

Физиология человека, 2021, T. 47, № 1, стр. 53-62
Особенности синергетического взаимодействия скелетных мышц при выполнении сложной координационной двигательной задачи
С. А. Моисеев 1, *, Р. М. Городничев 1
1 ФГБОУ ВО Великолукская государственная академия
физической культуры и спорта
Великие Луки, Россия
* E-mail: sergey_moiseev@vlgafc.ru
Поступила в редакцию 06.02.2020
После доработки 25.03.2020
Принята к публикации 20.06.2020
Аннотация
У двенадцати самбистов изучали особенности пространственно-временнóй организации мышечных синергий с применением методов выявления главных компонент, оценки авто- и кросскорреляционных функций электромиограмм (ЭМГ) скелетных мышц при выполнении броска “захватом ног” в разных его периодах. Установлено, что ЭМГ-активность обширных синергий меняется в зависимости от величины проявляемых мышечных усилий, характерных для разных периодов выполняемого движения. Синергетические взаимоотношения скелетных мышц оказываются пластичными в отношении их пространственной и временнóй организации, что обеспечивает надежное управление двигательной функцией в различных периодах выполнения движения.
Проблема сокращения избыточности степеней свободы нервно-мышечного аппарата в управлении произвольной двигательной активностью остается актуальным вопросом физиологии человека. Один из физиологических механизмов решения этой проблемы может заключаться в объединении сегментов тела, мышц и центральных нервных структур в функциональные образования – двигательные синергии [1–3]. Н.А. Бернштейн выделял понятие “обширные мышечные синергии” – способность выполнять высокослаженные движения всего тела, вовлекающие в согласованную работу многие десятки мышц. Параметры координационной структуры двигательных действий, реализуемых по такому принципу, имеют признаки согласованного управления на разных уровнях моторной системы. На кинематическом уровне такими признаками могут быть: сочетанное изменение суставных углов, градиента нарастания или снижения скорости движения отдельных сегментов тела, на мышечном – сопоставимое изменение параметров электроактивности ряда скелетных мышц, а на нейрональном уровне – изменение паттернов разрядов мотонейронов отдельных двигательных единиц [4, 5].
Для извлечения синергий, как правило, применяют методы снижения размерности данных, такие как метод главных компонент, неотрицательное матричное разложение и другие [6, 7]. Эти методы позволяют установить пространственную архитектуру мышечной активации, а также ее временнóй паттерн. Кросскорреляционный анализ электромиограмм (ЭМГ), отведенных от множества скелетных мышц одновременно, позволяет установить степень синфазности ЭМГ-активности и, соответственно, дает основание для отнесения ряда скелетных мышц к функциональному образованию – синергии. Кросскорреляционный анализ успешно применялся для изучения процессов иррадиации возбуждения на уровне спинного мозга при длительном мышечном напряжении, а также в отдельных фазах двигательных действий, различающихся по преобладающему режиму мышечного сокращения [8–10].
Как правило, при изучении мышечных синергий объектом исследования становятся локомоции, при этом анализируют синергетическое взаимодействие мышц нижних конечностей, а при выполнении произвольных движений чаще всего исследуют ЭМГ-активность мышц верхних конечностей. В плане изучения обширных функциональных синергий особый интерес представляет широкий спектр произвольных спортивных движений, для выполнения которых требуется проявление больших мышечных усилий и высокая точность движений. Такие сведения могут способствовать лучшему пониманию структуры целенаправленных спортивных движений и могут быть использованы в процессе обучения и совершенствования сложных двигательных навыков, характерных для определенного вида спорта. Однако вопросу изучения обширных мышечных синергий при выполнении таких движений уделяется недостаточное внимание. В связи с этим целью данной работы явилось изучение пространственно-временнóй организации мышечных синергий в разных периодах выполнения сложнокоординационного спортивного движения, различающихся по величине мышечных усилий.
МЕТОДИКА
В исследовании участвовали 12 спортсменов высокой спортивной квалификации (в возрасте 19–26 лет), занимающихся борьбой самбо. Исследования проводили на базе научно-исследовательского института проблем спорта и оздоровительной физической культуры Великолукской государственной академии физической культуры и спорта. Испытуемые выполняли серию бросков “захватом ног”. Порядок действий при выполнении броска представлен на рис. 1. Каждый испытуемый выполнял по 10 бросков с интервалом 30 с между ними. Ставилась задача выполнить бросок эффективно и с максимальной скоростью. В анализ включали только эффективные броски, выполненные без существенных нарушений техники. Броски выполняли без сопротивления, каждый борец выполнял броски на сопернике своей весовой категории. В общей сложности были проанализированы параметры 120 реализуемых бросков.
Рис. 1.
Регистрируемые периоды броска “захватом ног” и образец электромиограмм скелетных мышц. Вертикальными пунктирными линиями показаны граничные моменты периодов. Мышцы по порядку сверху вниз: ТА L, GM L, RF L, BF L, ТА R, GM R, RF R, BF R, TRAPS L, TRAPS R, TRAPI L, TRAPI R, ES L, ES R, RA L, RA R.

Во время выполнения движений регистрировали ЭМГ 16 билатеральных поверхностных мышц туловища, верхних и нижних конечностей: передней большеберцовой (TA), икроножной (медиальная головка) (GM), прямой бедра (RF), двуглавой бедра (BF), верхних пучков трапециевидной (TRAPS), средних пучков трапециевидной (TRAPI), выпрямляющей позвоночник (на уровне T9 позвонков) (ES), прямой живота (верхняя часть) (RA). Названные мышцы были выбраны для исследования в связи с ограничениями количества активных каналов применяемого биомонитора, а также, основываясь на результатах предварительных исследований, в которых регистрировали ЭМГ ряда мышц, не рассматриваемых в данной работе. Для решения задач данного исследования выбирали те мышцы, которые при выполнении модельного движения демонстрировали наибольшую ЭМГ–активность. Применяли беспроводной 16-канальный биомонитор МЕ 6000 (“Mega Electronics Ltd”, Финляндия), частота дискретизации ЭМГ составляла 2000 Гц. Использовали биполярные одноразовые отводящие электроды с диаметром 0.9 см, расстояние между электродами составляло 2 мм.
Синхронно осуществлялась запись видеоряда движений при помощи системы 3D-видеоанализа (“Qualisys”, Швеция) с частотой дискретизации 100 кадров в секунду. На основании определения граничных моментов, полученные вариационные ряды делили на 2 периода. Первый период включал движения от начала перемещения конечной антропометрической точки правой ноги до постановки ее на опору, второй – от момента завершения первой фазы до конца разгибания сзади стоящей ноги в коленном суставе (рис. 1).
Для извлечения мышечных синергий использовали метод главных компонент (Principal Component Analysis). ЭМГ предварительно сглаживались со временем интеграции 0.002 с. Отбор главных компонент осуществляли по правилу Кайзера, значимыми считали компоненты, имеющие собственные значения не менее единицы и описывающие в совокупности не менее 70% вариаций ЭМГ-активности. Поскольку значимые компоненты отражают результат одновременной активации ряда скелетных мышц, что является характерной чертой мышечных синергий, полученные компоненты (далее “основные сигналы ЭМГ”) считали синергией. Полученные таким образом ЭМГ-сигналы усредняли по группе испытуемых, применяли отрицательное экспоненциальное сглаживание и экстраполировали по времени относительно точки отсчета [11–13]. Анализ авто- и кросскорреляционных функций ЭМГ применяли для установления состава извлеченных мышечных синергий.
При анализе автокорреляционных функций учитывали скорость их затухания, т.е. время от начала координат до первого пересечения функции с нулевой линией, характеризующее наличие или отсутствие в ЭМГ периодических процессов. В кросскорреляционных функциях анализировали максимальную величину функции, а также наличие или отсутствие смещения фаз во времени (значимым считали сдвиг более 5 мс), характеризующее степень их синфазности [8]. Признаками синергетического взаимодействия считали наличие средних и высоких максимальных значений коэффициентов кросскорреляционных функций – более 0.5.
Математико-статистическая обработка данных включала расчет среднего арифметического (M), ошибки среднего арифметического (m), медианы (Me). Рассчитывали коэффициенты вариации (V) для сгруппированных данных. В вариационные ряды включали только значения, ограниченные верхним и нижним квартилями. Диапазон V от 0–30% считали низким уровнем, 30–60% – средним, от 60% и выше – высоким. Для оценки достоверности различий при сравнении параметров в разных периодах использовали однофакторный дисперсионный анализ (ANOVA) с Post–Hoc анализом по критерию Newman–Keuls. Различия считали значимыми при p < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Прежде чем приступить к описанию пространственно-временнóй организации обширных мышечных синергий, необходимо выяснить особенности ЭМГ-активности изучаемых мышц в рассматриваемых периодах броска “захватом ног”. Установлено, что в первом периоде ЭМГ-активность мышц голени регистрировалась в диапазоне от 95.65 до 126.16 мкВ, мышц бедра – от 66.38 до 128.41 мкВ. Электроактивность мышц туловища находилась в пределах от 80.31 до 175.97 мкВ (табл. 1). Наибольшая амплитуда была отмечена в TRAPI левой и правой стороны. Следует отметить, что в большинстве случаев различия между показателями среднего арифметического и медианы оказались незначительными, лишь в трех мышцах (GM L; TRAPS L, R) разница в значениях составила более 50 мкВ, в остальных мышцах она не превышала 23 мкВ. Вариативность ЭМГ-активности мышц правой голени и мышц бедра левой ноги, TRAPI и RA правой стороны оценивалась как средняя, остальные мышцы демонстрировали высокую внутригрупповую вариативность средней амплитуды ЭМГ.
Таблица 1.
Средняя амплитуда ЭМГ скелетных мышц в разных периодах выполнения броска “захватом ног” (мкВ)
| Скелетные мышцы | Период 1 | Период 2 | ||||
|---|---|---|---|---|---|---|
| M ± m | Me | V (%) | M ± m | Me | V (%) | |
| TA L | 115.39 ± 13.35 | 110.02 | 60.1 | 191.01 ± 13.20* | 190.48 | 35.9 |
| GM L | 126.16 ± 30.48 | 58.94 | 123.2 | 197.13 ± 30.75 | 139.33 | 79.5 |
| RF L | 82.62 ± 5.86 | 85.56 | 36.8 | 138.26 ± 13.27* | 118.18 | 49.9 |
| BF L | 66.38 ± 5.59 | 60.84 | 43.7 | 165.47 ± 10.85* | 170.96 | 34.1 |
| TA R | 113.67 ± 11.42 | 109.16 | 52.2 | 333.41 ± 33.48* | 304.76 | 52.2 |
| GM R | 95.65 ± 8.90 | 94.99 | 48.4 | 427.66 ± 70.01* | 323.17 | 85.1 |
| RF R | 68.62 ± 11.37 | 46.07 | 86.1 | 410.69 ± 66.22* | 256.58 | 83.8 |
| BF R | 128.41 ± 15.39 | 118.64 | 62.3 | 417.79 ± 44.68* | 445.95 | 55.6 |
| TRAPS L | 160.87 ± 23.73 | 103.46 | 76.6 | 418.79 ± 56.24* | 268.01 | 69.8 |
| TRAPS R | 175.97 ± 30.40 | 100.96 | 89.8 | 587.43 ± 88.39* | 366.65 | 78.2 |
| TRAPI L | 102.14 ± 11.89 | 99.35 | 60.5 | 381.36 ± 25.60* | 377.78 | 34.9 |
| TRAPI R | 91.66 ± 7.93 | 81.77 | 45.0 | 451.37 ± 49.25* | 444.01 | 56.7 |
| ES L | 80.31 ± 9.34 | 75.40 | 60.4 | 431.16 ± 46.29* | 330.89 | 55.8 |
| ES R | 92.07 ± 11.17 | 81.01 | 63.0 | 529.86 ± 53.16* | 491.72 | 52.1 |
| RA L | 84.4 ± 13.25 | 74.74 | 81.6 | 576.58 ± 62.38* | 492.79 | 56.2 |
| RA R | 101.45 ± 10.56 | 98.09 | 54.1 | 505.17 ± 44.57* | 500.69 | 45.8 |
Примечание: TA – передняя большеберцовая, GM – икроножная, RF – прямая бедра, BF – двуглавая бедра, TRAPS – верхние пучки трапециевидной, TRAPI – средние пучки трапециевидной, ES – выпрямляющая позвоночник, RA – прямая живота. L – левая сторона, R – правая сторона. * – достоверность различий средней амплитуды ЭМГ скелетных мышц в разных периодах движения, p < 0.05.
Второй период выполнения двигательного действия характеризовался достоверно большими значениями средней амплитуды ЭМГ практически всех исследуемых скелетных мышц, за исключением GM левой ноги. Диапазон значений ЭМГ-активности мышц голени левой ноги составил от 138.26 до 197.13 мкВ, правой ноги – от 333.41 до 427.66 мкВ. Электрическая активность мышц туловища регистрировалась в пределах от 381.36 до 587.43 мкВ. Разница между М и Ме более 50 мкВ установлена в GM левой и правой ноги, TRAPI левой и правой стороны, RF правой ноги, RA и ES левой стороны. Вариативность ЭМГ-активности этих мышц оценивалась как высокая. Другие исследуемые мышцы в этом периоде демонстрировали среднюю внутригрупповую вариативность. Таким образом, ЭМГ-активность большинства исследуемых скелетных мышц во втором периоде выполнения модельного двигательного действия оказалась существенно выше, чем в начальном. Кроме того, в начальном периоде выполнения движения установлена более высокая вариативность средней амплитуды ЭМГ регистрируемых мышц. Поскольку различия в величинах средней амплитуды ЭМГ и ее вариативности оказались значительными, логично было предположить, что и организация мышечных синергий в разные периоды движения должна иметь свою специфику.
Исходя из цели работы, были проанализированы основные сигналы ЭМГ исследуемых скелетных мышц в разные периоды изучаемого движения. По результатам анализа главных компонент всего движения выявлено три синергии (рис. 2). Первая синергия характеризовалась постепенным возрастанием ЭМГ-активности мышц, входящих в ее структуру, затем регистрировалось постепенное ее снижение при переходе из первого периода во второй. Длительность активности первой синергии оказалась сопоставима с длительностью рассматриваемого периода и составила 636 мс. Во втором периоде двигательного действия аналогично наблюдалось возрастание ЭМГ-активности синергии 1 и постепенное снижение к окончанию активных действий, длительность электроактивности данного периода составила 564 мс. Таким образом, активность первой установленной синергии имела волнообразный характер, причем во втором периоде регистрировалось несколько меньшая величина ЭМГ-активности, чем в первом. Следует также отметить, что в каждом рассматриваемом периоде движения амплитуда пиков ЭМГ-активности синергии 1 была больше амплитуды пиков синергий 2 и 3 (рис. 2).
Рис. 2.
Основные сигналы ЭМГ, характеризующие активность мышечных синергий (А). а – синергия 1, б – синергия 2, в – синергия 3. По оси абсцисс – время (мс), по оси ординат – величина электроактивности (у. е.). Вертикальными пунктирными линиями показан граничный интервал периодов движения. Автокорреляционные функции скелетных мышц (Б). По оси абсцисс – время (мс), по оси ординат – максимальный коэффициент кросскорреляционной функции. а – GM R, б – TRAPS L, в – TRAPI L, г – ES L.
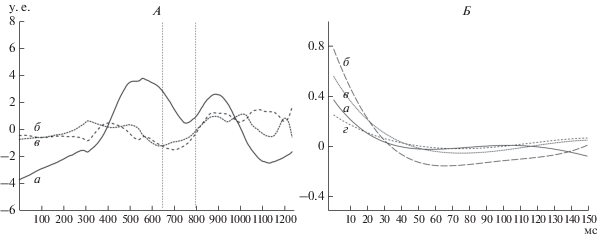
Мышечная синергия 2 характеризовалась возрастанием активности, и последующем снижении до исходного уровня, во временнóм отрезке от 300 до 500 мс. При переходе ко второму периоду движения активность мышц, входящих в синергию, постепенно нарастала и далее находилась на более высоком уровне, чем в первом периоде. Активность третьей синергии 3 демонстрировала приблизительно схожую динамику со второй синергией. Можно было определить, какие мышцы являются частью выявленных синергий в каждом рассматриваемом периоде модельного двигательного действия. Анализ параметров автокорреляционных функций позволил отнести исследуемые скелетные мышцы к определенным группам – синергиям.
Установлено, что время до первого пересечения автокорреляционной функции с нулевой линией TA, GM и RF левой стороны в начальном периоде движения составило 101–113 мс, в периоде окончания движения – 26–50 мс. В обоих изучаемых периодах названные мышцы имели схожие параметры автокорреляционных функций. Наряду с названными мышцами можно отметить наличие такой же закономерности в ЭМГ RA обеих сторон тела. Анализируя автокорреляционные функции ЭМГ других мышц в первом периоде можно выделить схожие ее параметры следующих: GM правой, TRAPI и TRAPS левой, ES левой. Во втором периоде установлены характерные паттерны ЭМГ следующих мышц: TA правой, TRAPI левой и правой стороны, TRAPS правой и ES левой стороны. Поскольку автокорреляционные функции отражают лишь наличие или отсутствие в сигналах ЭМГ регулярной составляющей, то она позволяет установить мышцы, имеющие схожие паттерны ЭМГ с регулярными колебаниями активности, либо нерегулярными, однако такой анализ не позволяет установить временнýю организацию мышечных синергий.
Для изучения временнóй структуры обширных мышечных синергий анализировали кросскорреляционные функции каждой пары скелетных мышц, вне зависимости от типа их взаимодействия. В целом проанализированы параметры 240 кросскорреляционных функций в двух периодах выполнения броска “захватом ног”. Установлено большее количество средних и высоких взаимосвязей в начальном периоде движения при отсутствии смещения кросскорреляционных функций. В начальном периоде статистически значимые взаимосвязи ЭМГ-активности обнаружены у ряда скелетных мышц, например, TA правой, BF правой, TRAPI и TRAPS левой стороны. Их кросскорреляционные функции представлены на рис. 3, А–В. Также установлены значимые связи RA правой стороны с RF левой, TA правой, с TRAPS левой; выпрямляющей позвоночник левой и прямой мышцы живота левой стороны тела. Были выявлены взаимосвязи и других скелетных мышц в данном периоде, однако они оказались не такими обширными, например, между передней большеберцовой, прямой бедра левой ноги и нижними пучками трапециевидной мышцы правой стороны тела. Наибольшее количество значимых взаимосвязей с другими исследуемыми мышцами в начальном периоде движения имели нижние пучки трапециевидной мышцы и прямая мышца живота правой стороны.
Рис. 3.
Кросскорреляционные функции некоторых исследуемых скелетных мышц и их ЭМГ в разных периодах выполнения броска “захватом ног”. Начальный период: А – TA L (а) и TRAPI L (б), Б – BF R (а) и TRAPI L (б), В – TRAPS L (а) и TRAPI L (б). Период завершения: Г – TRAPS L (а) и ES L (б), Д – TRAPS R (а) и ES L (б), Е – TRAPI R (а) и ES L (б).
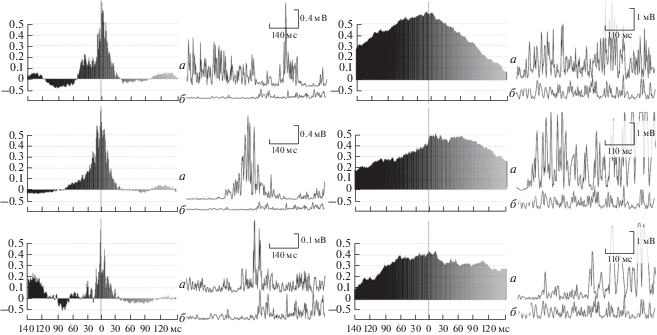
Во втором изучаемом периоде движения, где проявление мышечных усилий оказалось значительно выше, чем в первом, проанализированы параметры кросскорреляционных функций. Отличительной особенностью явилось то, что здесь количество средних и высоких коэффициентов, при отсутствии смещения функций, оказалось значительно меньше, чем в предыдущем периоде. Так, можно отметить взаимосвязи ЭМГ-активности ES левой стороны с ЭМГ-активностью TRAPI обеих сторон тела и TRAPS правой стороны (рис. 3, Г–Е). Также обнаружены взаимосвязи ЭМГ-активности прямой мышцы живота левой стороны с ЭМГ-активностью верхних пучков трапециевидной левой и мышцы, выпрямляющей позвоночник левой стороны.
Анализ кросскорреляционных функций, имеющих смещение в сторону отрицательных значений, в изучаемых периодах движения выявил следующие закономерности. В начальном периоде большее число взаимосвязей обнаружено с ES правой стороны. С ней оказались взаимосвязаны мышцы бедра левой ноги и GM правой ноги. С прямой мышцей живота правой стороны тела обнаружились взаимосвязи TA левой, TRAPS правой и ES левой стороны. В периоде проявления больших мышечных усилий такие синергетические взаимосвязи установлены между ES левой стороны, и мышцами бедра правой, а также BF левой нижней конечности.
При наличии смещения максимума кросскорреляционных функций в положительную сторону в первом периоде установлены обширные взаимосвязи BF правой ноги с TRAPS правой стороны, а также с мышцами туловища (табл. 2). Особенностью взаимосвязей мышечной активности во втором периоде явилось значительно меньшее их количество, чем в периоде начала выполнения двигательного действия. Можно отметить электроактивность прямой мышцы живота левой стороны и характерные паттерны ЭМГ-активности мышц бедра правой нижней конечности и мышцы, выпрямляющей позвоночник левой стороны. Максимальные величины их коэффициентов кросскорреляционных функций здесь оказались средними.
Таблица 2.
Максимальные величины кросскорреляционных функций некоторых скелетных мышц в разные периоды выполнения броска “захватом ног”
| Период | Мышцы | TA R | GM R | RF R | BF R | TRAPS L | TRAPS R | TRAPI L | TRAPI R | ES L |
|---|---|---|---|---|---|---|---|---|---|---|
| Период 1 | BF R | 0.63 | 0.58 | 0.40 | ||||||
| TRAPS L | 0.49 | 0.47 | 0.46 | 0.43 | ||||||
| TRAPS R | 0.21 | 0.28 | 0.52 | 0.17 | 0.30 | |||||
| TRAPI L | 0.70 | 0.66 | 0.50 | 0.73 | 0.63 | 0.30 | ||||
| TRAPI R | 0.01 | 0.28 | 0.52 | 0.52 | 0.30 | 0.57 | 0.43 | |||
| ES L | 0.01 | –0.18 | –0.10 | 0.52 | 0.32 | 0.25 | 0.53 | 0.31 | ||
| ES R | 0.63 | 0.69 | 0.49 | 0.56 | 0.49 | 0.42 | 0.60 | 0.47 | 0.42 | |
| RA L | 0.50 | 0.46 | 0.42 | 0.66 | 0.42 | 0.33 | 0.64 | 0.44 | 0.43 | |
| RA R | 0.62 | 0.57 | 0.54 | 0.73 | 0.40 | 0.25 | 0.67 | 0.64 | 0.51 | |
| Период 2 | BF R | 0.37 | 0.01 | 0.53 | ||||||
| TRAPS L | 0.50 | 0.01 | 0.47 | 0.45 | ||||||
| TRAPS R | 0.47 | –0.10 | 0.44 | 0.32 | 0.63 | |||||
| TRAPI L | 0.30 | 0.16 | 0.46 | 0.43 | 0.36 | 0.27 | ||||
| TRAPI R | 0.40 | 0.01 | 0.43 | 0.39 | 0.48 | 0.57 | 0.39 | |||
| ES L | 0.53 | 0.01 | 0.61 | 0.52 | 0.73 | 0.60 | 0.44 | 0.51 | ||
| ES R | –0.12 | 0.24 | 0.42 | 0.31 | 0.01 | 0.14 | 0.30 | 0.27 | 0.24 | |
| RA L | 0.45 | –0.20 | 0.52 | 0.55 | 0.70 | 0.45 | 0.45 | 0.43 | 0.65 | |
| RA R | 0.21 | 0.19 | 0.45 | 0.46 | 0.17 | 0.15 | 0.50 | 0.37 | 0.31 |
Примечание: TA – передняя большеберцовая, GM – икроножная, RF – прямая бедра, BF – двуглавая бедра, TRAPS – верхние пучки трапециевидной, TRAPI – средние пучки трапециевидной, ES – выпрямляющая позвоночник, RA – прямая живота. L – левая сторона, R – правая сторона. Полужирным шрифтом выделены средние и высокие коэффициенты кросскорреляционных функций, имеющие смещение в сторону положительных значений.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Результаты исследований показали значительные различия в величинах электроактивности скелетных мышц при выполнении броска “захватом ног” в разных периодах его реализации. Средняя амплитуда ЭМГ большинства скелетных мышц в завершающем периоде движения оказалась значительно выше, чем в начальном, причем более высокие показатели ЭМГ-активности характеризовались меньшей вариативностью. Можно считать, что наблюдаемая вариативность электроактивности скелетных мышц в начальном периоде движения более значительная, поскольку известно, что в движениях или его отдельных фазах, имеющих различную целевую значимость, используются разные механизмы управления на разных уровнях ЦНС [1, 12, 13]. Вероятно, завершающий период изучаемого движения имеет результирующее значение и поэтому диапазон допустимой вариативности ЭМГ-активности скелетных мышц, задействованных в движении, оказался ниже, чем в начальном.
В настоящем исследовании установлены три мышечные синергии, активность которых имела характерные особенности, проявляющиеся в возрастании электроактивности основных сигналов ЭМГ в начале движения и ее снижении на границе перехода между регистрируемыми периодами движения. В данном случае по относительной величине электроактивности синергии можно выяснить, какие мышцы являются ее составляющими. Например, по результатам анализа главных компонент установлено, что в одну из мышечных синергий входят мышцы, имеющие наибольшие величины средней амплитуды ЭМГ. В начальном периоде движения таковыми являлись мышцы голени левой нижней конечности, двуглавая мышца бедра, верхние и нижние пучки трапециевидных мышц левой и правой стороны. Такое же количество синергий было выявлено J. Frere и F. Hug при исследовании некоторых специализированных гимнастических, сложных по координации движений [14]. Аналогичное количество синергий было извлечено из ЭМГ-мышц, задействованных при выполнении жима лежа спортсменами высокой квалификации [15]. Однако в работах, где в качестве объекта исследования выступали локомоторные движения, регистрировалось от 4 до 5 мышечных синергий [6, 16]. Таким образом, для эффективного выполнения двигательных задач, связанных с проявлением значительных мышечных усилий и высокой координационной точности требуется более значительное ограничение степеней свободы мышечного аппарата, чем это необходимо при локомоторной активности. Возможно, такие особенности организации мышечных синергий являются механизмом, уменьшающим возможность получения травмы, вероятность которой, при выполнении сложных спортивных движений, достаточно велика.
В начальном периоде изучаемого движения установлена большая вариативность состава мышечных синергий, проявляющаяся в обширных взаимосвязях ЭМГ-активности мышц туловища с мышцами бедра и голени, причем такие взаимосвязи обнаруживаются и в билатеральных мышцах. В завершающем периоде броска “захватом ног” состав синергий оказался менее вариативен, чем в начальном периоде. Взаимосвязи ЭМГ-активности здесь в основном были сконцентрированы на мышцах туловища. Полученные нами данные в некоторых аспектах согласуются с результатами работs S. Shaharudin [17]. Авторами показана гибкость состава мышечных синергий при выполнении гребковых движений на различных тренажерах. Количество извлеченных синергий из ЭМГ-мышц в таких условиях оказалось одинаковое – 3, однако авторы наблюдали разную вариативность состава мышц каждой установленной синергии, что объясняется стратегией ЦНС, направленной на эффективное выполнение сложнокоординационного движения. Аналогичное количество синергий было установлено и в исследованиях, где объектом изучения являлся сложный гимнастический элемент – большие обороты на перекладине [14]. В этой работе показана вариабельность одной из трех установленных мышечных синергий, при этом другие две оказались схожи у разных спортсменов, что является подтверждением существования единой нервно-мышечной стратегии управления сложным специализированным спортивным движением. Три синергии было выявлено группой авторов при исследовании силового жима лежа, выполняемого спортсменами разной спортивной квалификации [15]. В исследовании не было установлено отличий в количестве мышечных синергий в различных фазах движения, различающихся по преобладающему режиму мышечных сокращений. Однако была установлена большая вариативность состава мышечных синергий у группы высококвалифицированных пауэрлифтеров в фазе концентрического сокращения, по сравнению c менее квалифицированными.
Анализ зависимостей ЭМГ, регистрируемых в нашем исследовании мышц, показал наличие максимальных коэффициентов кросскорреляционных функций при их смещении, как в положительную, так и в отрицательную сторону. Это свидетельствует о том, что синергетические взаимоотношения скелетных мышц имеют временнýю организацию, а именно, активность одной мышцы в структуре синергии может запаздывать или предшествовать активности другой. При этом ЭМГ-активность мышц сохраняет характерные черты, определяющие их отношение к синергии. Известно, что синергии организованы на корковом, подкорковом и спинальном уровнях, каждый из которых вносит свой вклад в их структуру, однако ведущая роль в управлении мышечными синергиями принадлежит структурам мозжечка [18]. Во время двигательной активности головной мозг корректирует работу спинальных механизмов на основании сигналов, поступающих от рецепторов исполнительного двигательного аппарата, в головной мозг также поступают сигналы о работе центральных генераторов [19]. Выходные сигналы таких генераторов являются программами синергий, определяющие их структурную и временнýю организацию [2, 20]. Такие параметры моторного выхода являются “кодом” моторной программы, управляющей активацией мотонейронных пулов различных сегментов спинного мозга, осуществляющих управление активностью скелетных мышц [2, 21]. Моторная программа в этом случае определяет последовательность и интенсивность управляющих сигналов в ЦНС для успешного решения реализуемой двигательной задачи.
Была установлена закономерность, заключающаяся в меньшем количестве активных мышечных синергий в периоде двигательного действия, где регистрировались более значительные мышечные напряжения (завершающий период). Такая организация мышечных синергий может осуществляться с использованием механизмов регуляции силы мышечных сокращений ЦНС, в том числе механизма синхронизации активности отдельных двигательных единиц (ДЕ) во времени. Суть такого управления заключается в суммации сокращений отдельных ДЕ, за счет чего увеличивается сила мышечного сокращения. Синхронизация активности ДЕ влияет на скорость сокращения и имеет важное значение в начальном периоде любого движения. Как правило, при выполнении быстрых движений синхронизация активности ДЕ более выражена в его начале, чем в завершающем периоде. Это обусловлено необходимостью преодолевать значительную внешнюю нагрузку в начале двигательного действия. Высокая частота импульсации и активность большого количества мотонейронов повышает вероятность совпадения сократительных циклов различных ДЕ в начале движения [22]. Таким образом, образование большего количества активных мышечных синергий в начальном периоде сложного координационного двигательного действия, вероятно, обусловлено необходимостью совершать быстрые и точные движения, направленные на преодоление значительной нагрузки.
ЗАКЛЮЧЕНИЕ
Таким образом, установленные закономерности вариативности электроактивности скелетных мышц, задействованных в реализации сложного координационного двигательного действия, в различных периодах его выполнения связаны с их целевой значимостью. Электроактивность обширных синергий меняется в зависимости от величины проявляемых мышечных усилий, характерных для разных периодов выполняемого движения. Синергетические взаимоотношения скелетных мышц демонстрируют пластичность их пространственной и временнóй организации. Большее количество активных мышечных синергий в начальном периоде броска “захватом ног”, вероятно, обусловлено необходимостью совершать быстрые и точные движения, направленные на преодоление значительной нагрузки, что обеспечивает надежное управление двигательной функцией. При изучении обширных мышечных синергий целесообразно использовать комплекс методик (метод главных компонент, анализ авто- и кросскорреляционных функций), позволяющий установить их пространственную и временнýю организацию.
Этические нормы. Все эксперименты проведены с соблюдением требований и принципов биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и одобрены биоэтическим комитетом Великолукской государственной академии физической культуры и спорта (Великие Луки).
Информированное согласие. Каждый участник представил добровольное письменное информированное согласие на участие в исследованиях, подписанное им после разъяснения ему потенциальных рисков, а также характера предстоящего исследования.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Бернштейн Н.А. Очерки по физиологии движений и физиологии активности. М.: Медицина, 1966. 349 с.
Latash M., Zatsiorsky V. Biomechanics and Motor Control. USA: Academic Press is an imprint of Elsevier, 2016. 409 p.
Николлс Д., Мартин Р., Валлас Б. и др. От нейрона к мозгу. М.: ЛКИ, 2008. 672 с.
Персон Р.С. Спинальные механизмы управления мышечным сокращением. М.: Наука, 1985. 184 с.
Rathelot J., Strick P. Muscle representation in the macaque motor cortex: an anatomical perspective // Proc. Natl. Acad. Sci. USA. 2006. V. 103. № 21. P. 8257.
Ivanenko Y.P, Cappellini G., Dominici N. et al. Coordination of locomotion with voluntary movements in humans // J. Neurosci. 2005. V. 25. № 31. P. 7238.
Tresch M.C., Cheung V.C., D’Avella A. Matrix factorization algorithms for the identification of muscle synergies: evaluation on simulated and experimental data sets // J. Neurosci. 2006. V. 95. № 4. P. 2199.
Персон Р.С. Электромиография в исследованиях человека. М.: Наука, 1969. 227 с.
Шелякин А.М., Преображенская И.Г., Богданов О.В. Кросскорреляционный анализ биоэлектрической активности мышц–антагонистов в исследовании произвольной двигательной активности человека при некоторых формах двигательных нарушений // Рос. физиол. журн. 1997. Т. 83. № 9. С. 88.
Tishya A.L. Wren, K. Patrick Do, Susan A. Rethlefsen et al. Cross-correlation as a method for comparing dynamic electromyography signals during gait // J. Biomech. 2006. V. 39. № 14. P. 2714.
Latash M.L., Scholz J.P., Schoner G. Motor control strategies revealed in the structure of motor variability // Exerc. Sport Sci. Rev. 2002. V. 30. № 1. P. 26.
Латаш М.Л. Структурированная вариабельность как отличительный признак биологических процессов // Вопросы психологии. 2016. № 3. С. 120.
Моисеев С.А., Пухов А.М. Роль функциональных синергий в управлении пространственно-временнóй структурой точностных движений человека (на примере стрельбы из лука) // Журн. медико-биологических исследований. 2019. Т. 7. № 4. С. 410.
Frère J., Hug F. Between-subject variability of muscle synergies during a complex motor skill // Front. Comput. Neurosci. 2012. V. 6. P. 99.
Kristiansen M., Samani A., Madelein P. et al. Effects of 5 weeks of bench press training on muscle synergies: a randomized controlled study // J. Strength Cond. Res. 2016. V. 30. № 7. P. 1948.
Kibushi B., Hagio S., Moritani T., Kouzaki M. Speed-Dependent Modulation of Muscle Activity Based on Muscle Synergies during Treadmill Walking // Front. Hum. Neurosci. V. 12. № 4. P. 4.
Shaharudin S., Zanotto D., Agrawal S. Muscle Synergies of Untrained Subjects during 6 min maximal rowing on slides and fixed ergometer // J. Sports Sci. Med. 2014. V. 13. № 4. P. 793.
McMorland A.J., Runnalls K.D., Byblow W.D. A neuroanatomical framework for upper limb synergies after stroke // Front. Hum. Neurosci. 2015. V. 9. P. 82.
Orlovsky G.N., Deliagina T.G., Grillner S. Neuronal Control of Locomotion. UK: Oxford Univ. Press, 1999. P. 336.
Gerasimenko Y., Sayenko D., Gad P. et al. Electrical spinal stimulation, and imagining of lower limb movements to modulate brain-spinal connectomes that control locomotor-like behavior // Front. Physiol. 2018. V. 9. P. 1196.
Bizzi E., Cheung V.C. The neural origin of muscle synergies // Front. Comput. Neurosci. 2013. V. 7. № 51. P. 51.
Гурфинкель В.С., Левик Ю.С. Скелетная мышца: структура и функция. М.: Наука, 1985. 143 с.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека