

Физиология человека, 2022, T. 48, № 5, стр. 38-48
Электроэнцефалографические и вегетативные корреляты воображаемых и реальных движений ног у правшей и левшей
О. А. Ведясова 1, *, К. А. Моренова 1, С. И. Павленко 1
1 Самарский национальный исследовательский университет
имени академика С.П. Королева
Самара, Россия
* E-mail: o.a.vedyasova@gmail.com
Поступила в редакцию 13.01.2022
После доработки 01.02.2022
Принята к публикации 18.03.2022
- EDN: XIEQAA
- DOI: 10.31857/S0131164622700047
Аннотация
Изучены изменения спектральной мощности (СМ) электроэнцефалограммы (ЭЭГ) в α-диапазоне и вариабельности сердечного ритма (ВСР) у испытуемых с правым и левым профилями моторного доминирования при воображаемом и реальном сгибании правой и левой стоп. При реализации двигательных заданий выявлена выраженная десинхронизация α-ритма во фронтальных и центральных отведениях и менее значимые его изменения в теменной и затылочной зонах обеих гемисфер. Особенностью правшей при мысленных и реальных движениях ног был вариабельный и асимметричный характер изменений СМ α-ритма в билатеральных отведениях от различных областей коры. У левшей при воображении и выполнении движений десинхронизация α-ритма в разных отведениях имела одинаковую выраженность и межполушарных различий СМ практически не наблюдалось. У правшей в ходе выполнения моторных задач отмечался относительно стабильный вегетативный статус, изменения ВСР возникали в основном при реальных движениях ног. У левшей параметры ВСР менялись сильнее и главным образом при мысленных действиях ногами. Между СМ α-ритма и параметрами ВСР выявлены корреляционные связи, зависящие от вида движения и профиля моторного доминирования испытуемых. Полученные данные отражают особенности механизмов регуляции произвольных движений ног и различия вегетативного обеспечения двигательной активности у правшей и левшей.
Одним из очевидных проявлений функциональной межполушарной асимметрии (ФМА) является моторная асимметрия [1–3], представления о которой развиваются благодаря анализу внутрицентральных, в том числе межполушарных, взаимодействий в процессе воображения двигательных актов [4–6] и их выполнения [7, 8]. Важным инструментом изучения данного аспекта ФМА служит сравнительный анализ электроэнцефалограммы (ЭЭГ) у правшей и левшей, выявляющий особенности центральных механизмов регуляции движений у лиц с разными профилями моторного доминирования [7].
Паттерн ЭЭГ при воображаемых и реальных движениях изучен преимущественно на примере рук. Основным маркером корковой активности в этих случаях являются синхронизация или десинхронизация α-ритма [9, 10], причем двигательное воображение обычно сопровождается уменьшением μ-ритма, регистрируемого в α-диапазоне над сенсомоторными областями коры больших полушарий [11]. У правшей воображаемые движения правой и левой руки вызывают контралатеральную десинхронизацию μ-ритма и его ипсилатеральную синхронизацию [12]. У левшей при этом наблюдаются синфазные изменения когерентности α-волн справа и слева, что говорит о преобладании межполушарного взаимодействия. При реальных движениях у правшей и левшей выявлена зависимость увеличения/уменьшения когерентности ритмов ЭЭГ разных частот от использования ведущей или неведущей руки [7]. Что касается исследований динамики ЭЭГ при мысленных и реальных движениях ног, то они не многочисленны и аспект ФМА в них практически не рассматривается [13, 14].
Выполнение двигательных актов отражается на вегетативном статусе организма, что подтверждается изменениями вариабельности сердечного ритма (ВСР). Во время движений некоторые показатели ВСР (SDNN, RMSSD) снижаются, а в динамике спектральных характеристик кардиоритма выявляется увеличение доли HF-волн, уменьшение LF-компонента и индекса вагосимпатического взаимодействия [15]. Закономерно ожидать определенные вегетативные реакции и на фоне воображения движений, поскольку оно основано на создании мысленного эквивалента двигательного акта, что является типичной когнитивной деятельностью [14, 16], разные виды которой сопровождаются изменениями ВСР [17–19]. В ряде работ отмечены функциональная значимость α-ритма в регуляции мыслительных процессов и корреляции между паттернами α-активности и ВСР [20–22]. Показано, что на разных этапах когнитивной деятельности изменения межполушарной когерентности α-ритма совпадают с динамикой RR-интервалов и спектральных параметров ВСР, отражающих активность отделов вегетативной нервной системы (ВНС) [20]. Значительно меньше внимания уделяется связям между ВСР и ЭЭГ в процессе выполнения в уме двигательных заданий. Имеются отдельные данные о корреляциях активности механизмов кардиорегуляции с изменениями ЭЭГ в центральных отведениях при мысленных движениях рук [12]. Выявлены замедление частоты сердечных сокращений (ЧСС) и сопутствующая этому перицентральная десинхронизация ЭЭГ в ситуации планирования и подготовки движений ног [23]. Однако вопрос о взаимосвязи ВСР с ЭЭГ при воображении и выполнении движений ног у лиц с разными профилями моторного доминирования остается открытым. Вместе с тем, его разработка важна для развития представлений о произвольной регуляции движений и понимания вегетативных механизмов адаптации к двигательным нагрузкам у правшей и левшей, а также для решения прикладных задач в аспекте создания интерфейсов мозг-компьютер.
Цель исследования состояла в анализе изменений спектральной мощности (СМ) α-ритма и параметров ВСР и выявлении их корреляционных связей при мысленных и реальных движениях ног у правшей и левшей.
МЕТОДИКА
Исследование выполняли на 26 правшах и 22 левшах в возрасте 18–23 лет. Индивидуальный профиль моторного доминирования испытуемых оценивали по результатам тестов на определение ведущей руки и ноги [1, 7]. ЭЭГ и ВСР регистрировали в состоянии покоя и при выполнении двигательных задач в виде реальных и мысленных сгибаний и разгибаний ног в голеностопном суставе. Испытуемые размещались в удобном кресле в темной звукоизолированной комнате и находились с закрытыми глазами в течение всего исследования. Двигательные задания выполняли в следующем порядке: реальное движение правой стопы, реальное движение левой стопы, воображаемое движение правой стопы, воображаемое движение левой стопы.
Для регистрации ЭЭГ использовали нейровизор NVX 36 digital DC EEG (Россия) и набор отводящих электродов, располагаемых на голове по системе “10–20”. Электроды фиксировали при помощи шлема-сетки, референтный (объединенный ушной) электрод закрепляли на мочке правого уха. Сначала записывали ЭЭГ в состоянии покоя (в течение 1 мин). После этого давали словесную команду к выполнению двигательного задания (например, “согните правую стопу”). Через 3 с следовала команда “разогните стопу”. Данный сценарий применяли ко всем заданиям, пауза между которыми составляла 10 с. Учитывая значимость α-ритма в осуществлении движений [10, 12], мы сопоставляли паттерны волн ЭЭГ α-диапазона (8–13 Гц) в симметричных фронтальных (Fp1–Fp2, F7–F8), центральных (С3–С4), париетальных (Р3–Р4) и окципитальных (О1–О2) отведениях. ЭЭГ-сигналы усиливали, пропускали через фильтр нижних частот 100 Гц и оцифровывали с частотой дискретизации 250 Гц с помощью программы “Нейрокортекс-С” (версия 2.10, Россия). Анализировали СМ α-ритма (мкВ2) в покое и при выполнении двигательных заданий.
Параметры ВСР регистрировали с помощью приборно-программного обеспечения “ЭЛОКС-01С2” (Россия), используя фотооптический датчик, надеваемый на указательный палец левой руки. Ритмограммы в покое и при реальных и мысленных движениях записывали в течение 5 мин. Каждое задание испытуемые выполняли по команде экспериментатора и повторяли с 15-секундным интервалом на протяжении вышеуказанного времени. Оценивали статистические и диагностические показатели ВСР: ЧСС (HR, уд./мин), коэффициенты активности симпатического (SIM, усл. ед.) и парасимпатического (PAR, усл. ед.) отделов ВНС, индекс напряжения по Баевскому (IB, усл. ед.). Анализ спектра ВСР проводили по общей СМ колебаний кардиоритма (Total, мс2), СМ в диапазонах очень низких (VLF, мс2), низких (LF, мс2) и высоких (HF, мс2) частот и индексу централизации (IC, усл. ед.) [24].
Для статистической обработки применяли программный пакет SigmaPlot 12.5 (Systat Incorporated, США). Нормальность распределения выборок оценивали по критерию Шапиро–Уилка. Данные, имеющие нормальное распределение, представлены как средние значения (M) ± стандартная ошибка среднего (m) и как разница средних значений (D) ± стандартная ошибка разницы (mD). Для выборок, не имеющих нормального распределения, указаны медианы (Me) c межквартильным размахом (Р25–Р75). Достоверность различий выявляли в тестах Стьюдента, Уилкоксона и Манна–Уитни. Коэффициенты корреляции (r) определяли по Спирмену. Статистически значимыми считали различия при p < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В аспекте зависимости ЭЭГ-коррелятов реальных и мысленных движений от ФМА особый интерес вызывают фронтальные отведения F7 и F8, где у правшей и левшей десинхронизация α-ритма была сильнее, чем в других областях коры. У правшей в F7 СМ α-волн при всех двигательных заданиях снижалась относительно покоя в среднем на 6.8 ± 1.8 мкВ2 (p < 0.05). В F8 уменьшение СМ происходило только при воображаемом движении левой и реальном действии правой ногой в среднем на 5.1 ± 1.2 мкВ2 (p < 0.05). Следует отметить межполушарную асимметрию α-ритма в F7–F8. У правшей (рис. 1, А) фоновая СМ слева (F7) была на 26.7% (p < 0.05) выше, чем справа (F8). В случае воображения движения ведущей ногой асимметрия сохранялась за счет левостороннего преобладания СМ на 30.9% (p < 0.05). При выполнении других заданий асимметрия α-волн у правшей сглаживалась за счет преимущественной десинхронизации слева (F7). У левшей в покое асимметрия α-ритма в F7−F8 также выражалась левосторонним преобладанием СМ (на 30.8%; р < < 0.05). При мысленных и реальных движениях ног у них, в отличие от правшей, в F7 и F8 СМ α-ритма снижалась примерно одинаково, в среднем на 4.6 ± 1.6 мкВ2 (p < 0.05) от уровня покоя, и поэтому межполушарные различия сохранялись (рис. 1, Б).
Рис. 1.
Межполушарные различия спектральной мощности (СМ) фронтального α-ритма у правшей (А) и левшей (Б). а – Левосторонние отведения, б – правосторонние; ВДП и ВДЛ – воображаемые движения правой и левой ног; РДП и РДЛ – реальные движения правой и левой ног; * (p < 0.05), ** (p < 0.01) – достоверность изменений относительно покоя; # (p < 0.05), ## (p < 0.01) – достоверность межполушарных различий.
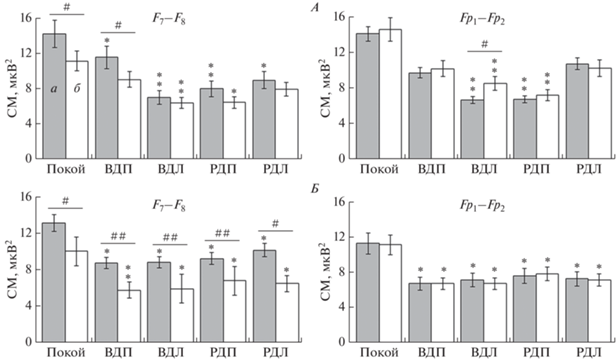
В передних лобных отведениях ЭЭГ регистрировалась иная картина. В частности, фоновая асимметрия α-волн между Fp1 и Fp2 у испытуемых отсутствовала. У правшей СМ α-ритма снижалась при воображении движения неведущей ногой, в том числе в Fp1 на 7.8 ± 1.5 мкВ2 и Fp2 на 5.9 ± ± 1.3 мкВ2 (р < 0.01) относительно покоя, что приводило к правостороннему преобладанию α-волн (рис. 1, А). Во время реального действия ведущей ногой у правшей на ЭЭГ наблюдалось симметричное снижение СМ в среднем на 7.6 ± 1.8 мкВ2 (р < 0.01). У левшей при всех заданиях СМ α-ритма одинаково уменьшалась в Fp1 и Fp2, в среднем на 4.7 ± 0.9 мкВ2 (p < 0.05) от уровня покоя, и, таким образом, межполушарных различий ЭЭГ в данном диапазоне не наблюдалось (рис. 1, Б).
В центральных (С3, С4), париетальных (P3, P4) и окципитальных (О1, О2) отведениях изменения СМ α-ритма у правшей при мысленных и реальных движениях правой и левой ног имели примерно одинаковую выраженность (табл. 1). У левшей же более значимые реакции отмечались при задачах на воображение. Например, при мысленном действии ведущей ногой СМ α-ритма в С3, С4 и О2 снижалась у них сильнее, чем при реальном движении на 8.4, 8.6 и 21.0% соответственно. Что касается межполушарных различий, то у правшей во время реального движения правой ногой устанавливалась асимметрия α-ритма в О1−О2 за счет преобладания СМ справа, тогда как у левшей СМ α-ритма распределялась асимметрично в покое в P3−P4, преобладая слева (табл. 1).
Таблица 1.
Спектральная мощность α-ритма (мкВ2) в центральных, париетальных и окципитальных отведениях у правшей и левшей при выполнении двигательных заданий
| Отведения | Покой | ВДП | ВДЛ | РДП | РДЛ |
|---|---|---|---|---|---|
| Правши | |||||
| C3 | 16.7 ± 2.8 | 13.7 ± 1.6* | 8.5 ± 3.2* | 14.8 ± 2.4* | 11.2 ± 3.1* |
| C4 | 15.1 ± 2.4 | 10.3 ± 2.9* | 7.6 ± 2.8* | 8.5 ± 2.1** | 11.9 ± 1.8* |
| P3 | 18.9 ± 3.3 | 15.4 ± 0.7 | 13.7 ± 2.9*** | 10.8 ± 2.0*** | 13.4 ± 1.3*** |
| P4 | 19.5 ± 2.0 | 14.6 ± 0.9 | 12.3 ± 2.7** | 12.0 ± 1.8*** | 14.3 ± 2.1* |
| O1 | 17.9 ± 1.7 | 13.3 ± 2.7* | 14.1 ± 1.2* | 9.3 ± 2.9***, # | 12.1 ± 2.6*** |
| O2 | 18.2 ± 3.2 | 13.7 ± 2.4*** | 14.0 ± 1.5* | 11.1 ± 3.2**, # | 11.7± 2.9** |
| Левши | |||||
| C3 | 14.5 ± 2.3 | 9.9 ± 1.6*** | 8.4 ± 2.7**, + | 8.7 ± 0.8*** | 9.5 ± 1.1**, + |
| C4 | 13.6 ± 1.8 | 8.9 ± 1.1** | 8.1 ± 2.3**, + | 10.1 ± 0.7* | 9.6 ± 1.5**, + |
| P3 | 20.8 ± 3.5# | 13.5 ± 2.4*** | 12.7 ± 2.1*** | 13.9 ± 2.9*** | 12.9 ± 2.6** |
| P4 | 17.5 ± 2.7# | 12.1 ± 2.7** | 11.6 ± 1.9*** | 11.7 ± 2.1*** | 13.1 ± 2.8** |
| O1 | 17.1 ± 1.8 | 11.2 ± 2.0*** | 10.2 ± 1.7*** | 14.8 ± 1.8*** | 12.0 ± 1.4** |
| O2 | 16.5 ± 2.2 | 14.7 ± 1.8 | 11.0 ± 1.4***, + | 11.9 ± 1.5 | 14.9 ± 1.9+ |
Примечание: ВДП и ВДЛ – воображаемые движения правой и левой стоп; РДП и РДЛ – реальные движения правой и левой стоп; * (p < 0.05), ** (p < 0.01), *** (p < 0.001) – достоверность изменений относительно покоя; # (p < 0.05) – достоверность межполушарных различий; + (p < 0.05) – достоверность различий между изменениями при ВДЛ и РДЛ.
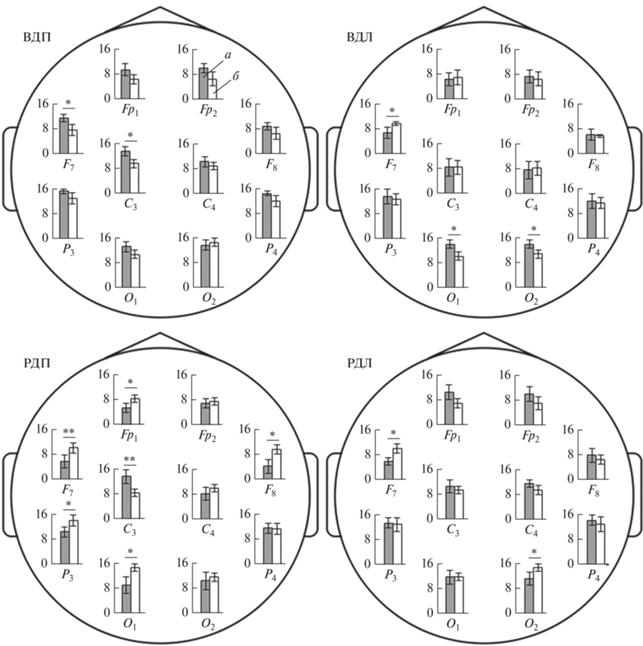
Связь паттерна ЭЭГ с профилем моторного доминирования в процессе движений подтвердилась результатами межгруппового сравнения абсолютных значений СМ α-ритма у правшей и левшей. Выявлено, что во время двигательного воображения более высокий уровень СМ α-ритма отмечался у правшей, в частности, в отведениях F7 и C3 для ведущей ноги, а в O1 и O2 – для неведущей. У левшей в большинстве отведений ЭЭГ α-ритм преобладал при реальных движениях, особенно неведущей ногой (рис. 2).
Рис. 2.
Различия спектральной мощности (СМ) α-ритма (мкВ2) у правшей и левшей при выполнении двигательных задач. а – Правши, б – левши; ВДП и ВДЛ – воображаемые движения правой и левой ног; РДП и РДЛ – реальные движения правой и левой ног; * (p < 0.05), ** (p < 0.01) – достоверность межгрупповых различий.
Изучение динамики ВСР в ходе выполнения двигательных задач также показало наличие некоторых различий между испытуемыми с разными типами ФМА. Например, для правшей было типичным увеличение значений HR во время реальных движений как правой, так и левой ногами в среднем на 13.9% (p < 0.05) от исходного уровня (58.7 (56.3–63.1) уд./мин). У левшей, напротив, рост HR происходил при воображаемых движениях ног в среднем на 12.3% (p < 0.05) от исходной величины (57.9 (55.8–65.1) уд./мин). Это сочеталось с динамикой SIM, величина которого в ходе воображения и выполнения действий левой ногой составляла у левшей 3.0 (1.8–4.0) и 2.0 (1.8–2.3) усл. ед. соответственно, что в 3 и 2 раза (p < < 0.05) превышало значения SIM у правшей. Такая разница, возможно, связана с различиями в активации вегетативных механизмов при решении моторных задач.
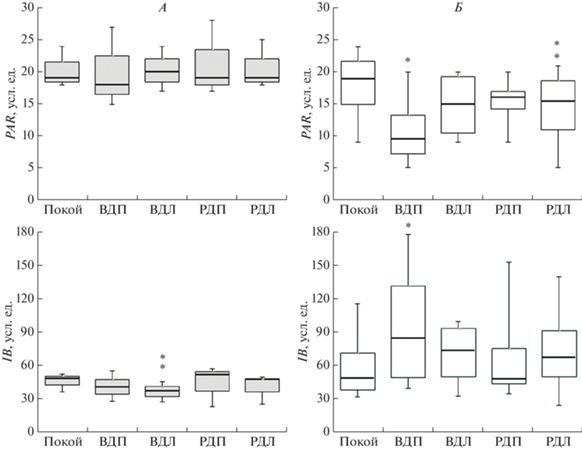
Данному заключению соответствуют изменения PAR и IB (рис. 3). Так, PAR менялся только у левшей, снижаясь на 38.0 и 19.7% (p < 0.05) от фона (19.0 (15.0–21.8) усл. ед.) при воображении движения правой и реальном движении левой ногой соответственно. Изменения IB наблюдались у правшей и левшей при мысленном действии неведущей ногой и имели разную направленность. У правшей IB снижался при мысленном движении левой ногой на 20.5% (p < 0.01), а у левшей увеличивался при аналогичном действии правой ногой на 56.1% (p < 0.05), что говорит о разной напряженности регуляторных систем у лево- и правополушарных лиц. На это также указывают результаты спектрального анализа ВСР, который выявил статистически значимые изменения только у левополушарных испытуемых в виде уменьшения LF-компонента от 9545 (6021–9882) мс2 в покое до 5358 (3449–5744) мс2 во время реального движения ведущей ногой (p < 0.05).
Рис. 3.
Изменения значений PAR и IB у правшей и левшей при выполнении двигательных задач. А (темные боксы) – правши, Б (светлые) – левши (на боксах указаны: медианы – линия внутри боксов; квартили – верхняя и нижняя границы боксов; 10-й и 90-й перцентили – нижняя и верхняя границы “усов”); ВДП и ВДЛ – воображаемые движения правой и левой ног; РДП и РДЛ – реальные движения правой и левой ног; * (p < 0.05), ** (p < < 0.01) – достоверность изменений относительно покоя.
Для оценки взаимодействия процессов центральной регуляции движений и механизмов их вегетативного обеспечения у испытуемых с разными типами ФМА мы исследовали корреляционные связи СМ α-ритма и параметров ВСР. Установлено, что в покое у правшей СМ α-волн во всех отведениях коррелировала положительно (p < 0.05) с VLF и отрицательно (p < 0.05) с LF, TP и IC. У левшей в покое наблюдались положительные корреляции (p < 0.05) α-волн во фронтальных отведениях с HF, TP и PAR, отрицательные (p < 0.05) – с SIM, IB и IC. При движениях ног (мысленных и реальных) у правшей отмечались корреляции ВСР с α-ритмом в большинстве отведений ЭЭГ, у левшей доминировали связи ВСР со СМ фронтального α-ритма. Следует обратить внимание на то, что более выраженные корреляции формировались при воображаемых движениях, однако у правшей они количественно преобладали при мысленном действии ведущей ногой, тогда как у левшей число связей при мысленных движениях обеих ног было примерно одинаковым (табл. 2).
Таблица 2.
Коэффициенты корреляции (r) между параметрами ВСР и СМ α-ритма в разных отведениях у правшей и левшей при воображении движений
| Воображение движения правой ноги | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| параметры | правши | |||||||||
| Fp1 | Fp2 | F7 | F8 | C3 | C4 | P3 | P4 | O1 | O2 | |
| HR | –0.64 | –0.76 | ||||||||
| VLF | –0.65 | –0.63 | –0.55 | –0.61 | –0.77 | –0.92 | –0.97 | |||
| LF | –0.95 | –0.84 | –0.90 | –0.93 | –0.89 | –0.89 | –0.85 | –0.79 | –0.75 | –0.96 |
| HF | –0.53 | –0.58 | ||||||||
| TP | –0.83 | –0.89 | –0.97 | –0.86 | –0.92 | –0.95 | –0.94 | –0.89 | –0.86 | –0.88 |
| SIM | 0.97 | 0.98 | 0.89 | 0.98 | 0.92 | 0.77 | 0.91 | 0.94 | 0.89 | 0.66 |
| PAR | 0.54 | |||||||||
| IB | 0.77 | 0.78 | 0.84 | 0.79 | 0.64 | 0.89 | 0.94 | 0.96 | ||
| IC | –0.63 | –0.61 | –0.53 | –0.59 | –0.75 | –0.90 | –0.96 | |||
| параметры | левши | |||||||||
| Fp1 | Fp2 | F7 | F8 | C3 | C4 | P3 | P4 | O1 | O2 | |
| HR | 0.97 | 0.74 | –0.50 | |||||||
| VLF | 0.70 | |||||||||
| LF | 0.97 | 0.94 | 0.97 | 0.98 | 0.61 | 0.87 | 0.93 | |||
| HF | 0.96 | 0.91 | 0.93 | 0.97 | 0.74 | 0.90 | 0.95 | |||
| TP | 0.95 | 0.97 | 0.97 | 0.98 | 0.69 | 0.92 | 0.92 | |||
| SIM | –0.92 | –0.82 | –0.92 | –0.90 | –0.67 | –0.93 | –0.50 | |||
| PAR | 0.97 | 0.95 | 0.94 | 0.97 | 0.68 | 0.94 | 0.88 | |||
| IB | –0.80 | –0.68 | –0.76 | –0.77 | –0.67 | –0.72 | –0.50 | |||
| IC | –0.64 | –0.60 | –0.72 | –0.65 | –0.82 | |||||
| Воображение движения левой ноги | ||||||||||
| параметры | правши | |||||||||
| Fp1 | Fp2 | F7 | F8 | C3 | C4 | P3 | P4 | O1 | O2 | |
| HR | –0.65 | –0.80 | –0.97 | |||||||
| VLF | –0.67 | –0.82 | –0.98 | |||||||
| LF | –0.98 | –0.98 | –0.92 | –0.97 | –0.86 | –0.73 | –0.91 | –0.93 | –0.94 | –0.40 |
| HF | –0.63 | 0.74 | ||||||||
| TP | –0.87 | –0.88 | –0.98 | –0.83 | –0.66 | –0.48 | –0.98 | –0.98 | –0.91 | |
| PAR | 0.64 | 0.90 | ||||||||
| IB | –0.66 | –0.90 | ||||||||
| IC | –0.58 | –0.56 | –0.63 | –0.81 | –0.92 | –0.94 | ||||
| параметры | левши | |||||||||
| Fp1 | Fp2 | F7 | F8 | C3 | C4 | P3 | P4 | O1 | O2 | |
| HR | 0.61 | 0.55 | 0.66 | |||||||
| VLF | 0.73 | |||||||||
| HF | 0.79 | 0.69 | 0.74 | |||||||
| TP | 0.67 | 0.52 | 0.53 | |||||||
| SIM | –0.91 | –0.98 | –0.92 | –0.97 | –0.78 | –0.95 | –0.83 | –0.85 | –0.77 | –0.91 |
| PAR | 0.92 | 0.97 | 0.97 | 0.98 | 0.69 | 0.89 | 0.77 | 0.77 | 0.67 | 0.92 |
| IB | –0.90 | –0.84 | –0.93 | –0.90 | –0.69 | –0.54 | –0.53 | –0.90 | ||
| IC | –0.68 | –0.71 | –0.64 | –0.61 | –0.97 | –0.90 | –0.79 | –0.98 | –0.93 | –0.63 |
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Из полученных результатов следует, что характерной реакцией на воображение и выполнение движений ногами у правшей и левшей является десинхронизация α-ритма, более выраженная во фронтальных и центральных областях коры, что соответствует данным об активации при реальном и мысленном движениях одних и тех же нейронных сетей [25]. Считается, что инициация произвольного движения и его выполнение начинаются с возбуждения префронтальных регионов коры, распространяющегося в первичные моторные и сенсомоторные зоны [26]. Вовлечение фронтальных областей в двигательное воображение может объясняться модулирующим влиянием премоторных зон на эфферентные сигналы из первичной моторной коры [27]. Десинхронизация или синхронизация α-ритма в центральных отведениях С3–С4 расценивается как основной показатель когнитивной и двигательной деятельности человека [12, 14, 28], а кроме того, десинхронизация фронтального и центрального α-ритма может быть связана с активацией мотивационной системы и усилением внимания [22].
В настоящем исследовании на ЭЭГ у испытуемых с разными типами ФМА регистрировалось снижение СМ центрального α-ритма при реальных и воображаемых движениях обеих ног, однако при использовании левой ноги в отведении С3 у левшей отмечался более низкий уровень СМ, чем у правшей. Низкая мощность α-ритма в левой центральной области соотносится с высокой тревожностью [22], которая является особенностью лиц с доминированием правого полушария [2, 29, 30]. С позиций ФМА также важно отметить разную степень изменений α-ритма в симметричных фронтальных отведениях F7 и F8 при выполнении двигательных задач у левополушарных испытуемых и их сходную выраженность у правополушарных. Это согласуется с представлениями о более лабильных механизмах мозговой активности правшей, что позволяет им быстрее адаптироваться к изменяющимся условиям, по сравнению с левшами [7, 31]. Интересно добавить, что у правшей снижение СМ α-волн в Fp1, Fp2, F7, F8 преобладало в случае реального действия ведущей ногой, что в целом совпадает с результатами других исследований [9]. У левшей такая реакция доминировала при воображаемом действии ведущей ногой и чаще возникала в отведениях C3 и C4. При задачах на неведущую ногу у правшей более выраженное угнетение α-волн отмечалось во фронтальных отведениях ЭЭГ при воображении движения, а у левшей эффекты мысленного и реального движений неведущей ногой проявлялись одинаковым уровнем десинхронизации по всей коре. Эти данные свидетельствуют о зональных особенностях активности нейронных сетей, участвующих в создании двигательных образов, подготовке к движению и его осуществлении у лиц с разными типами ФМА. Со ссылкой на представления о динамической нейронной сети, обеспечивающей мысленные двигательные манипуляции [4], и результаты наших предыдущих исследований [31] можно считать, что у правшей данная нейросеть более тесно связана с моторной и премоторной корой, чем у левшей.
В управлении двигательными актами на этапе подготовки к движению и в ходе его выполнения также участвует теменно-затылочная кора [26], что обусловлено необходимостью построения перцептивных моделей движения, особенно при его мысленном представлении [5]. В нашей работе включение теменных и затылочных областей в воображение и выполнение движений ног подтвердилось уменьшением СМ α-ритма в Р3, Р4, О1 и О2 у всех испытуемых, однако у левшей в О2 такой эффект отмечался только в случае мысленного действия ведущей ногой, что, согласно данным литературы, может быть связано с сенсорной модальностью, в рамках которой формируется движение или его образ [32]. Например, угнетение α-волн в теменно-затылочных областях объясняется преимущественно зрительным типом воображения [10], которое, возможно, у обследованных левшей менее выражено, чем у правшей. В целом задача выявления конкретных различий паттерна ЭЭГ при движениях правой и левой ног требует дальнейшего изучения, тем более что некоторые авторы указывают на наблюдаемое при этом сходство активации корковых зон [12].
В условиях воображения и выполнения движений ногами тип ФМА влиял не только на паттерн ЭЭГ, но и на параметры ВСР, изменения которых у правшей и левшей имели некоторые особенности. К примеру, значения HR у правшей возрастали при реальных движениях, а у левшей – при воображаемых, что, на наш взгляд, связано с особенностями активации регуляторных механизмов. Как известно, HR увеличивается при физической и умственной нагрузке, психологическом стрессе [15]. Рост HR также расценивается как признак концентрации на осуществлении целенаправленной деятельности, тогда как снижение HR трактуется в качестве кардио-компонента ориентировочного рефлекса [33], сопровождающего целенаправленный акт. С учетом этого, можно полагать, что напряжение механизмов кардиорегуляции при реальном движении выше у правшей и связано с высокой степенью психического усилия, сосредоточенностью на выполняемом действии. Воображаемые движения ног, очевидно, вызывают большее напряжение вегетативных механизмов у левшей, что дополнительно подтверждается снижением у них PAR и ростом IB и может быть обусловлено высокой эмоциональностью и чувствительностью к стрессам правополушарных лиц [7].
В пользу представлений об особенностях вегетативного обеспечения двигательных актов у правшей и левшей также говорят результаты корреляционного анализа связей α-ритма с ВСР, в характере которых у представителей с разными типами моторной асимметрии отмечались как сходства, так и различия. Так, существенным общим признаком правшей и левшей при выполнении двигательных задач была корелляция между десинхронизацией α-волн и ростом IC, отражающим преобладание центрального контура регуляции сердца над автономным [24]. В тоже время у левшей, в отличие от правшей, были выявлены устойчивые отрицательные связи α-ритма с SIM и IB, что, опираясь на мнение [34], можно объяснить более сильной активацией центрального контура регуляции синусного узла при моторных актах у лиц с доминированием правого полушария. Различия между группами испытуемых проявились также в динамике положительных корреляционных связей α-ритма и ВСР, количество которых при воображении и выполнении движений ног преобладало у левшей. На основании установленных закономерностей допустимо говорить о различиях в динамическом взаимодействии механизмов регуляции движений и кардиоритма у правшей и левшей в ходе мысленных и реальных двигательных актов ногами.
ЗАКЛЮЧЕНИЕ
Таким образом, у лиц с правым и левым профилями моторного доминирования двигательный контроль осуществляется при разной степени вовлечения кортикальных нейронных сетей и механизмов вегетативной регуляции сердечного ритма в процессы воображения и выполнения движений ног. Для правшей при этом типичен более лабильный характер изменений ЭЭГ и стабильный вегетативный статус организма, что, вероятно, позволяет им быстрее и легче адаптироваться к перестройке двигательных программ. У левшей воображение и выполнение двигательных актов ногами вызывает выраженную десинхронизацию α-ритма, сконцентрированную преимущественно во фронтальных и центральных областях мозга, в сочетании со значительным ростом напряженности вегетативных механизмов. Выявленная взаимосвязь в динамике СМ α-волн и параметров ВСР у правшей и левшей отражает особенности взаимодействия механизмов произвольного моторного контроля и отделов мозга, участвующих в вегетативном обеспечении движений ног у лиц с разными профилями моторного доминирования. Очевидно, что левшам присущи более тесные связи коры больших полушарий со структурами, обеспечивающими симпатический контроль висцеральных функций, что определяет у них большую, чем у правшей, активацию центрального контура регуляции кардиоритма во время выполнения и, особенно, воображения движений ног.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены локальным биоэтическим комитетом Самарского национального исследовательского университета им. академика С.П. Королева (Самара).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа поддержана РФФИ (грант № 18-29-14 073).
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Вклад авторов в публикацию. О.А. Ведясова – идея, планирование, разработка дизайна и координация исследования, анализ и трактовка результатов, подготовка текста, оформление рукописи. К.А. Моренова – участие в планировании исследования, подготовка и проведение исследования, расчеты и анализ результатов, подготовка рисунков и текста, оформление рукописи. С.И. Павленко – участие в проведении исследования, анализ результатов, подготовка текста, оформление рукописи.
Список литературы
Брагина Н.Н., Доброхотова Т.А. Функциональные асимметрии человека. М.: Медицина, 1988. 240 с. Bragina N.N., Dobrokhotova T.A. Functional asymmetry of a human. Moscow: Medicine, 1988. 240 p.
Игнатова Ю.П., Макарова И.И., Зенина О.Ю., Аксенова А.В. Современные аспекты изучения функциональной межполушарной асимметрии мозга (обзор литературы) // Экология человека. 2016. № 9. С. 30. Ignatova J.P., Makarova I.I., Zenina O.J., Aksenova A.V. [Current aspects of functional hemispheric asymmetry studying (literature review)] // Ekologiya Cheloveka. 2016. № 9. P. 30.
Berenguer-Rocha M., Baltar A., Rocha S. et al. Interhemispheric asymmetry of the motor cortex excitability in stroke: relationship with sensory-motor impairment and injury chronicity // Neurol. Sci. 2020. V. 41. № 9. P. 2591.
Sasaoka T., Mizuhara H., Inui T.J. Dynamic parieto-premotor network for mental image transformation revealed by simultaneous EEG and fMRI measurement // Cogn. Neurosci. 2014. V. 26. № 2. P. 232.
Lazurenko D.M., Kiroy V.N., Aslanyan E.V. et al. Electrographic properties of movement-related potentials // Neurosci. Behav. Physiol. 2018. V. 48. № 9. P. 1078.
Liburkina S.P., Vasilyev A.N., Yakovlev L.V. et al. A motor imagery-based brain-computer interface with vibrotactile stimuli // Neurosci. Behav. Physiol. 2018. V. 48. № 9. P. 1067.
Жаворонкова Л.А. Правши – левши: межполушарная асимметрия электрической активности мозга человека. М.: Наука, 2006. 222 c. Zhavoronkova L.A. Right-handers – left-handers: interhemispheric asymmetry of electrical activity of the human brain. M.: Nauka, 2006. 222 p.
Боброва Е.В., Богачева И.Н., Ляховецкий В.А. и др. Запоминание последовательностей движений правой и левой руки правшами и левшами: векторное кодирование // Физиология человека. 2017. Т. 43. № 1. С. 18. Bobrova E.V., Bogacheva I.N., Lyakhovetskii V.A. et al. Memorization of sequences of right and left hand movements in right- and left-handers: vector coding // Human Physiology. 2017. V. 43. № 1. P. 13.
Morash V., Bai O., Furlani S. Classifying EEG signals preceding right hand, left hand, tongue, and right foot movements and motor imageries // Clin. Neurophysiol. 2008. V. 119. № 11. P. 2570.
Фролов А.А., Федотова И.Р., Гусек Д., Бобров П.Д. Ритмическая активность мозга и интерфейс мозг-компьютер, основанный на воображении движений // Успехи физиологических наук. 2017. Т. 48. № 3. С. 72. Frolov A.A., Fedotova I.R., Husek D., Bobrov P.D. [Rhythmic brain activity and brain computer interface based on motor imagery] // Usp. Fiziol. Nauk. 2017. V. 48. № 3. P. 72.
Xiao R., Qi X., Patino A. et al. Characterization of infant mu rhythm immediately before crawling: A high-resolution EEG study // NeuroImage. 2017. V. 146. P. 47.
Alonso-Valerdi L.M., Gutiérrez-Begovich D.A., Argüello-García J. et al. User experience may be producing greater heart rate variability than motor imagery related control tasks during the user-system adaptation in brain-computer interfaces // Front. Physiol. 2016. № 7. P. 279.
Боброва Е.В., Решетникова В.В., Фролов А.А., Герасименко Ю.П. Воображение движений нижних конечностей для управления системами “интерфейс мозг-компьютер” // Ж. высш. нервн. деят. им. И.П. Павлова. 2019. Т. 69. № 5. С. 529. Bobrova E.V., Reshetnikova V.V., Frolov A.A., Gerasimenko Y.P. [Motor imagery of lower limbs movements to control brain-computer interface] // Zh. Vyssh. Nerv. Deyat. Im. I.P. Pavlova. 2019. V. 69. № 5. P. 529.
Tariq M., Trivailo P.M., Simic M. Mu-beta event-related (de)synchronization and EEG classification of left-right foot dorsiflexion kinaesthetic motor imagery for BCI // PLoS One. 2020. V. 15. № 3. P. e0230184.
Шлык Н.И. Сердечный ритм и тип регуляции у детей, подростков и спортсменов. Ижевск: “Удмуртский университет”, 2009. 259 с. Shlyk N.I. Heart rate and type of regulation in children, adolescents and athletes. Izhevsk: Udmurtskij universitet, 2009. 259 p.
Mokienko O.A., Chervyakov A.V., Kulikova S.N. et al. Increased motor cortex excitability during motor imagery in brain-computer interface trained subjects // Front. Comput. Neurosci. 2013. V. 7. P. 168.
Terkelsen A.J., Mölgaard H., Hansen J. et al. Heart rate variability in complex regional pain syndrome during rest and mental and orthostatic stress // Anesthesiology. 2012. V. 116. № 1. P. 133.
Ведясова О.А., Павленко С.И., Кретова И.Г., Комарова М.В. Влияние информационной нагрузки на динамику спектральных параметров вариабельности сердечного ритма у студентов с разными хронотипами // Рос. физиол. журн. им. И.М. Сеченова. 2016. Т. 102. № 8. С. 990. Vedyasova O.A., Pavlenko S.I., Kretova I.G., Komarova M.V. [The influence of information load on the timecourse of spectral parameters of heart rate variability in students with different chronotypes] // Ross. Fiziol. Zh. Im. I.M. Sechenova. 2016. V. 102. № 8. P. 990.
Андрианов В.В., Василюк Н.А., Бирюкова Е.В. Когнитивная деятельность студентов и ее гемодинамическое обеспечение // Вестн. новых мед. технологий. 2017. Т. 24. № 2. С. 130. Andrianov V.V., Vasilyuk N.A., Biryukova E.V. [Cognitive activity of the students and its haemodynamic ensuring] // Vest. Nov. Med. Tekhnol. 2017. V. 24. № 2. P. 130.
Джебраилова Т.Д., Коробейникова И.И., Каратыгин Н.А., Дудник Е.Н. Динамика параметров альфа-активности ЭЭГ и вариабельности сердечного ритма при интеллектуальной деятельности человека // Физиология человека. 2015. Т. 41. № 6. С. 36. Dzhebrailova T.D., Korobeinikova I.I., Karatygin N.A., Dudnik E.N. Dynamics of EEG α activity and heart rate variability in subjects performing cognitive tests // Human Physiology. 2015. V. 41. № 6. P. 599.
Павлов К.И., Мухин В.Н., Сырцев А.В. и др. Связь периодических модуляций сердечного ритма с уровнем активности сенсомоторной коры // Рос. физиол. журн. им. И.М. Сеченова. 2020. Т. 106. № 2. С. 205. Pavlov K.I., Mukhin V.N., Syrtsev A.V. et al. [Association between periodic modulations of the heart rate and sensomotor cortex activation] // Ross. Fiziol. Zh. Im. I.M. Sechenova. 2020. V. 106. № 2. P. 205.
Джебраилова Т.Д., Коробейникова И.И., Каратыгин Н.А. и др. Динамика спектральных характеристик ЭЭГ у лиц с разной личностной тревожностью при когнитивной деятельности // Физиология человека. 2021. Т. 47. № 1. С. 20. Dzhebrailova T.D., Korobeinikova I.I., Karatygin N.A. et al. Dynamics of EEG spectral characteristics in subjects with various trait anxiety levels performing cognitive tests // Human Physiology. 2021. V. 47. № 1. P. 14.
Pfurtscheller G., Solis-Escalante T., Barry R.J. Brisk heart rate and EEG changes during execution and withholding of cue-paced foot motor imagery // Front. Hum. Neurosci. 2013. V. 7. P. 379.
Баевский Р.М., Иванов Г.Г., Чирейкин Л.В. и др. Анализ вариабельности сердечного ритма при использовании различных электрокардиографических систем (методические рекомендации) // Вестник аритмологии. 2002. № 24. С. 65. Baevskij R.M., Ivanov G.G., Chirejkin L.V. et al. [Analiz variabel’nosti serdechnogo ritma pri ispol’zovanii razlichnykh elektrokardiograficheskikh sistem (metodicheskie rekomendatsii)] // Vestnik Aritmologii. 2002. № 24. P. 65.
Pelletier R., Higgins J., Bourbonnais D. Addressing neuroplastic changes in distributed areas of the nervous system associated with chronic musculoskeletal disorders // Phys. Ther. 2015. V. 95. № 11. P. 1582.
Desmurget M., Sirigu A. A parietal-premotor network for movement intention and motor awareness // Trends Cogn. Sci. 2009. V. 13. № 10. P. 411.
Côté S.L., Hamadjida X.A., Quessy X.S., Dancause X.N. Contrasting modulatory effects from the dorsal and ventral premotor cortex on primary motor cortex outputs // J. Neurosci. 2017. V. 37. № 24. P. 5960.
Kurgansky M.E., Bobrov P.D., Frolov A.A., Semenova E.I. Corticomuscular interactions in real and imaginary arm movements // Neurosci. Behav. Physiol. 2021. V. 51. № 6. P. 724.
Куликов В.Ю., Антропова Л.К., Козлова Л.А. Влияние функциональной асимметрии мозга на стратегию поведения индивида в стрессовой ситуации // Медицина и образование в Сибири. 2010. № 5. С. 10. Kulikov V.Y., Antropova L.K., Kozlova L.A. [Effect of functional brain asymmetry to adopt strategies of individual behavior in stressful situations] // Meditsina i Obrazovanie v Sibiri. 2010. № 5. P. 10.
Logue D.D., Logue R.T., Kaufmann W.E., Belcher H.M. Psychiatric disorders and left-handedness in children living in an urban environment // Laterality. 2015. V. 20. № 2. P. 249.
Моренова К.А., Ведясова О.А. Изменение паттерна электроэнцефалограммы у правшей и левшей при воображении движений // Ульяновский медико-биологический журн. 2019. № 1. С. 85. Morenova K.A., Vedyasova O.A. [Changes in electroencephalogram pattern in right- and left-handers during imaginary movements] // Ulyanovsk. Med. Biol. Zh. 2019. № 1. P. 85.
Gonzalez-Rosa J.J., Natali F., Tettamanti A. et al. Action observation and motor imagery in performance of complex movements: evidence from EEG and kinematics analysis // Behav. Brain Res. 2015. V. 281. P. 290.
Данилова Н.Н., Астафьев С.В. Внимание человека как специфическая связь ритмов ЭЭГ с волновыми модуляторами сердечного ритма // Журн. высш. нервн. деят. им. И.П. Павлова. 2000. Т. 50. № 5. С. 791. Danilova N.N., Astafiev S.V. [Human attention as a specific correlation between the EEG rhythms and activity of heart rate wave modulators] // Zh. Vyssh. Nerv. Deyat. Im. I.P. Pavlova. 2000. V. 50. № 5. P. 791.
Капустян Е.Г. Спектральные характеристики вариабельности сердечного ритма у женщин в послеродовом периоде в зависимости от латеральной конституции и гестационных асимметрий // Современные проблемы науки и образования. 2016. № 6. С. 11. Kapustyan E.G. [Spectral characteristics of heart rate variability with women in postnatal period depending on lateral constitution and gestational asymmetry] // Sovremennye Problemy Nauki i Obrazovaniya. 2016. № 6. P. 11.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека