEEG Пономарев В.А., Кропотов Ю.Д., № государственной регистрации 2 001 610 516 от 08.05.2001). Преобразование ЭЭГ-сигнала к плотности источников тока позволяет уменьшить воздействие эффекта объемного проведения на сигнал, регистрируемый с поверхности головы и локализовать источники ЭЭГ-сигналов [52, 53]. Как показано в работе [54], использование данного преобразования в сравнении с анализом ЭЭГ в референтном монтаже позволяет выделять локальные различия, которые в ином случае могут быть маскированы активностью соседних зон коры.
Для сравнительного анализа и последующей классификации ЭЭГ-паттернов использовали фрагменты безартефактных проб с биоэлектрической активностью, зарегистрированной от лобных и теменных областей коры: отведения F3, Fz, F4 и теменных отведений – P3, Pz, P4 согласно модифицированной системе 10–20. Для анализа изменений вызванной синхронизации/десинхронизации ЭЭГ использовали данные 20 испытуемых, число усредненных проб у которых было не менее 30 на каждый тип стимула. Для классификации использовали данные 15 чел. с количеством проб в среднем для классов: Тв. (придумать оригинальное окончание пословицы): 60 ± 19 проб, К. (вспомнить общеизвестное окончание пословицы): 59 ± 14 проб, Син. (придумать синоним окончания пословицы): 49 ± 9 проб.
Для анализа изменений вызванной мощности ЭЭГ при многократном предъявлении задач – вспомнить, найти синоним или придумать окончание пословиц, было использовано вейвлет-преобразование Морле. Методика расчета представлена в работе [55]. Были рассчитаны частотно-временны́е характеристики сигнала ЭЭГ для эпохи анализа 1800 мс (300 мс – предстимульный интервал, 400 мс – длительность предъявления стимула и 1100 мс после предъявления стимула – начало выполнения инструкции). Интервал 1500 мс после предъявления стимула был выбран на основе анализа времени нахождения ответа испытуемыми в простой контрольной задаче – вспомнить окончание пословиц, и не пересекался со временем подготовки к произвольному нажатию (согласно литературным данным, за ~500 мс произвольному нажатию предшествует медленная негативная волна [56, 57]). Вейвлет преобразование вычисляли для различных центральных частот в диапазоне 2–30 Гц с шагом 1 Гц с шириной вейвлета 5 циклов. Вычисляли усредненные по группам проб оценки спектральной мощности для каждого испытуемого, каждого электрода и каждой группы проб по отдельности. Значения показателей, характеризующих динамику мощности сигналов испытуемых, усреднялись. Для уменьшения влияния компонент вызванных потенциалов на связанную с событием синхронизацию/десинхронизацию ЭЭГ ВП вычисляли и вычитали из каждого фрагмента ЭЭГ. По разностным частотно-временны́м картам были определены частотные диапазоны и интервалы различий для исследованных состояний в областях интереса – лобных и теменных зонах коры. Статистический анализ усредненных значений мощности для выбранных временны́х окон проводили при помощи дисперсионного анализа ANOVA для повторных измерений с применением поправки Хьюнха-Фельдта [58]. В качестве факторов рассматривали: СОСТОЯНИЕ (3 типа заданий), ЗОНА (6 отведений) и их взаимодействие.
Анализ поведенческих показателей (результатов самоотчета и времени ответа в заданиях) проводили при помощи непараметрических критериев – рангового дисперсионного анализа Фридмана и конкордации Кендалла.
Методы классификации. Для классификации ЭЭГ-сигналов использовали опции программного пакета classifier learning в среде matlab. В качестве “эталонных” рассматривали результаты классификации на основе линейного дискриминантного анализа, метода опорных векторов на основе радиальной базисной функции и результаты метода, давшего лучший процент классификации для отдельного испытуемого. Как упоминалось ранее, в качестве одного из инструментов классификации применяли предложенный В. Вапником и А. Червонкисом метод опорных векторов [59]. Этот метод относится к методам линейной классификации и заключается в разделении выборки на классы с помощью оптимальной разделяющей гиперплоскости, уравнение которой в общем случае имеет вид: f(x) = = $(\omega ,(x)) + b,$ где $\omega = \sum\nolimits_{i = 1}^N {{{\lambda }_{i}}{{y}_{i}}\left( {{{x}_{i}}} \right)} ,$ коэффициенты ${{{\lambda }_{i}}}$ зависят от ${{{y}_{i}}}$ (векторов меток класса принадлежности) и от значения скалярных произведений $\left( {\left( {{{x}_{i}}} \right){\text{,}}\left( {{{x}_{j}}} \right)} \right).$ Таким образом, для нахождения решающей функции необходимо знать значения скалярных произведений. Преобразования данных определяются функцией-ядром: K(x, y) = = $\left( {\varphi \left( x \right),\varphi \left( y \right)} \right).$ На основании результатов исследований по выбору предпочтительного типа SVM [60, 61] для классификации сигналов ЭЭГ в настоящей работе в качестве функции-ядра рассматривали радиальную базисную функцию Гаусса: $K\left( {{{x}_{i}}{\text{,}}{{x}_{j}}} \right)$ = ${\text{exp}}\left( { - \gamma {{{\left\| {{{x}_{i}} - {{x}_{j}}} \right\|}}^{2}}} \right),$ для $\gamma > 0.$
Эмпирическую оценку обобщающей способности алгоритмов проводили автоматически при помощи кросс-валидации: общее число проб разделяли последовательно на 5 выборок, четыре из которых (80% проб) входили в обучающую, а одна (20% проб) – в тестовую выборки. Обучающая и тестовая выборки проб не пересекались. Для уменьшения размерности входного вектора признаков использовали анализ главных компонент, встроенный в пакет matlab.
Статистический анализ результатов 3-х классовой классификации проводили с использованием дисперсионного анализа ANOVA при рассмотрении фактора ТИП КЛАССИФИКАТОРА.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Поведенческие данные. Среднее время выполнения творческой задачи, т.е. время до обозначения испытуемым нахождения решения с помощью нажатия на кнопку, составляло 3345 ± 414 мс, подбор синонимов окончания пословиц занимал 3020 ± 426 мс, среднее время выполнения контрольной задачи на воспроизведение окончаний пословиц и поговорок из памяти – 2261 ± 418 мс от момента предъявления стимула. Таким образом, для анализа был выбран интервал 1800 мс, состоящий из 300 мс – предстимульного интервала и 1500 мс от момента предъявления стимула. Выбранный интервал не включал время нажатия на кнопку. Время нахождения решений достоверно различалось для разных заданий в группе исследованных испытуемых: ДА χ2 (n = 20, сс = 2) = = 40.0. p < 0.001, коэфф. конкордации = 1.0, средн. ранг r = 1.0. Показатели субъективной сложности выполняемых задач различались (ДА χ2 (n = 20, сс = 2) = 12.8 p < 0.002, коэфф. конкордации = 0.3, средн. ранг r = 0.28) и составляли: сложность выполнения творческой задачи – 7.5 ± 1.8, сложность подбора синонимов окончания пословиц – 6.3 ± 1.9, сложность контрольной задачи на воспроизведение окончания по памяти – 5.3 ± 2.1. Субъективный уровень эмоций между заданиями достоверно не отличался.
Связанная с событиями синхронизация/десинхронизация. Выполнение творческой задачи и поиск синонимов окончаний пословиц сопровождались увеличением синхронизации ЭЭГ в диапазоне частот 17–21 Гц на интервале 450–1050 мс после предъявления стимула (рис. 1), и частот 14–15, 15–16 Гц в поздних интервалах 850–950 и 1180–1300 мс после предъявления стимула. После предъявления стимула были выявлены бóльшие значения мощности ЭЭГ в частотном диапазоне 8–9 Гц – в теменной области левого полушария при создании оригинальных и синонимичных окончаний по сравнению с контрольной задачей вспомнить окончание. Мощность этих же частот в лобной области правого полушария (отведение F4) была выше при придумывании оригинальных окончаний (рис. 2).
Результаты статистического анализа приведены в табл. 1.
Таблица 1.
Различия вызванной синхронизации/десинхронизации ЭЭГ в короткие интервалы после предъявления стимула в заданиях: придумать оригинальное окончание пословицы (Тв.), найти синоним окончания (Син.), и вспомнить общеизвестное окончание пословицы (К.)
| Частота (Гц) | Временнóй интервал (мс) | Достоверность эффектов | Области различий (апостериорный анализ): эффект | ||
|---|---|---|---|---|---|
| Тв. vs К. | Син. vs К. | Тв. vs Син. | |||
| 8–9 | 400–730 | С × З: F10,190 = 2.0, e(H-F) = 0.88, p < 0.05 | F4, P3: Тв. > K. | P3: Син. > К. | P3: Син. > Тв. |
| 14–15 | 850–950 | С: F2,38 = 5.8, e(H-F) = 0.88, p < 0.01 | Fz: Тв. > K. | Fz, F4, P4: Син. > К. | n.s. |
| 15–16 | 1180–1300 | С: F2,38 = 5.5, e(H-F) = 1.0, p < 0.01 | Pz, P4: Тв. > K. | n.s. | n.s. |
| 17–18 | 450–1050 | С: F2,38 = 5.8, e(H-F) = 1.0, p < 0.01 | Fz, P3, P4: Тв. > K. | Fz, P3, P4: Син. > К. | n.s. |
| 18–21 | 450–1050 | С: F2,38 = 7.2, e(H-F) = 0.88, p < 0.01 | P3, P4: Тв. > K. | Fz, P3, P4: Син. > К. | n.s. |
Примечание: C – фактор СОСТОЯНИЕ, С × З – взаимодействие факторов СОСТОЯНИЕ × ЗОНА, е(H-F) – поправка Хьюнха-Фельдта; Fz, F4, P3, Pz, P4 – положение электродов на поверхности головы (согласно системе 10/20); знак > обозначает, в каком из сравниваемых заданий мощность ЭЭГ в соответствующем частотно-временнóм диапазоне выше. n.s. – согласно апостериорному анализу достоверных различий между заданиями не наблюдалось.
В рамках реализации “single-trial” подхода проведена классификация единичных проб на основе сырых преобразованных к плотности источника тока данных ЭЭГ для общего временнóго интервала 1500 мс после предъявления стимула для каждого испытуемого. Результаты 3-х классовой классификации на основе сигналов от лобных (F3, Fz, F4) и теменных отведений (P3, Pz, P4) на интервале 1500 мс после предъявления стимула представлены в табл. 2.
Таблица 2.
Результаты 3-х классовой классификации ЭЭГ-паттернов при выполнении творческих и нетворческих задач
| Испытуемый | Точность классификации (%) | |||
|---|---|---|---|---|
| линейный дискриминантный анализ (LDA) | метод опорных векторов (Fine Gaussian SVM) и другой вариант SVM c наилучшими результатами | лучший классификатор | ||
| Исп. 1 | 40.1 | 41.2 Fine Gaussian | 44.4 | Ensemble (Subspace KNN) |
| Исп. 2 | 43.2 | 47.1 Fine Gaussian | 47.1 | SVM Fine Gaussian\Сubic SVM |
| Исп. 3 | 46.7 | 43.8 Fine Gaussian 46.2 Cubic |
46.7 | LDA |
| Исп. 4 | 45.6 | 45.1 Fine Gaussian 47.5 Quadrant\Cubic |
47.5 | Quadratic\Cubic SVM |
| Исп. 5 | 49.3 | 40.5 Fine 48.6 Quadrant |
49.3 | LDA |
| Исп. 6 | 44.8 | 46.9 Fine Gaussian 46.9 Linear\Cubic |
46.9 | Fine Gaussian\Linear\Cubic SVM |
| Исп. 7 | 43.6 | 35.3 Fine Gaussian 44.2 Cubic |
44.2 | Cubic SVM |
| Исп. 8 | 37.8 | 37.8 Fine Gaussian 39.5 Cubic |
40.7 | Quadratic DA\Fine KNN |
| Исп. 9 | 54.7 | 45.1 Fine Gaussian 48.4 Quadratic\Cubic |
54.7 | LDA |
| Исп. 10 | 42.3 | 49.2 Fine Gaussian | 49.2 | Fine Gaussian SVM |
| Исп. 11 | 44.6 | 40.0 Fine Gaussian 46.9 Cubic |
46.9 | Cubic SVM |
| Исп. 12 | 58.5 | 48.3 Fine Gaussian 58.0 Quadratic |
58.5 | LDA |
| Исп. 13 | 39.9 | 41.1 Fine Gaussian | 44.9 | Ensemble (Boosted trees) |
| Исп. 14 | 50.0 | 38.9 Fine Gaussian 50.0 Quadratic |
54.6 | Quadratic DA\ Ensemble (subspace discriminant) |
| Исп. 15 | 54.4 | 50.0 Fine Gaussian 52.9 Quadratic |
54.4 | LDA\ Ensemble (subspace discriminant) |
| Средняя точность классификации | 46 ± 6%[SD] | 43 ± 4%[SD] Fine Gaussian 48 ± 5%[SD] Other SVM |
48 ± 5%[SD] | |
У всех испытуемых точность классификации единичных проб (“single-trial” подход) с использованием указанных протестированных классификаторов превышала случайный уровень. Средняя максимальная точность классификации составляет 48 ± 5%[SD], максимальный результат среди испытуемых – 54.4% (ЛДА – линейный дискриминантный анализ) и 54.6% (Quadratic ДА). При статистическом анализе результатов классификации выявлен достоверный эффект фактора “ТИП КЛАССИФИКАТОРА”: F2.28 = 11.1, e(H-F) = = 0.62, p < 0.003. Апостериорный анализ выявил значимо более высокие точности классификации при использовании ЛДА и рассмотрении “лучшего классификатора” по сравнению с SVM на основе радиальной базисной функции. При этом точности ЛДА и “лучшего классификатора” значимо не различались.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В проведенном исследовании предпринята попытка выделения нейрофизиологических различий и классификации проб при работе с вербальным материалом (пословицы без последнего слова) в зависимости от поставленной инструкции – быть оригинальным и придумать окончание пословицы, меняющей ее смысл (Тв.), предложить синоним окончания – т.е. придумать новое окончание без изменения смысла пословицы (Син.) и вспомнить общеизвестное окончание пословицы (К.). Согласно поведенческим данным (время ответа) и результатам самоотчетов задания расположились в соответствии со схемой “многоуровневого контроля”: время выполнения и сложность контрольной задачи было минимальным, среднее время выполнения и средняя сложность соответствовали задаче, придумать синоним окончания пословицы, и бóльшее время выполнения и сложность соответствовали творческой задаче с инструкцией быть оригинальным. При этом анализ нейрофизиологических данных выявил различия и схожесть процессов создания меняющего смысл окончания пословиц и создания синонимичного ответа. Различия вызванной синхронизации/десинхронизации ЭЭГ проявляются во временнóм интервале – начиная с 400 мс после предъявления стимула – вероятно, отражая процессы, связанные с семантическим анализом [62, 63]. Можно предположить, что при восприятии знакомой пословицы без последнего слова на первых этапах, вне зависимости от инструкции, вспоминается окончание пословицы, и в дальнейшем в зависимости от инструкции – это окончание либо актуализуется, либо “подавляется” при ответе. Таким образом, отличия вызванной синхронизации/десинхронизации ЭЭГ после 400 мс могут быть связаны с разницей между процессами актуализации и подавлением семантической памяти. При этом поиск оригинального окончания пословицы связан с необходимостью задействовать широкую семантическую сеть ассоциативных связей. Запуск этих процессов происходит уже при восприятии стимула и обусловлен выполняемой инструкцией (так как стимульный материал для всех типов заданий одинаков). Обращает на себя внимание схожесть вызванных ответов при подборе синонимичного окончания в ряде эффектов с контрольной деятельностью, в других – с творческой деятельностью. Так в диапазоне α1-ритма (8–9 Гц) в теменных областях мощность ЭЭГ была выше при поиске нового окончания (синонимичного и оригинального), в то время как в лобной области правого полушария мощность ЭЭГ была выше при творческой деятельности по сравнению как с контрольной деятельностью, так и с поиском синонимов. Увеличение мощности ЭЭГ в диапазоне частот α1-ритма в правой лобной области может соотноситься с необходимостью торможения семантической сети общеизвестного окончания пословицы: было показано, что произвольное забывание в экспериментальных условиях вовлекает правые лобные и левые теменные области [64], при этом в произвольное забывание вовлечены области средней и нижней лобной извилины правого полушария [65]. Можно предположить, что в нашем исследовании при творческой деятельности наблюдается произвольное активное подавление семантической сети долговременной памяти, связанное с вовлечением лобных областей правого полушария, и запуск поиска ответа в широкой ассоциативной сети, предполагающий активацию фронтопариетальной системы мультимодальных третичных областей коры. По всей видимости, активация сетей, обеспечивающих долговременную семантическую память, наблюдается и при поиске синонимов окончания пословиц, что обуславливает похожую картину изменений вызванной синхронизации ЭЭГ в высокочастотных диапазонах при поиске оригинальных и синонимичных окончаний.
Таким образом, можно предположить, что создание оригинального (меняющего смысл пословицы) ответа имеет сходство с поиском синонимичного окончания (иного, нежели общеизвестного, но не меняющего смысл пословицы) в активации сетей долговременной семантической памяти, затрагивающей мультимодальные области теменной и лобной коры, и отличается от данной и контрольной деятельности торможением ассоциативного семантического поля общеизвестного смысла пословицы, что, по всей видимости, требует вовлечения лобных областей коры правого полушария.
В работе предпринята также попытка классификации отдельных проб (single-trial) при восприятии пословиц без последнего слова в зависимости от инструкций. Для классификации использовались “сырые” данные, преобразованные к плотности источника тока (current source density) от шести целевых электродов, расположенных в лобных и теменных зонах коры – F3, Fz, F4, P3, Pz, P4. В результате классификации проб у всех испытуемых были получены точности классификации, превышающие порог случайного уровня классификации (теоретический порог при 3-х классах = = 33.3%). При этом сравнение “лучшего классификатора”, автоматически выбираемого в пакете “classifier learner” в среде matlab и “классических” методов классификации – линейного дискриминантного анализа и метода опорных векторов (на основе радиальной базисной функции) продемонстрировали отсутствие значимых отличий точности “лучшего” классификатора и ЛДА. Таким образом, при групповом подходе классификатором выбора будет ЛДА. В целом точности классификации были не очень высоки (максимальный процент 54.6%), что свидетельствует о необходимости предобработки сигнала и выделения информативных признаков для классификации, которые могут быть индивидуальными [41, 66], и использования других средств классификации, возможно – на основе искусственных нейронных сетей, которые смогут продемонстрировать большую точность при классификации единичных проб [40]. Однако и при использовании “классических” средств классификации продемонстрирована принципиальная различимость состояний творческой и нетворческой деятельности при использовании отдельных “реализаций”.
ЗАКЛЮЧЕНИЕ
В проведенном исследовании продемонстрирована возможность взаимодополняющих подходов рассмотрения физиологических данных – анализа результатов для выявления физиологических эффектов и успешной классификации проб, предполагающих творческую и нетворческую вербальную деятельность. Достигнутая средняя точность классификации по единичным пробам 3-х типов заданий – поиск оригинального, меняющего смысл пословицы, окончания, поиск синонимичного окончания пословицы и вспоминание общеизвестного окончания пословицы – 46 ± 6%[SD] (максимальная точность – 54.6%). Применение анализа вызванной синхронизации/десинхронизации ЭЭГ с использованием вейвлет-преобразования продемонстрировало как черты сходства процессов поиска оригинального и синонимичного окончания пословицы по сравнению с контрольной деятельностью – увеличение мощности ЭЭГ в диапазоне частот 17–21 Гц, так и их отличия – топографические особенности различий для мощности в диапазоне 8–9 и 15–16 Гц. Это может свидетельствовать о запуске дополнительных механизмов когнитивного контроля при творческой деятельности – торможении непроизвольных ассоциаций долговременной памяти с вовлечением лобных отделов правого полушария и активации мультимодальных областей фронтопариетальной системы для вербального поиска в широком семантическом поле.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Этическим комитетом Института эволюционной физиологии и биохимии им. И.М. Сеченова РАН (Санкт-Петербург).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа поддержана РФФИ 19-015-00412a.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Guilford J.P. The nature of human intelligence. Mc-Graw-Hill, 1967. 538 p.
Guilford J.P. Higher-Order Structure-Of-Intellect Abilities // Multivariate Behav. Res. 1981. V. 16. № 4. P. 411.
Gruzelier J.H. EEG-neurofeedback for optimising performance. II: creativity, the performing arts and ecological validity // Neurosci Biobehav. Rev. 2014. V. 44. P. 142.
Fink A., Benedek M., Grabner R.H. et al. Creativity meets neuroscience: experimental tasks for the neuroscientific study of creative thinking // Methods. 2007. V. 42. № 1. P. 68.
Arden R., Chavez R.S., Grazioplene R., Jung R.E. Neuroimaging creativity: a psychometric view // Behav. Brain Res. 2010. V. 214. № 2. P. 143.
Dietrich A., Kanso R. A review of EEG, ERP, and neuroimaging studies of creativity and insight // Psychol. Bull. 2010. V. 136. № 5. P. 822.
Lopata J.A., Nowicki E.A., Joanisse M.F. Creativity as a distinct trainable mental state: An EEG study of musical improvisation // Neuropsychologia. 2017. V. 99. P. 246.
Shemyakina N.V., Nagornova Zh.V. EEG “Signs” of Verbal Creative Task Fulfillment with and without Overcoming Self-Induced Stereotypes // Behav. Sci. (Basel). 2020. V. 10. № 1. P. 17.
Razumnikova O.M. Creativity related cortex activity in the remote associates task // Brain Res. Bull. 2007. V. 73. № 1–3. P. 96.
Marron T.R., Lerner Y., Berant E. et al. Chain free association, creativity, and the default mode network // Neuropsychologia. 2018. V. 118(Pt A). P. 40.
Martindale C., Hasenfus N. EEG differences as a function of creativity, stage of the creative process, and effort to be original // Biol. Psychol. 1978. V. 6. № 3. P. 157.
Qiu J., Li H., Yang D. et al. The neural basis of insight problem solving: an event-related potential study // Brain Cogn. 2008. V. 68. № 1. P. 100.
Bowden E.M., Jung-Beeman M. Aha! Insight experience correlates with solution activation in the right hemisphere // Psychon. Bull. Rev. 2003. V. 10. № 3. P. 730.
Tik M., Sladky R., Luft C.D.B. et al. Ultra-high-field fMRI insights on insight: Neural correlates of the Aha!-moment // Hum. Brain Mapp. 2018. V. 39. № 8. P. 3241.
Bindman L., Lippold O., Redfearn J.W.T. The action of brief polarizing currents on the cerebral cortex of the rat (1) during current flow and (2) in the production of long-lasting after-effects // J. Physiol. 1964. V. 172. № 3. P. 369.
Jaušovec N., Jaušovec K. Increasing working memory capacity with theta transcranial alternating current stimulation (tACS) // Biol. Psychol. 2014. V. 96. P. 42.
Ohn S.H., Park C.I., Yoo W.K. et al. Time-dependent effect of transcranial direct current stimulation on the enhancement of working memory // Neuroreport. 2008. V. 19. № 1. P. 43.
Zaehle T., Sandmann P., Thorne J.D. et al. Transcranial direct current stimulation of the prefrontal cortex modulates working memory performance: combined behavioural and electrophysiological evidence // BMC Neurosci. 2011. V. 12. № 2. https://doi.org/10.1186/1471-2202-12-2
Hertenstein E., Waibel E., Frase L. et al. Modulation of creativity by transcranial direct current stimulation // Brain Stimul. 2019. V. 12. № 5. P. 1213.
Fink A., Schwab D., Papousek I. Sensitivity of EEG upper alpha activity to cognitive and affective creativity interventions // Int. J. Psychophysiol. 2011. V. 82. № 3. P. 233.
Subramaniam K., Beeman M., Faust M., Mashal N. Positively valenced stimuli facilitate creative novel metaphoric processes by enhancing medial prefrontal cortical activation // Front. Psychol. 2013. V. 4. P. 211.
Subramaniam K., Kounios J., Parrish T.B., Jung-Beeman M. A brain mechanism for facilitation of insight by positive affect // J. Cogn. Neurosci. 2009. V. 21. № 3. P. 415.
Subramaniam K., Vinogradov S. Improving the neural mechanisms of cognition through the pursuit of happiness // Front. Hum. Neurosci. 2013. V. 7. P. 452.
Шемякина Н.В., Данько С.Г. Изменения мощности и когерентности бета2-диапазона ЭЭГ при выполнении творческих заданий с использованием эмоционально-значимых и эмоционально-нейтральных слов // Физиология человека. 2007. Т. 33. № 1. С. 27. Shemyakina N.V., Dan’ko S.G. Changes in the power and coherence of the β2 EEG band in subjects performing creative tasks using emotionally significant and emotionally neutral words // Human Physiology. 2007. V. 33. № 1. P. 20.
Шемякина Н.В., Данько С.Г. Влияние эмоциональной окраски воспринимаемого сигнала на электроэнцефалографические корреляты творческой деятельности // Физиология человека. 2004. Т. 30. № 2. С. 22. Shemyakina N.V., Dan’ko S.G. Influence of the Emotional Perception of a Signal on the Electroencephalographic Correlates of Creative Activity // Human Physiology. 2004. V. 30. № 2. P. 145.
Бехтерева Н.П. Нейрофизиологические аспекты психической деятельности человека. 2-е издание, переработанное и дополненное. Л.: Медицина, 1974. 151 с. Bechtereva N.P. The Neurophysiological Aspects of Human Mental Activity. N.Y.: Oxford University Press, 1978. 181 p.
Бехтерева Н.П., Гоголицын Ю.Л., Кропотов Ю.Д., Медведев С.В. Нейрофизиологические механизмы мышления. Л.: Наука, 1985. 272 с.
Бехтерева Н.П. Здоровый и больной мозг человека. 2-е издание, переработанное и дополненное. Л.: Наука, 1988. 262 с.
Шемякина Н.В., Данько С.Г., Нагорнова Ж.В. и др. Динамика спектров мощности и когерентности ритмических компонентов ЭЭГ при решении вербальной творческой задачи преодоления стереотипа // Физиология человека. 2007. Т. 33. № 5. С. 14. Shemyakina N.V., Danko S.G., Nagornova Zh.V. et al. Changes in the power and coherence spectra of the EEG rhythmic components during solution of a verbal creative task of overcoming a stereotype // Human Physiology. 2007. V. 33. № 5. P. 524.
Данько С.Г., Шемякина Н.В., Нагорнова Ж.В., Старченко М.Г. Сравнение воздействия субъективной сложности и творческого характера вербальной деятельности на спектральные показатели мощности ЭЭГ // Физиология человека. 2009. V. 35. № 3. С. 132. Danko S.G., Shemyakina N.V., Nagornova Zh.V., Starchenko M.G. Comparison of the effects of the subjective complexity and verbal creativity on EEG spectral power parameters // Human Physiology. 2009. V. 35. № 3. P. 381.
Bechtereva N.P., Korotkov A.D., Pakhomov S.V. et al. PET study of brain maintenance of verbal creative activity // Int. J. Psychophysiol. 2004. V. 53. № 1. P. 11.
Razumnikova O.M., Bryzgalov A.O. Frequency-spatial organization of brain electrical activity in creative verbal thought: the role of the gender factor // Neurosci. Behav. Physiol. 2006. V. 36. № 6. P. 645.
Bechtereva N.P., Shemyakina N.V., Starchenko M.G. et al. Error detection mechanisms of the brain: background and prospects // Int. J. Psychophysiol. 2005. V. 58. № 2–3. P. 227.
Jung-Beeman M., Bowden E.M., Haberman J. et al. Neural activity when people solve verbal problems with insight // PLoS Biology. 2004. V. 2. № 4. P. 500.
Howard-Jones P.A., Blakemore S.J., Samuel E.A. et al. Semantic divergence and creative story generation: an fMRI investigation // Brain Res. Cogn. Brain Res. 2005. V. 25. № 1. P. 240.
Beaty R.E. The neuroscience of musical improvisation // Neurosci. Biobehav. Rev. 2015. V. 51. P. 108.
Pfurtsheller G., Aranibar A. Event-related cortical desynchronization detected by power measurement of scalp EEG // Electroencephalography and Clinical Neurophysiology. 1977. V. 42. № 6. P. 817.
Kiebel S., Tallon-Baudry C., Friston K.J. Parametric analysis of oscillatory activity as measured with EEG/MEG // Hum. Brain Mapp. 2005. V. 26. № 3. P. 170.
Wolpaw J.R., Wolpaw E.W. Brain-computer interfaces: principles and practice. N.Y.: Oxford University Press, 2012. 400 p.
Sonkin K.M., Stankevich L.A., Khomenko J.G. et al. Development of electroencephalographic pattern classifiers for real and imaginary thumb and index finger movements of one hand // Artif. Intell. Med. 2015. V. 63. № 2. P. 107.
Станкевич Л.А., Сонькин К.М., Шемякина Н.В. Классификация ЭЭГ-паттернов воображаемых движений пальцами одной руки, выполняемых в заданном ритме // Физиология человека. 2016. Т. 42. № 1. С. 40. Stankevich L.A., Sonkin K.M., Shemyakina N.V. et al. EEG pattern decoding of rhythmic individual finger imaginary movements of one hand // Human Physiology. 2016. V. 42. № 1. P. 32.
Frolov A.A., Bobrov P.D. Brain–Computer Interfaces: Neurophysiological Bases and Clinical Applications // Neurosci. Behav. Physiol. 2018. V. 48. № 9. P. 1033.
Kwon J., Shin J., Im C.H. Toward a compact hybrid brain-computer interface (BCI): Performance evaluation of multi-class hybrid EEG-fNIRS BCIs with limited number of channels // PLoS One. 2020. V. 15. № 3. P. e0230491.
Craik A., He Y., Contreras-Vidal J.L. Deep learning for electroencephalogram (EEG) classification tasks // J. Neural Eng. 2019. V. 16. № 3. P. 031001.
Craik A., He Y., Contreras-Vidal J.L. Deep learning for electroencephalogram (EEG) classification tasks: a review // J. Neural Eng. 2019. V. 16. № 3. P. 031001.
Sasaki M., Iversen J., Callan D.E. Music Improvisation Is Characterized by Increase EEG Spectral Power in Prefrontal and Perceptual Motor Cortical Sources and Can be Reliably Classified From Non-improvisatory Performance // Front. Hum. Neurosci. 2019. V. 13. P. 435.
Petersen T.H., Puthusserypady S. Assessing tDCS Placebo Effects on EEG and Cognitive Tasks / 41st Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC). Berlin, Germany, 2019. P. 4509.
Kozhushko N.J., Nagornova Zh.V., Evdokimov S.A. et al. Specificity of spontaneous EEG associated with different levels of cognitive and communicative dysfunctions in children // Int. J. Psychophysiol. 2018. V. 128. P. 22.
Jung T.P., Makeig S., Humphries C. et al. Removing electroencephalographic artifacts by blind source separation // Psychophysiology. 2000. V. 37. № 2. P. 163.
Терещенко Е.П., Пономарев В.А., Кропотов Ю.Д., Мюллер А. Сравнение эффективности различных методов удаления артефактов морганий при анализе количественной электроэнцефалограммы и вызванных потенциалов // Физиология человека. 2009. Т. 35. № 2. С. 124. Tereshchenko E.P., Ponomarev V.A., Kropotov Y.D., Müller A. Comparative efficiencies of different methods for removing blink artifacts in analyzing quantitative electroencephalogram and event-related potentials // Human Physiology. 2009. V. 35. № 2. P. 241.
Perrin F., Pernier J., Bertrand O., Echallier J.F. Spherical splines for scalp potential and current density mapping [Corrigenda EEG 02274, EEG Clin. Neurophysiol. 1990. 76. 565] // Electroencephalogr. Clin. Neurophysiol. 1989. V. 72. № 2. P. 184.
Tenke C.E., Kayser J. Generator localization by current source density (CSD): implications of volume conduction and field closure at intracranial and scalp resolutions // Clin. Neurophysiol. 2012. V. 123. № 12. P. 2328.
Kayser J., Tenke C. Issues and considerations for using the scalp surface Laplacian in EEG/ERP research: A tutorial review // Int. J. Psychophysiol. 2015. V. 97. № 3. P. 189.
Ponomarev V.A., Mueller A., Candrian G. et al. Group Independent Component Analysis (gICA) and Current Source Density (CSD) in the study of EEG in ADHD adults // Clin. Neurophysiol. 2014. V. 125. № 1. P. 83.
Tallon-Baudry C., Bertrand O. Oscillatory gamma activity in humans and its role in object representation // Trends Cogn. Sci. 1999. V. 3. № 4. P. 151.
Deecke L., Grözinger B., Kornhuber H.H. Voluntary finger movement in man: cerebral potentials and theory // Biol. Cybern. 1976. V. 23. № 2. P. 99.
Libet B., Gleason C.A., Wright E.W., Pearl D.K. Time of conscious intention to act in relation to onset of cerebral activity (readiness-potential). The unconscious initiation of a freely voluntary act // Brain. 1983. V. 106. P. 623.
Huynh H., Feldt L.S. Estimation of the Box Correction for Degrees of Freedom from Sample Data in Randomized Block and Split-Plot Designs // J. Educ. Stat. 1976. V. 1. P. 69.
Cortes C., Vapnik V.N. Support-Vector Networks // Machine Learning. 1995. V. 20 № 3. P. 273.
Сонькин К.М., Станкевич Л.А., Хоменко Ю.Г. и др. Классификация электроэнцефалографических паттернов воображаемых и реальных движений пальцев одной руки методом опорных векторов // Тихоокеанский медицинский журн. 2014. Т. 2. С. 30.
Shawe-Taylor J., Cristianini N. Kernell methods for pattern analysis. Cambridge University Press, 2004. 478 p.
Kutas M., Hillyard S.A. Brain potentials during reading reflect word expectancy and semantic association // Nature. 1984. V. 307. № 5947. P. 161.
Cermolacce M., Scannella S., Faugère M. et al. “All that glitters is not … alone”. Congruity effects in highly and less predictable sentence contexts // Neurophysiol. Clin. 2014. V. 44. № 2. P. 189.
Hanslmayr S., Leipold P., Pastötter B., Bäuml K.H. Anticipatory signatures of voluntary memory suppression // J. Neurosci. 2009. V. 29. № 9. P. 2742.
Depue B.E., Curran T., Banich M.T. Prefrontal regions orchestrate suppression of emotional memories via a two-phase process // Science. 2007. V. 317. № 5835. P. 215.
Sonkin K.M., Stankevich L.A., Khomenko Ju.G. et al. Neurological Classifier Committee Based on Artificial Neural Networks and Support Vector Machine for Single-Trial EEG Signal Decoding / Lecture Notes in Computer Science book series (LNCS, V. 9719). Advances in Neural Networks. ISNN-2016. P. 100.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека