

Физиология человека, 2021, T. 47, № 6, стр. 5-17
Роль скрытых звеньев в работе мозговых систем обеспечения идентификации и категоризации зрительных стимулов
М. В. Киреев 1, 2, *, Р. С. Машарипов 1, А. Д. Коротков 1, С. В. Медведев 1
1 ФБГУН Институт мозга человека имени Н.П. Бехтеревой РАН
Санкт-Петербург, Россия
2 ФГБОУ ВО Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
* E-mail: kireev@ihb.spb.ru
Поступила в редакцию 10.12.2020
После доработки 07.04.2021
Принята к публикации 26.05.2021
Аннотация
Одним из пока неразрешенных вопросов в области изучения системной активности мозга человека является противоречивость данных о локальной активности отдельных звеньев мозговых систем и их дистантных взаимодействий. В рамках развития этого направления исследований недавно был обнаружен феномен скрытых звеньев – целый ряд структур мозга мог вовлекаться в их работу без выявляемого изменения энергопотребления, только за счет изменения характера дистантных функциональных связей. В данной работе продолжили изучение феномена скрытых звеньев на материале фМРТ-данных, полученных в условиях идентификации и категоризации изображений. В статье был предложен способ определения местонахождения скрытых звеньев, а также с привлечением методов теории графов получено подтверждение о ключевом характере участия “скрытых звеньев” в системной активности мозга. Помимо этого, показана возможность существования разных типов скрытых звеньев, выполняющих разные функции в конфигурации системной активности мозга.
Изучение системной организации мозга человека издавна является одним из актуальнейших направлений психофизиологии. Сейчас уже мало кто сомневается в том, что поведение человека, его психические функции обеспечиваются распределенными корково-подкорковыми мозговыми системами. В современных исследованиях наряду с традиционными активационными исследованиями таких систем лидирующие позиции занимают исследования функциональных взаимодействий. При этом представления о закономерностях реорганизации межзвеньевых взаимодействий в процессе обеспечения высших видов нервной деятельности человека находятся в стадии формирования [1–4]. Одним из пока неразрешенных вопросов этого направления исследований функциональной организации мозга человека является противоречивость данных о локальной активности отдельных звеньев мозговых систем и их дистантных взаимодействиях [3, 5–7]. Зачастую в рамках обеспечения внешне однородной деятельности, например, речевой, могут наблюдаться несоответствия между направлениями изменений локального энергопотребления и дистантных взаимодействий [7, 8]. Внимательное изучение данного эффекта позволило обнаружить новый феномен системной активности мозга – феномен скрытых звеньев [3, 8]. На основании наблюдаемого феномена была сформулирована гипотеза о новом свойстве работы мозговых систем, при котором целый ряд структур мозга мог вовлекаться в их работу без выявляемого изменения энергопотребления, только за счет изменения характера дистантных функциональных связей. Из этого следует, что звено может считаться “скрытым”, только если продемонстрировано, во-первых, значимое практическое отсутствие (practical equivalence testing) изменений энергопотребления, а во-вторых, включение такого звена в функциональные взаимодействия. Таким образом, для выявления “скрытых звеньев” необходимо применение специализированных методов, учитывающих характер их вовлечения – отсутствие проявления их работы в рамках активационных исследований. Поэтому до недавнего времени отсутствие статистических методов по анализу групповых данных фМРТ-исследований, позволяющих признать статистически значимой нулевую гипотезу о равенстве средних, являлось препятствием в изучении “скрытых звеньев”. Так, при использовании классической частотной статистики (тестирование значимости нулевой гипотезы, null hypothesis significance testing – NHST) отсутствие статистически значимых различий не может быть содержательно интерпретировано, т.е. не свидетельствует об отсутствии различий между переменными. В последнее время стали появляться новые подходы, преодолевающие данное ограничение. В рамках апробации одного из таких подходов, основанного на Байесовской статистике и позволяющего принять нулевую гипотезу об отсутствии практических различий между сравниваемыми групповыми переменными при анализе фМРТ-данных, были получены предварительные данные, свидетельствующие в пользу принципиальной возможности в выборе областей мозга, которые являются кандидатами на роль скрытых звеньев [9]. Это открывает и новые экспериментальные возможности в изучении функциональной роли “скрытых звеньев”.
В соответствии с выдвинутой ранее гипотезой [3] скрытыми звеньями могут являться такие структуры мозга, которые проявляют свое участие в обеспечении текущей деятельности, меняя характер своего участия в функциональных взаимодействиях с другими звеньями мозговых систем. При этом конкретная функциональная роль скрытых звеньев в работе мозговых систем является неисследованной. Исходя из этого, в рамках настоящей работы было предположено, что, по крайней мере, некоторые из скрытых звеньев могут иметь важное значение в организации нейросетевой активности, являясь так называемыми хабами. В данном случае под хабом понимается структура мозга, которая играет связующую роль между взаимодействующими звеньями на внутри- и меж-системном уровне взаимодействия [4, 6]. Если при обеспечении какой-либо деятельности скрытое звено выступает в роли хаба, то соответствующим образом должны меняться и конфигурационные свойства дистантных взаимодействий такого звена [10].
Для экспериментальной проверки гипотезы настоящего исследования использовали экспериментальный материал, полученный в условиях категоризации изображений, который подразумевал как определение одушевленности/неодушевленности объекта, так и идентификацию категории объекта (реально существующий или нет), предъявляемых на изображениях. Использование именно таких тестовых заданий было продиктовано тем, что распознавание, идентификация и категоризация объектов окружающего мира являются одними из ключевых процессов высшей нервной деятельности, а мозговые системы их обеспечения относительно подробно исследованы. Считается, что ключевыми звеньями мозговых систем обеспечения идентификации и категоризации объектов разных сенсорных модальностей являются структуры вентрального пути, включая нижнюю височную кору и вентролатеральную префронтальную кору [11, 12]. Однако феноменология “скрытых звеньев” предполагает, что скрытым звеном может оказаться структура мозга, которая ранее не проявляла и не проявляет своего участия в обеспечении исследуемой деятельности изменением своей локальной активности. А с учетом того, что представления о многих мозговых системах основываются на данных активационных исследований, авторы настоящей статьи предполагали обнаружить новые звенья, о которых не сообщалось в предыдущих исследованиях.
МЕТОДИКА
В двух фМРТ-исследованиях анализировали результаты, полученные у двух независимых групп здоровых праворуких добровольцев. Первая группа состояла из 41 добровольца (17 мужчин, средний возраст ± стандартное отклонение = 23.9 ± 2.3 лет). Вторая группа состояла из 38 добровольцев (18 мужчин, 24.8 ± 4.6 лет). Праворукость оценивали по вопроснику Олдфилда [13].
Тестовые задания. В первом исследовании (рис. 1, А) добровольцам предъявляли: 1) неискаженные изображения реальных объектов (“normal”, Norm); 2) частично искаженные изображения, на которых возможно идентифицировать реальные объекты (“scrambled ”, Scr); 3) полностью искаженные изображения, на которых невозможно идентифицировать реальные объекты (“totally scrambled”, TotScr). Первые два типа стимулов могли включать как одушевленные, так и неодушевленные объекты. Согласно инструкции добровольцы старались как можно быстрее распознать реальный объект на изображении. Во втором исследовании (рис. 2, Б) добровольцам предъявлялись: 1) неискаженные изображения одушевленных объектов (“animate”, Anim); 2) неискаженные изображения неодушевленных объектов (“inanimate”, Inanim); 3) полностью искаженные изображения, на которых невозможно идентифицировать реальные объекты (TotScr). Согласно инструкции, добровольцы старались как можно быстрее определить, относятся ли объекты неискаженных изображений к одушевленной или неодушевленной категории. В случае искаженных изображений добровольцам необходимо было сделать предположение о том, какой объект (одушевленный или неодушевленный) присутствовал на изображении до его искажения. В обоих заданиях отождествление объектов с той или иной категорией производили нажатием кнопок джойстика большим и указательным пальцем правой руки.
Каждое задание состояло из 102 зрительных стимулов (по 34 стимула каждого типа) длительностью 700 мс. Межстимульный интервал варьировался от 3100 до 3500 мс с шагом 100 мс. В отсутствии стимуляции в центре экрана появлялся крест для фиксации взгляда. Для увеличения статистической эффективности дизайна между экспериментальными пробами случайным образом предъявляли так называемые пробы “пустышки” [14] длительностью 4000 мс, неотличимые от межстимульного интервала (изображение креста для фиксации взгляда). Их количество составило 51 (1/3 от общего числа проб). Одно тестовое задание длилось 11 мин.
В качестве стимулов использовали эмоционально нейтральные изображения стандартизированной базы “International affective picture system, IAPS” [15]. Искажение (scrambling) изображений производили при помощи метода диффеоморфных трансформаций, сохраняющего базовые перцептивные свойства изображений (цвета, пространственная частота и т.д.) [16].
Регистрация и предварительная обработка фМРТ-изображений. Исследование проводили на томографе Philips Achieva 3.0 Тесла. Структурные T1-изображения регистрировали с параметрами: поле обзора (field of view, FOV) – 240 × 240 мм; repetition time (TR) – 25 мс; echo time (TE) – 2.2 мс; 130 аксиальных срезов толщиной 1 мм и размером пикселя 1 × 1 мм; угол отклонения намагниченности (flip angle) – 30°. Для регистрации функциональных T2*-изображений использовали эхопланарную одноимпульсную последовательность. Время регистрации данных с 32 аксиальных срезов составляло 2 с (TR = 2 c, TE = 35 мс). FOV – 200 × 186 мм, flip angle – 90°. Размер воксела 3 × × 3 × 3 мм. Предварительная обработка изображений включала: пространственное выравнивание изображений, коррекцию на время регистрации срезов одного изображения, корегистрацию структурных и функциональных изображений, сегментацию структурных изображений на разные виды тканей, нормализацию к стандартному координатному пространству Монреальского Неврологического Института (MNI, Monreal Neurological Institute, Канада) и пространственное сглаживание (Гауссовский фильтр с шириной 8 мм). Обработку изображений и статистический анализ проводили в программном пакете SPM12 (Statistical parametric mapping) (http://www.fil.ion.ucl.ac.uk/spm).
Статистический анализ. Внутрисубъектный или первый уровень анализа локальной нейрональной активности по данным фМРТ проводили с помощью классической частотной статистики (метод максимального правдоподобия, maximum likelihood estimation – MLE). Межсубъектный или второй уровень анализа проводили с помощью Байесовской статистики (метод оценки апостериорного максимума, maximum a posteriori estimation – MAP) [17–19]. На первом этапе для каждого добровольца определялась и активность мозга, связанную с выполнением тестовых заданий. В общую линейную модель (ОЛМ) было включено три регрессора для интересующих экспериментальных условий. Ошибки при выполнении тестового задания указывали в отдельном регрессоре. Регрессоры свертывались с канонической гемодинамической функцией. Для учета влияния артефактов движения использовали 24 регрессора согласно K.J. Friston et al. [20]. Параметры регрессоров ОЛМ были переведены в проценты изменения BOLD-сигнала (Blood-oxygen-level-dependent signal) относительно средних значений сигнала во всем объеме мозга на базовом уровне [21]. На первом уровне анализа для первого варианта тестового задания рассчитывали следующие линейные контрасты для определения разницы BOLD-сигнала в интересующих экспериментальных условиях: 1) “0.5 × Norm + 0.5 × × Scr” – “TotScr”; 2) “Scr” – “Norm”. Для второго варианта тестового задания: 3) “0.5 × Anim + 0.5 × × Inanim” – “TotScr”; 4) “Anim” – “Inanim”. Данные контрасты использовали в качестве переменных на втором уровне анализа. Первый и третий контраст позволил определить области мозга, в которых наблюдается повышение сигнала при восприятии распознаваемых изображений по сравнению с нераспознаваемыми изображениями. Второй и четвертый контраст при помощи Байесовской статистики позволил определить области мозга, в которых наблюдается значимо не различающийся уровень локальной нейрональной активности при восприятии разных категорий распознаваемых изображений. Пересечение результатов (conjunction analysis) перечисленных контрастов позволило определить возможных кандидатов на роль “скрытых звеньев”. То есть определить области, в которых, с одной стороны, наблюдается повышение локальной активности мозга при распознании объектов в целом, но, с другой стороны, не наблюдается значимых различий при распознании разных категорий объектов. При дальнейшем анализе, в случае, если для этих областей мозга будет обнаружено значимое изменение силы дистантных взаимодействий при распознавании разных категорий объектов, их можно будет отнести к “скрытым звеньям”.
Для определения наличия значимых различий (активаций) в первом и третьем контрасте (“распознаваемые vs. нераспознаваемые изображения”) использовали классический частотный вывод (NHST) с порогом статистической значимости p < < 0.05 с коррекцией на множественность сравнений по методу контроля групповой вероятности ошибки первого рода (Family-wise error correction, FWE-correction). Для определения наличия или отсутствия практически значимых различий во втором и четвертом контрасте (“распознаваемые изображения разных категорий”) использовали Байесовский вывод, основанный на расчете апостериорной вероятности обнаружения эффекта определенной величины. Если апостериорная вероятность обнаружения практически значимого эффекта в определенном контрасте превышает порог вероятности 95%, то принимается гипотеза о наличии эффекта. Практически значимый эффект определяется с помощью порога для размера эффекта γ. Если эффект попадает в интервал [–γ; γ] (область практической эквивалентности, region of practical equivalence, ROPE) с вероятностью более 95%, то принимается нулевая гипотеза об отсутствии эффекта [9, 22], что позволяет говорить о практической эквивалентности локальной нейрональной активности в сравниваемых пробах. Иными словами, это дает возможность считать нулевую гипотезу о равенстве средних значимой. В настоящей работе пороги для практически значимого размера эффекта составили: 1) γ = 0.043% (контраст “Scr” – “Norm”), 2) γ = 0.058% (контраст “Anim” – “Inanim”). При таком размере эффекта обеспечивалось максимальное соответствие Байесовского вывода и классического вывода с FWE-коррекцией для выявления значимых различий. Выявление аналогичного паттерна активаций с помощью данных порогов γ показывает адекватность контроля ложноположительных результатов и обосновывает их использование для определения областей практической эквивалентности (ROPE). Соответствие между паттернами активаций, выявляемых с помощью двух методов, определяли с помощью коэффициента Дайса [23]: 1) Dicemax = 0.93 (контраст “Scr” – “Norm”); 2) Dicemax = 0.92 (контраст “Anim” – “Inanim”). Для того, чтобы при групповом сравнении анализировались только те воксели, которые относились к серому веществу головного мозга, была создана маска на основе данных сегментации структурных T1-изображений каждого испытуемого.
Для проверки гипотезы исследования анализ по выявлению “скрытых звеньев”, являющихся ключевыми для организации нейросетевой активности мозга, проводили в два этапа. На первом этапе, для того, чтобы определить, являются ли области-кандидаты на роль “скрытых звеньев” важными хабами мозговых систем распознавания и категоризации изображений, проводили анализ функциональных связей разноудаленных структур мозга с использованием методов теории графов. Для построения симметричных матриц функциональных связей использовали метод корреляции серий β-коэффициентов (beta-series correlation, BSC) [24, 25]. Функциональные связи рассчитывали между функциональными областями интереса во всем объеме мозга [26] и областями-кандидатами на роль “скрытых звеньев”. Набор 300 областей из [26] был сокращен до 218 областей, входящих в маску серого вещества, полученную в настоящей работе, и не пересекающихся с областями-кандидатами в “скрытые звенья”. В результате, для двух контрастов типа “распознаваемые vs. нераспознаваемые изображения” (“0.5 × Norm + 0.5 × Scr” – “TotScr” и “0.5 × Anim + 0.5 × Inanim” – “TotScr”) были получены усредненные по группам матрицы функциональных связей. Положительные значения (взвешенные матрицы) были использованы для расчета графовых метрик с помощью программ GRETNA (http://www.nitrc.org/projects/gretna/) и Brain Connectivity Toolbox (https://sites.google.com/ site/bctnet/). Для определения конфигурационных характеристик связей использовали следующие метрики: 1) степень посредничества узла (Betweenness Сentrality); 2) степень узла (Nodal Degree); 3) коэффициент участия (Participation Coefficient); 4) локальную эффективность узла (Local Efficiency). Степень посредничества узла (1) зависит от количества кратчайших путей, проходящих через данный узел/звено: чем выше их количество, тем выше степень посредничества. Узел с высоким значением посредничества может рассматриваться в качестве “бутылочного горлышка” при передаче информации в системе взаимодействующих между собой звеньев. Степень узла (2) зависит от количества связей данного узла с остальными узлами графа. Узлы графов с высокой степенью облегчают передачу информации в системе и увеличивают ее функциональную интеграцию – чем больше звеньев с высокими значениями степени узла, тем более интегрированной является такая система. Коэффициент участия (3) характеризует участие узла в передаче информации между функциональным модулем (обособленными группами связанных между собой звеньев), к которому он принадлежит, и другими модулями системы. Локальная эффективность узла (4) обратно пропорционально зависит от длины кратчайших путей от соседей данного узла. Узлы с высоким значением локальной эффективности обеспечивают надежность передачи информации при выключении других звеньев системы. В настоящей работе хабами считались узлы, в которых значения графовых метрик входили в 25% максимальных значений распределения для двух тестовых заданий (по аналогии с [6]).
На втором этапе для областей-кандидатов в “скрытые звенья”, проявляющих также свойства хабов мозговых систем распознавания и категоризации изображений, проводили повоксельный анализ изменения дистантных функциональных взаимодействий. Наличие таких изменений является вторым критерием соответствия “скрытому звену”. При этом использовали обобщенный метод психофизиологических взаимодействий (generalized form of psychophysiological interactions, gPPI) [27]. В данном виде анализа общая линейная модель, описанная ранее, дополнительно включает в себя также физиологический регрессор и регрессор психофизиологического взаимодействия. Физиологический регрессор, Xphysio(t), представляет собой BOLD-сигнал в области интереса. Расчет регрессора психофизиологического взаимодействия, XPPI(t), включает в себя три этапа [28]. Сначала производится обратная свертка (⨂ – оператор свертки) функции физиологического регрессора с гемодинамической функцией, HRF(t), для того, чтобы оценить лежащую в основе BOLD-сигнала нейрональную активность Zphysio(t): Xphysio(t) = Zphysio(t) ⨂ HRF(t). Далее производится перемножение физиологического и психологического регрессора, Zpsy(t), принимающего значение 1 при наличии тестового условия и значение 0 при отсутствии тестового условия. Полученное произведение представляет собой психофизиологическое взаимодействие на нейрональном уровне. Для того чтобы смоделировать это взаимодействие на уровне BOLD-сигнала производится свертка этого произведения с функцией гемодинамического ответа: XPPI(t) = (Zphysio(t) × Zpsy(t)) ⨂ HRF(t). Регрессоры психологических взаимодействий создавали отдельно для каждого из интересующих авторов настоящей статьи тестовых условий. Данный метод позволяет отделить контекстно-зависимые изменения силы функциональных связей от коактиваций и корреляций спонтанных флуктуаций сигнала, наблюдаемых в состоянии оперативного покоя. Рассматривали контрасты, сравнивающие пробы с разными типами распознаваемых изображений: 1) “Scr” – “Norm”, 2) “Anim” – “Inanim”. Порог для определения статистически значимых различий функциональных взаимодействий составил p < 0.05 (с FWE-коррекцией на кластерном уровне).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Результаты классического частотного анализа и байесовского анализа локальной нейрональной активности. Установлены области, демонстрирующие больший уровень BOLD-сигнала в условиях с неискаженными и частично искаженными объектами, по сравнению с полностью искаженными изображениями, на которых невозможно идентифицировать реальный объект (рис. 2, A – для первого исследования, рис. 2, Б – для второго исследования, а также табл. 1).
Таблица 1.
Кластеры повышения локальной нейрональной активности при предъявлении распознаваемых изображений по сравнению с нераспознаваемыми
| № | Размер кластера, кол-во вокселей | MNI-координаты локального максимума [x, y, z], мм | t-значение в локальном максимуме | Анатомическая локализация (Л/П – левое/правое полушарие; ПБ – поле Бродмана) |
|---|---|---|---|---|
| Исследование № 1: “0.5Norm + 0.5Scr” > “TotScr” | ||||
| 1 | 1770 | –30 –46 –10 | 14.73 | Л: веретенообразная и парагиппокампальная извилина, гиппокамп (ПБ 20, 28, 34, 35, 36, 37) |
| –42 –73 –7 | 11.72 | Л: нижняя затылочная и средняя височная извилина (ПБ 19, 37) | ||
| –39 –76 20 | 10.66 | Л: верхняя и средняя височная извилина, угловая извилина (ПБ 21, 22, 37, 39) | ||
| 2 | 1687 | 54 –64 11 | 13.71 | П: верхняя и средняя височная извилина, угловая извилина (ПБ 21, 22, 37, 39) |
| 42 –49 –19 | 13.24 | П: веретенообразная и парагиппокампальная извилина, гиппокамп, амигдала (ПБ 20, 28, 34, 35, 36, 37) | ||
| 48 –73 –7 | 11.10 | П: нижняя затылочная и средняя височная извилина (ПБ 19, 37) | ||
| 3 | 97 | –33 32 –16 | 8.71 | Л: орбитофронтальная кора (ПБ 11, 47) |
| 4 | 186 | 18 –55 14 | 7.78 | П: предклинье, задняя поясная кора (ПБ 23, 29, 30) |
| –9 –55 11 | 7.47 | Л: предклинье, задняя поясная кора (ПБ 23, 29, 30) | ||
| 5 | 58 | 36 32 –13 | 7.66 | П: орбитофронтальная кора (ПБ 11, 47) |
| 6 | 59 | –51 –7 –16 | 7.21 | Л: средняя височная извилина (ПБ 21) |
| 7 | 49 | –39 8 29 | 5.68 | П: средняя лобная извилина (ПБ 9) |
| 8 | 26 | 57 –1 –19 | 5.56 | П: средняя височная извилина (ПБ 21) |
| 9 | 18 | –51 32 5 | 5.33 | Л: нижняя лобная извилина, триангулярная часть (ПБ 47) |
| Исследование № 2: “0.5Anim + 0.5Inanim” > “TotScr” | ||||
| 1 | 2658 | 45 –55 14 | 13.24 | П: верхняя и средняя височная извилина, угловая извилина (ПБ 21, 22, 37, 39) |
| 48 –76 –1 | 10.48 | П: нижняя затылочная и средняя височная извилина (ПБ 19, 37) | ||
| 60 –25 23 | 9.36 | П: надкраевая извилина (ПБ 40) | ||
| 54 2 8 | 8.93 | П: роландический оперкулум, средняя островковая кора (ПБ 13, 43) | ||
| 27 –25 –16 | 8.57 | П: парагиппокампальная и веретенообразная извилина, гиппокамп, амигдала (ПБ 35, 36, 37) | ||
| 42 –10 –1 | 6.38 | П: задняя островковая кора (ПБ 13) | ||
| 2 | 3541 | –48 –58 8 | 12.15 | Л: верхняя и средняя височная извилина, угловая извилина (ПБ 21, 22, 37, 39) |
| –42 –82 –4 | 10.26 | Л: нижняя затылочная и средняя височная извилина (ПБ 19, 37) | ||
| –33 –40 –16 | 9.84 | Л: парагиппокампальная и веретенообразная извилина, гиппокамп, амигдала (ПБ 35, 36, 37) | ||
| –63 –34 26 | 8.94 | Л: надкраевая извилина (ПБ 40) | ||
| –36 –13 –1 | 8.81 | Л: задняя островковая кора (ПБ 13) | ||
| –39 2 8 | 7.98 | Л: роландический оперкулум, средняя островковая кора (ПБ 13, 43) | ||
| –18 –61 17 | 7.71 | Л: клин, предклинье, задняя поясная кора (ПБ 23, 29, 30) | ||
| 21 –55 20 | 7.42 | П: клин, предклинье, задняя поясная кора (ПБ 23, 29, 30) | ||
| 12 –55 59 | 7.31 | П/Л: Верхняя теменная долька (ПБ 7) | ||
| 3 | 111 | 51 –7 –16 | 8.78 | Л: средняя височная извилина (ПБ 21) |
| 4 | 92 | –60 –7 –13 | 6.85 | П: средняя височная извилина (ПБ 21) |
| 5 | 108 | –3 62 –7 | 6.67 | Л/П: медиальная префронтальная кора (ПБ 10, 11) |
| 6 | 74 | 12 –25 38 | 6.67 | П: средняя поясная кора (ПБ 24) |
В первом исследовании (“0.5 × Norm + 0.5 × × Scr” > “TotScr”) повышение локальной нейрональной активности обнаружено билатерально в веретенообразных и парагиппокампальных извилинах, гиппокампах, нижних затылочных извилинах, нижних и средних височных извилинах, угловых извилинах, предклинье и орбитофронтальной коре. Во втором исследовании (“0.5 × × Anim + 0.5 × Inanim” > “TotScr”) повышение обнаружено билатерально в тех же областях, а также билатерально в амигдалах, задней и средней островковой коре, надкраевых извилинах, верхних теменных дольках и в медиальной префронтальной коре. Одинаковый уровень нейрональной активности в пробах с распознаваемыми изображениями разного типа наблюдается во множестве структур мозга, среди которых невозможно выделение отдельных кластеров из-за природы “нулевых” эффектов. По сути это один большой кластер, располагающийся между кластерами активаций и деактиваций. Это согласуется с результатами предыдущих фМРТ-исследований, когда выделялись небольшие кластеры значимых изменений BOLD-сигнала, тогда как на бóльшей части мозга таких изменений не наблюдалось. Кроме того, целью данной работы было выявление общих “нулевых звеньев” для двух типов процессов категоризации зрительных стимулов, что подразумевает поиск пересечений между “нулевыми эффектами” двух исследований, проводившийся на следующем этапе. Сопоставление пересечений (conjunction analysis) между результатами значимых изменений и отсутствиями изменений BOLD-сигнала в двух исследованиях позволило установить структуры мозга, которые являются кандидатами в “скрытые звенья” (рис. 3).
Рис. 1.
Дизайн исследования. А – общая схема тестового задания. а) – варианты предъявляемых стимулов для исследования № 1, б) – межстимульный интервал, в) – пробы “пустышки”, предъявляемые с частотой 1/3. “Norm” и “Scr” – неискаженные и частично искаженные изображения одушевленных или неодушевленных объектов. “TotScr” – полностью искаженные изображения. Б – варианты предъявляемых стимулов в исследовании № 2. “Anim” и “Inanim” – неискаженные одушевленные и неодушевленные изображения.
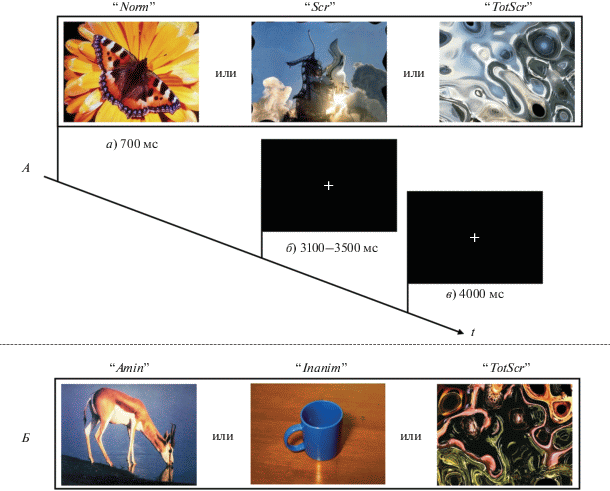
Рис. 2.
Результаты анализа локальной активности. A – категоризация неискаженных и частично искаженных объектов. Б – категоризация одушевленных и неодушевленных объектов. а – области мозга с увеличением BOLD-сигнала при распознавании объекта. б – области мозга, в которых принимается нулевая гипотеза о равенстве средних – сигнал значимо не отличается между категоризацией типов распознаваемых объектов. в – пересечение результатов. Z – координата MNI-пространства по оси Z. L/R – левое/правое полушарие.
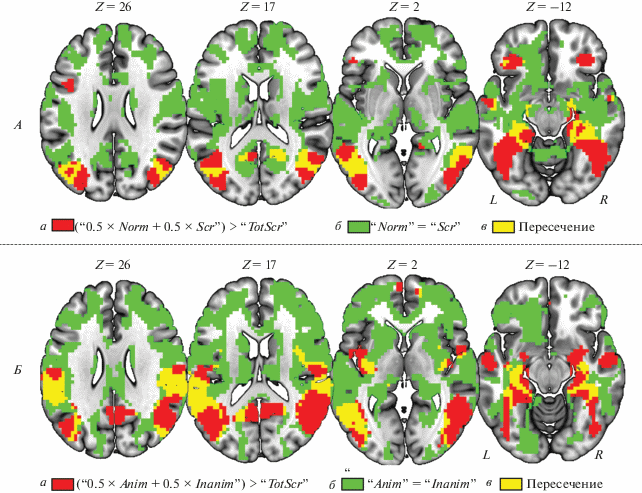
Рис. 3.
Области-кандидаты на роль “скрытых звеньев” и соответствующие им оценки изменения BOLD-сигнала в двух исследованиях (среднее значение BOLD-сигнала в кластере и 90% доверительный интервал). A – каудальная часть внутритеменной борозды билатерально, Б – предклинье билатерально, В – средняя височная извилина (ПБ 37) слева, Г – парагиппокампальные извилины билатерально. L/R – левое/правое полушарие.
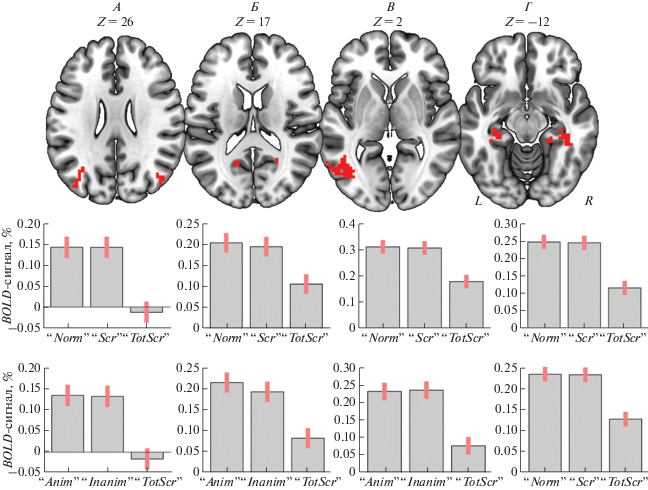
В результате применения анализа графов к показателям функциональной связности было выявлено два “скрытых звена”, играющих ключевую роль в структуре дистантных взаимодействий при обеспечении процессов смысловой категоризации зрительных стимулов в обоих заданиях:
1) каудальная часть средней височной извилины левого полушария (ПБ 37), относящаяся к дорзальной системе внимания, характеризуется высокими показателями локальной эффективности (local efficiency);
2) кора предклинья левого полушария, относящаяся к дефолтной системе мозга, характеризуется высокими показателями степени посредничества (betweeness centrality).
Повоксельный анализ психофизиологических взаимодействий, проводившийся для того, чтобы определить, являются ли данные структуры “скрытыми звеньями”, установил для области интереса в левой средней височной извилине увеличение силы функциональных связей при категоризации неискаженного изображения, по сравнению с частично искаженным с левой нижней лобной извилиной и червячком мозжечка. При этом то же самое звено при категоризации неодушевленного объекта, по сравнению с категоризацией одушевленного, демонстрировало увеличение силы функциональных связей со средними и нижними лобными извилинами обоих полушарий (рис. 4, А, Б, табл. 2).
Рис. 4.
Статистически значимые изменения силы функциональных связей со “скрытыми звеньями” при категоризации объектов разных типов. А – исследование № 1, сравнение “Norm > Scr”: a – “скрытое звено” в левой средней височной извилине, б – левая средняя лобная извилина, в – червячок мозжечка. Б – исследование № 2, сравнение “Inanim > Anim”: a – “скрытое звено” в левой средней височной извилине, г – левая нижняя и средняя лобная извилина, д – правая средняя и нижняя извилина. В – исследование № 1, сравнение “Norm > Scr”: е – “скрытое звено” в левом предклинье, ж – билатерально предклинье, верхняя теменная долька, задняя поясная кора.
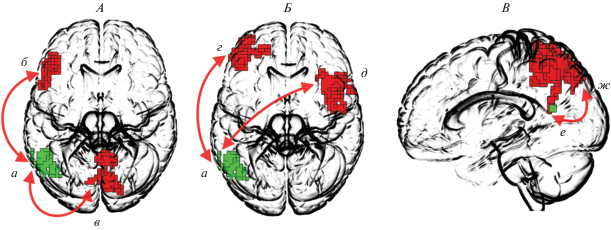
Таблица 2.
Кластеры статистически значимого изменения функциональных связей с двумя выбранными областями интереса (ОИ) при категоризации объектов разных типов
| № | Размер кластера, кол-во вокселей | MNI-координаты локального максимума [x, y, z], мм | t-значение в локальном максимуме | Анатомическая локализация (Л/П – левое/правое полушарие; ПБ – поле Бродмана) |
|---|---|---|---|---|
| ОИ: Левая средняя лобная извилина. Исследование № 1: “Norm” > “Scr” | ||||
| 1 | 208 | 6 –73 –34 | 5.52 | П/Л: мозжечок, пирамида и втулочка червя |
| 12 –58 –13 | 4.61 | П/Л: мозжечок, верхушка червя | ||
| 2 | 210 | –51 17 38 | 5.41 | Л: средняя лобная извилина (ПБ 8, 9) |
| –51 32 26 | 5.39 | Л: нижняя лобная извилина, триангулярная часть (ПБ 46) | ||
| ОИ: Левая средняя лобная извилина. Исследование № 2: “Inanim” > “Anim” | ||||
| 1 | 376 | 39 14 29 | 5.88 | П: нижняя лобная извилина, оперкулярная и триангулярная часть (ПБ 45, 46) |
| 33 26 44 | 3.86 | П: средняя лобная извилина (ПБ 8, 9) | ||
| 39 –1 38 | 3.73 | П: прецентральная и средняя лобная извилина (ПБ 6) | ||
| 2 | 286 | –45 35 14 | 4.25 | Л: средняя лобная извилина, нижняя лобная извилина, триангулярная часть (ПБ 10, 45, 46) |
| ОИ: Левое предклинье Исследование № 1: “Norm” > “Scr” | ||||
| 1 | 712 | –6 –58 50 | 5.44 | Л: предклинье (ПБ 7) |
| 3 –61 65 | 4.35 | П/Л: верхняя теменная долька (ПБ 5, 7) | ||
| 6 –79 44 | 4.33 | П: предклинье, задняя поясная кора (ПБ 7, 31) | ||
Для второй области в левом предклинье удалось установить значимое изменение силы функциональных связей только при восприятии неискаженных изображений, по сравнению с восприятием частично искаженных изображений. Обнаруженные кластеры с повышенными параметрами функциональных связей локализовались билатерально в предклинье, верхней теменной дольке и левой задней поясной коре (рис. 4, B, табл. 2). В задаче по определению одушевленности или неодушевленности изображаемых объектов статистически значимых изменений функциональных связей для области интереса в левом предклинье установлено не было.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В результате проведенного исследования были получены новые данные, расширяющие представления о звеньевом составе мозговых систем обеспечения процессов идентификации и категоризации зрительных стимулов. Показано, что помимо структур вентрального пути и латеральной префронтальной коры в процессы категоризации вовлекаются “скрытые звенья” локализующиеся в области средней височной коры и предклинья левого полушария. Так, процессы идентификации зрительных изображений в условиях категоризации на реальность и одушевленность характеризовались увеличением BOLD-сигнала в областях мозга, относящихся к так называемому вентральному пути обработки [29], а также вентральной и дорзальной нейрональным системам внимания [30, 31]. Это воспроизводит результаты предыдущих исследований и соответствует известным литературным данным о мозговом обеспечении процессов категоризации. Однако целенаправленный поиск “скрытых” звеньев позволил не только выявить новые, ранее не обнаруживаемые структуры мозга, но и прояснить их функциональную роль в системной активности мозга. Так, в соответствии с представлениями о феномене “скрытых звеньев” структуры задней средней височной извилины и задней поясной извилины (предклинье) левого полушария не проявляли своего участия в обеспечении двух исследованных типов категоризации (идентификации объекта как реально существующего и определении одушевленности) путем изменения уровня энергопотребления, но значимо меняли характер дистантных взаимодействий. Подобное вовлечение наблюдалось для обоих изучавшихся типов процессов категоризации, что позволяет предполагать универсальный характер их вовлечения. Полученный результат показателен, так как демонстрирует тот факт, что помимо многократно обнаруживаемых в активационных исследованиях структур мозга, есть другие “скрытые” области, которые не проявляют своего участия изменением локальной активности, а только изменяют характер своего включения в системную активность мозга.
Помимо этого, полученные данные свидетельствуют в пользу того, что “скрытые звенья” могут различаться по своей функциональной роли в качестве звена мозговых систем обеспечения поведения. Так, скрытое звено в левой средней височной извилине в зависимости от типа процесса категоризации меняло набор структур мозга, с которыми устанавливались дистантные взаимодействия. Это позволяет говорить о том, что скрытые звенья могут выступать в роли хабов, которые при смене характера деятельности гибко переключают набор звеньев, с которыми устанавливаются дистантные взаимодействия [4, 6]. Конкретно скрытое звено в левой средней височной извилине может выступать в качестве коннектора, “переключая” связи между разными структурами префронтальной коры: при относительно более сложноорганизованном процессе определения категории по признаку одушевленности. А по сравнению с определением реальности изображенного объекта, функциональные связи устанавливаются с относительно большими территориями латеральной префронтальной коры билатерально. Кроме того, результат применения методов, основанных на теории графов, показал, что скрытое звено в средней лобной извилине левого полушария характеризуется высокими показателями “локальной эффективности” (local efficiency) при обоих типах исследуемых процессов категоризации. Данный показатель отражает как степень функциональной специализации данного звена [32], так и устойчивость подсистемы к выпадению ее отдельных звеньев [33] за счет относительно большего количества межзвеньевых связей внутри специализированной системы. Отчасти полученные в настоящем исследовании данные дополняют результаты, полученные в исследовании M. Corbetta [30]. Известно, что повреждение вентролатеральной префронтальной коры и височной коры правого полушария может приводить к агнозии, которая впоследствии может быть компенсирована за счет сохранных структур вентральной и дорсальной системы внимания левого полушария. Тот факт, что обнаруженное нами скрытое звено локализуется в одной из структур дорсальной системы внимания левого полушария и характеризуется высокими показателями локальной эффективности функциональных связей, подтверждает данное предположение.
Второе скрытое звено локализуется в задней области поясной извилины (предклинье) левого полушария, уровень активности которой напрямую связан с уровнем бодрствования, играющей центральную роль в обеспечении произвольного внимания, необходимого для эффективной реализации текущей деятельности [34]. Функциональная значимость вовлечения данного звена, по всей видимости, связана с обеспечением межсистемных взаимодействий, поскольку оно характеризуется высокими показателями “центральности по посредничеству” (betweenness centrality). Это находит подтверждение в литературных данных, демонстрирующих зависимость между повреждениями этой области и нарушением процессов восприятия и нарушениями сознания [34].
ЗАКЛЮЧЕНИЕ
Таким образом, развитие изучения феномена скрытых звеньев на материале фМРТ-данных, полученных в условиях идентификации и категоризации изображений, позволило: 1) разработать способ объективного определения местонахождения скрытых звеньев, 2) подтвердить ключевое предположение о характере участия “скрытых звеньев” в системной активности мозга и расширить сложившиеся представления о звеньевом составе мозговых систем, вовлекаемых в обеспечение смысловой обработки изображений, 3) продемонстрировать принципиальную возможность разных типов скрытых звеньев, отражающих их роль в конфигурации системной активности мозга.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Комитетом по этике Института мозга человека имени Н.П. Бехтеревой РАН (Санкт-Петербург).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Данное исследование выполнено в рамках РНФ (грант № 19-18-00454).
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Di X., Biswal B.B. Toward Task Connectomics: Examining Whole-Brain Task Modulated Connectivity in Different Task Domains // Cerebral Cortex. 2018. V. 29. № 4. P. 1572.
Медведев С.В., Киреев М.В., Коротков А.Д. Организация нейрональных систем обеспечения целенаправленной деятельности человека: новые данные // Физиология человека. 2018. Т. 44. № 4. С. 131. Medvedev S.V., Kireev M.V., Korotkov A.D. Organization of the brain systems of aim-directed behavior: new data // Human Physiology. 2018. V. 44. № 4. P. 488.
Медведев С.В., Коротков А.Д., Киреев М.В. Cкрытые звенья мозговых систем // Физиология человека. 2019. Т. 45. № 5. С. 110. Medvedev S.V., Korotkov A.D., Kireev M.V. Hidden nodes of the brain systems // Human Physiology. 2019. V. 45. № 5. P. 552.
Ito T., Hearne L.J., Cole M.W. A cortical hierarchy of localized and distributed processes revealed via dissociation of task activations, connectivity changes, and intrinsic timescales // Neuroimage. 2020. V. 221. № 1. P. 117141.
Cole M.W., Reynolds J.R., Power J.D. et al. Multi-task connectivity reveals flexible hubs for adaptive task control // Nat. Neurosci. 2013. V. 16. № 9. P. 1348.
Gratton C., Laumann T.O., Gordon E.M. et al. Evidence for Two Independent Factors that Modify Brain Networks to Meet Task Goals // Cell Reports. 2016. V. 17. № 5. P. 1276.
Kireev M., Slioussar N., Korotkov A.D. et al. Changes in functional connectivity within the fronto-temporal brain network induced by regular and irregular Russian verb production // Front. Hum. Neurosci. 2015. V. 9. № 36. P. 10.
Medvedev S. The hidden systems in the human brain // Int. J.Psychophysiol. 2018. V. 131. № 1. P. S7.
Masharipov R.S., Kireev M.V., Korotkov A.D., Medvedev S.V. P.212 Providing support for the null hypothesis in functional magnetic resonance imaging: testing group-level Bayesian inference // Eur. Neuropsychopharm. 2020. V. 40. Suppl. 1. P. S121.
Fornito A., Zalesky A., Bullmore E. Fundamentals of brain network analysis. Academic Press, Cambridge, 2016. 476 p.
Pan X., Sakagami M. Category representation and generalization in the prefrontal cortex // Eur. J. Neurosci. 2012. V. 35. № 7. P. 1083.
Seger C.A., Miller E.K. Category learning in the brain // Annu. Rev. Neurosci. 2010. V. 33. № 1. P. 203.
Oldfield R.C. The assessment and analysis of handedness: The Edinburgh inventory // Neuropsychologia. 1971. V. 9. № 1. P. 97.
Friston K.J., Zarahn E., Josephs O. et al. Stochastic designs in event-related fMRI // Neuroimage. 1999. V. 10. № 5. P. 607.
Lang P.J., Bradley M.M., Cuthbert B.N. International affective picture System (IAPS): Affective ratings of pictures and instruction manual. Technical Report A-8. University of Florida, Gainesville, FL, 2008. P. 5.
Stojanoski B., Cusack R. Time to wave good-bye to phase scrambling: Creating controlled scrambled images using diffeomorphic transformations // J. Vis. 2014. V. 14. № 12. P. 6.
Neumann J., Lohmann G. Bayesian second-level analysis of functional magnetic resonance images // NeuroImage. 2003. V. 20. № 2. P. 1346.
Friston K.J., Glaser D.E., Henson R.N.A. et al. Classical and Bayesian Inference in Neuroimaging: Applications // NeuroImage. 2002. V. 16. № 2. P. 484.
Friston K.J., Penny W. Posterior probability maps and SPMs // NeuroImage. 2003. V. 19. № 3. P. 1240.
Friston K.J., Williams S., Howard R. et al. Movement-Related effects in fMRI time-series // Magn. Reson. Med. 1996. V. 35. № 3. P. 346.
Poldrack R.A., Mumford J.A., Nichols T.E. Handbook of Functional MRI Data Analysis. Cambridge, United Kingdom: Cambridge University Press, 2011. 239 p.
Kruschke J.K. Rejecting or Accepting Parameter Values in Bayesian Estimation // Adv. Methods Pract. Psychol. Sci. 2018. V. 1. № 2. P. 270.
Bennett C.M., Miller M.B. How reliable are the results from functional magnetic resonance imaging? // Ann. N.Y. Acad. Sci. 2010. V. 1191. № 1. P. 133.
Rissman J., Gazzaley A., D’Esposito M. Measuring functional connectivity during distinct stages of a cognitive task // NeuroImage. 2004. V. 23. № 2. P. 752.
Mumford J.A., Turner B.O., Ashby F.G., Poldrack R.A. Deconvolving BOLD activation in event-related designs for multivoxel pattern classification analyses // NeuroImage. 2012. V. 59. № 3. P. 2636.
Seitzman B.A., Gratton C., Marek S. et al. A set of functionally-defined brain regions with improved representation of the subcortex and cerebellum // NeuroImage. 2020. V. 206. № 1. P. 116290.
McLaren D.G., Ries M.L., Xu G., Johnson S.C. A generalized form of context-dependent psychophysiological interactions (gPPI): A comparison to standard approaches // NeuroImage. 2012. V. 61. № 4. P. 1277.
Gitelman D.R., Penny W.D., Ashburner J., Friston K.J. Modeling regional and psychophysiologic interactions in fMRI: the importance of hemodynamic deconvolution // NeuroImage. 2003. V. 19. № 1. P. 200.
Ungerleider L., Haxby J. “What” and “where” in the human brain // Curr. Opin. Neurobiol. 1994. V. 4. № 2. P. 157.
Corbetta M., Kincade M.J., Lewis C. et al. Neural basis and recovery of spatial attention deficits in spatial neglect // Nat. Neurosci. 2005. V. 8. № 11. P. 1603.
Yeo B.T., Krienen F.M., Sepulcre J. et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity // J. Neurophysiol. 2011. V. 106. № 3. P. 1125.
Stanley M.L., Moussa M.N., Paolini B.M. et al. Defining nodes in complex brain networks // Front. Comput. Neurosci. 2013. V. 7. № 169. P. 14.
Achard S., Bullmore E. Efficiency and cost of economical brain functional networks // PLoS Comput. Biol. 2007. V. 3. № 2. P. e17.
Leech R., Sharp D.J. The role of the posterior cingulate cortex in cognition and disease // Brain. 2014. V. 137. № 1. P. 12.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека