

Физиология человека, 2022, T. 48, № 5, стр. 60-71
Площадь поперечного сечения мышечных волокон различного типа в скелетных мышцах спортсменов, тренирующих выносливость, силу и скоростно-силовые способности
С. С. Миссина 1, *, А. С. Крючков 1, М. А. Дикунец 1, Е. Б. Мякинченко 1
1 ФГБУ “Федеральный научный центр физической культуры и спорта”
Москва, Россия
* E-mail: missina.s.s@vniifk.ru
Поступила в редакцию 22.06.2021
После доработки 26.05.2022
Принята к публикации 27.05.2022
- EDN: OWMDPI
- DOI: 10.31857/S0131164622700060
Аннотация
Целью исследования являлось сравнение средней площади поперечного сечения мышечных волокон (ППСМВ) разных типов у спортсменов, тренирующих выносливость (СВ), силу (СС) и скоростно-силовые способности (ССС), по опубликованным данным. Был осуществлен поиск статей за период 1972–2021 гг. с данными ППСМВ у спортсменов-мужчин старше 18 лет. Отобрано 50 статей с результатами 807 испытуемых. Данные разделены на 4 группы. В первую группу включены статьи с ППСМВ у СВ. В этой группе данные по ППСМВ проанализированы отдельно для более нагружаемых и менее нагружаемых основных мышц, участвующих в упражнении. Во вторую и третью включены данные по наиболее нагружаемым мышцам у СС и ССС; в четвертую – контроль (К) для аналогичных мышц из тех же исследований. ППСМВ типа I и II у СВ в наиболее нагружаемых основных мышцах (6438 ± 1074 и 7296 ± 1242 мкм2) была выше (p < 0.05), чем в менее нагружаемых (5310 ± 1200 и 5800 ± 1100 мкм2). Значения ППСМВ типа I в наиболее нагружаемых мышцах были выше у СВ относительно СС и ССС (6438 ± 1074, 5715 ± 1090, 5690 ± 1010 мкм2, p < 0.05). ППСМВ типа II у СВ была ниже относительно СС (7296 ± 1242 и 8712 ± 1520 мкм2, p < 0.05), но не отличалась относительно ССС (6820 ± 1200 мкм2, p > 0.05). ППСМВ типа II, приведенная к массе тела, в наиболее нагружаемых основных мышцах была выше у СВ по сравнению с ССС (103.1 ± 20 и 84.0 ± 18 мкм2/кг, p < 0.05 соответственно) и не различалась от СС (97.3 ± 18 мкм2/кг, p > 0.05). Таким образом, адаптация мышц СВ к многолетней тренировке связана не только с повышением окислительных способностей, но и с гипертрофией МВ в наиболее нагружаемых основных мышцах. При сравнении ППСМВ у спортсменов разных специализаций необходимо учитывать функциональную нагрузку мышц в основном соревновательном упражнении.
Волокна скелетных мышц (МВ) обладают высокой способностью адаптироваться к физическим нагрузкам. Силовая тренировка приводит к значительному увеличению площади поперечного сечения МВ (ППСМВ) [1–3]. Такие же изменения, но менее выраженные, наблюдаются при тренировке скоростных и скоростно-силовых способностей спортсменов [4–6]. Тренировка выносливости приводит к значительному повышению окислительных способностей МВ [7–9]. В отношении ППСМВ при тренировке выносливости данные противоречивы. Например, показано, что в начале регулярной аэробной тренировки, при многолетней тренировке марафонцев или при повышении интенсивности аэробной тренировки, диаметр волокон – особенно медленных – снижается, а окислительные способности возрастают [10–15]. Однако, с другой стороны, имеются примеры высокой ППСМВ типов I и II у спортсменов в циклических видах спорта, тренирующих выносливость (СВ), относительно контроля и спортсменов других видов спорта. В частности, в отдельных работах показано, что абсолютные значения ППСМВ у высококвалифицированных спортсменов, несмотря на меньшую массу тела, оказываются на уровне или даже выше показателей ППСМВ аналогичных мышц у спортсменов, тренирующих силу и объем мышц (далее – “силовые виды спорта”), скоростно-силовые и спринтерские способности (далее – “скоростно-силовые виды спорта”) [8, 10, 16]. Кроме этого, найдено, что у одних СВ (например, велогонки, конькобежный спорт) ППСМВ может иметь относительно большие значения, в то время как у других СВ (например, бег) высокие значения ППСМВ могут не наблюдаться [17].
В этой связи, в настоящем исследовании проверялась гипотеза, что при длительной многолетней спортивной тренировке у СВ может наблюдаться не только повышение окислительных способностей МВ, но и увеличение ППСМВ. Однако эффект увеличения ППСМВ будет проявляться, главным образом, в мышцах, вносящих основной вклад в создание моментов силы в суставах конечностей и/или испытывающих относительно высокие пиковые механические нагрузки при взаимодействии с опорой или инвентарем во время длительной (многолетней) тренировки.
Целью настоящей работы являлось сравнение опубликованныx в литературе данныx ППСМВ типов I и II у СВ средней и высокой квалификации в мышцах, несущих различную механическую нагрузку при спортивной тренировке, с данными по ППСМВ спортсменов силовых, скоростно-силовых видов спорта и нетренированных лиц.
МЕТОДИКА
Для сравнения ППСМВ типа I и II у спортсменов разных видов спорта был проведен поиск и анализ экспериментальных данных, опубликованных в период с 1972 по 2021 гг. [1–10, 12–16, 18–55 ].
Поиск осуществляли в базах данных PubMed и Google Scholar по ключевым словам “cross sectional area”, “fiber composition”, “fiber size”, “skeletal muscle fiber types”, “athletes”. Во внимание принимали только данные спортсменов мужчин в возрасте 18–33 лет, в отношении которых можно было сделать вывод, что исследовали спортсменов “средней и/или высокой квалификации”. Оценку проводили по следующим критериям: значения максимального потребления кислорода (МПК) (60–79.9 мл/кг/мин) (для СВ), и/или объем тренировочной нагрузки (например, выше 10 ч в неделю, и/или стажа непрерывных профессиональных занятий спортом в момент исследования (например, более 5 лет), и/или спортивная квалификация (например, спортсмены регионального, национального или международного уровня). В результате поиска были найдены 103 статьи, из них были исключены 53 работы, не соответствующие требуемым критериям: 1) 20 исследований с данными спортсменов низкой квалификации и без деления по специализациям; 2) 29 статей, в которых публиковались результаты оценки ППСМВ в процентных соотношениях площадей, занимаемых волокнами на поперечном срезе без указания размеров МВ; 3) 3 статьи – спортсмены старше 33 лет; 4) 1 статья – спортсмены, тренирующиеся в сложно-координационном виде спорта (прыжки с вышки). В итоге былo отобранo 50 статей, в которых в сумме представлены данные по ППСМВ 807 спортсменов и лиц, отнесенных к контрольной группе.
Полученные данные были разделены на 4 группы. Первая группа включала статьи с данными по ППСМВ у СВ. В этой группе были определены мышцы, несущие наиболее высокую механическую нагрузку (далее – “наиболее нагружаемые” мышцы), из числа тех мышц, которые подвергались гистохимическому анализу, и, напротив, мышцы конечностей, которые активны в данной локомоции, но несущие меньшую нагрузку, были обозначены как “менее нагружаемые” мышцы. К категории “наиболее нагружаемые” мышцы были отнесены: m. gastrocnemius (mGS) – у бегунов и триатлетов, m. lateral vastus (mVL) – у велосипедистов, конькобежцев и триатлетов; m. deltoideus – у пловцов, гребцов-каноистов и каякеров. У лыжников данные были распределены следующим образом: ППСМВ mVL из статей, опубликованных ранее 1985 г., т.е. до появления конькового хода, в котором mVL не несет высокую механическую нагрузку, были отнесены к категории “менее нагружаемые” мышцы. И, напротив, данные для той же мышцы из статей после 1985 г. были отнесены к группе “наиболее нагружаемые”, так как в коньковом лыжном ходе mVL играет существенную роль. Кроме этого, к категории “наиболее нагружаемые” мышцы у лыжников была отнесена m. triceps brachii (mTB), как несущая высокую нагрузку при отталкивании руками. Во вторую группу – силовые виды спорта – вошли данные для “наиболее нагружаемых” (т.е. несущих основную нагрузку и, соответственно, тренируемых) мышц спортсменов, специализирующихся в тяжелой атлетике, пауэрлифтинге и бодибилдинге. В третью – скоростно-силовые виды спорта – данные спринтеров, прыгунов, метателей, футболистов, волейболистов, хоккеистов и борцов. Четвертая группа включала данные нетренированных мужчин из тех же статей для исключения возможного влияния фактора различий в методике определения ППСМВ.
Из каждой статьи брали средние значения ППСМВ по каждому подтипу МВ отдельно по видам спорта. Выделяли четыре подгруппы типов МВ, а именно: I, II (без подразделения на подтипы), IIA и IIХ. К типу IIX были отнесены все подтипы, которые авторы классифицировали как IIAB/IIB/IIX/IIC/IIAC и др. Данные тех статей, в которых были представлены значения диаметров МВ, пересчитывали в значения ППСМВ. В работах, где оценивали изменение ППСМВ до и после экспериментального периода тренировки (не более нескольких месяцев), данные ППСМВ усреднялись. Число наблюдений (спортсменов), характеристики спортсменов и значения ППСМВ для разных мышц из каждой статьи представлены в табл. 1–5 для соответствующих категорий испытуемых.
Таблица 1.
Сводные данные по средним значениям МПК, возраста, длины и массы тела, площади поперечного сечения для мышечных волокон различных типов у контрольной группы
| Мышца | МПК, мл/кг/мин | n | Возраст, лет | Длина тела, см | Масса тела, кг | Площадь поперечного сечения мышечного волокна, мкм2 × 103 | Источник | |||
|---|---|---|---|---|---|---|---|---|---|---|
| тип I | тип II | тип IIА | тип IIХ | |||||||
| mGS | – | 11 | 27 | 177 | 78.2 | 5.70 | 4.97 | – | – | [4] |
| mGS | – | 8 | 25 | 176.9 | 76.8 | 4.72 ± 0.4 | 4.71 ± 0.3 | – | – | [5] |
| mVL | 54.2 | 9 | 23 | – | 72 | 5.29 ± 0.6 | 5.43 ± 0.4 | 5.57 ± 0.5 | 5.15 ± 0.3 | [6] |
| mVL | – | 10 | 23.3 ± 2 | – | – | 4.67 ± 0.6 | 5.04 ± 0.7 | 5.28 ± 0.6 | 4.65 | [8] |
| mVL | 49.8 | 6 | 25 | – | 80 | 3.80 ± 0.1 | 4.76 ± 0.1 | 5.11 ± 0.1 | 4.11 | [9] |
| mVL | 42.9 | 4 | 24–30 | 182 | 74.5 | 4.28 ± 0.3 | 5.22 ± 0.3 | – | – | [16] |
| m. delt | – | 1 | – | – | – | 3.23 ± 0.2 | 3.59 ± 0.2 | – | – | |
| mVL | 43.5 | 8 | 23–32 | – | 70.5 | 4.56 ± 1.8 | 5.07 ± 1.7 | – | – | [18] |
| mVL | – | 5 | – | – | 3.30 ± 0.9 | 3.83 ± 1.0 | 4.11 ± 0.2 | 3.42 ± 0.2 | [19] | |
| mGS | 38.4 | 19 | 27.4 | 178.4 | 82.3 | 5.46 ± 0.6 | 4.95 ± 0.5 | – | – | [20] |
| mVL | 38.2 | 19 | 27.4 | 178.4 | 82.3 | 5.50 ± 0.6 | 4.95 ± 0.5 | – | – | [21] |
| mVL | 41.4 | 5 | 29 | 180 | 83 | 4.73 ± 0.9 | 6.48 ± 1.8 | 6.86 ± 1.8 | 6.18 ± 0.1 | [22] |
| mVL | – | 12 | 21.6 ± 4.5 | 181.7 | 74.5 | 5.71 ± 1.4 | 6.49 ± 1.5 | – | – | [23] |
| m. delt | – | 12 | – | – | – | 4.66 ± 1.0 | 5.98 ± 1.4 | – | – | |
| mVL | 45.6 | 12 | 21.7 ± 0.8 | 179 | 76 | 3.30 ± 0.3 | 4.79 ± 0.1 | 5.21 ± 0.5 | 4.30 ± 0.6 | [24] |
| mVL | – | 7 | 30 ± 1 | 180 ± 6 | 77 ± 19 | 4.04 ± 0.9 | 4.28 ± 0.9 | – | – | [25] |
| mBB | – | 4 | 28 ± 0.4 | 181 ± 4 | 75 ± 2 | 5.52 ± 0.5 | 6.74 ± 0.3 | 7.29 ± 0.2 | 6.21 ± 0.3 | [26] |
| mVL | – | 8 | 20.6 ± 0.9 | – | – | 5.54 ± 1.2 | 6.76 ± 1.0 | 6.98 ± 1.3 | 6.43 ± 0.6 | [27] |
| mBB | – | 2 | 27.5 | 173.5 | 76.7 | 4.08 ± 0.2 | 4.54 ± 0.3 | – | – | [28] |
| mVL | – | 10 | 22.9 ± 0.9 | 180 ± 1 | 72.3 ± 2 | 4.57 ± 0.3 | 5.04 ± 0.3 | – | – | [29] |
| m. trapez | – | 6 | 23.3 ± 3 | – | – | 4.17 ± 1.0 | 4.54 ± 0.7 | 4.63 ± 0.6 | 4.40 ± 0.7 | [30] |
| Х ± SD | 44.3 ± 5 | 9 ± 5 | 25.4 ± 3 | 179 ± 2 | 76.7 ± 4 | 4.61 ± 0.8 | 5.15 ± 0.9 | 5.67 ± 1.1 | 4.98 ± 1.1 | |
Таблица 2.
Сводные данные по средним значениям МПК, возраста, длины и массы тела, площади поперечного сечения для мышечных волокон различных типов более нагружаемых мышц у спортсменов циклических видов спорта
| Специализация спортсменов | Мышца | МПК, мл/кг/мин | n | Возраст, лет | Длина тела, см | Масса тела, кг | Площадь поперечного сечения мышечного волокна, мкм2 × 103 | Источник | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| тип I | тип II | тип IIА | тип IIХ | ||||||||
| Бег | mGS | – | 7 | 19.9 | 179 | 66 | 6.10 | 7.12 | – | – | [4] |
| – | 5 | 24.2 | 180 | 71 | 6.61 | 7.63 | – | – | |||
| Велоспорт (шоссе) | mVL | – | 20 | 23 | – | – | 5.89 ± 0.7 | 5.71 ± 0.8 | 5.74 ± 0.8 | 5.67 ± 0.8 | [8] |
| Бег | mGS | 71 ± 1 | 8 | 21 | 177 | 65 | 7.73 ± 0.3 | 8.54 ± 0.5 | – | – | [10] |
| Плавание | m. delt | 79.9 ± 1 | 5 | 21 | 181 | 78.3 | 6.12 ± 0.2 | 6.65 ± 0.3 | – | – | [16] |
| Велоспорт | mVL | 68.2 | 4 | 24 | 182 | 74.5 | 8.65 ± 0.8 | 9.95 ± 0.4 | – | – | |
| Бег | mGS | 77.4 | 14 | 26.2 | 178.9 | 63.9 | 6.49 ± 0.7 | 8.34 ± 0.7 | – | – | [20] |
| 70.1 | 18 | 25.3 | 180 | 68 | 6.28 ± 0.4 | 6.38 ± 0.4 | – | – | |||
| Велоспорт (шоссе) | mVL | 67.1 | 11 | 24.6 | 180 | 72.8 | 6.33 ± 0.4 | 6.12 ± 0.4 | – | – | [21] |
| Гребля на каяках | m. delt | – | 9 | 23.2 | 185 | 81 | 4.92 ± 1.3 | 7.04 ± 2.6 | – | – | [24] |
| Лыжные гонки | mTB | 76.3 ± 2 | 6 | 19 ± 1 | – | 71.3 | 6.23 ± 0.5 | 8.48 ± 0.7 | 8.48 ± 0.7 | – | [31] |
| Лыжные гонки (конек) | mVL | 6.05 ± 0.3 | 6.85 ± 0.4 | 6.85 ± 0.4 | – | ||||||
| Лыжные гонки | mTB | 69 | 10 | 22 ± 1 | 181 | 79 | 5.36 ± 0.2 | 8.11 ± 0.4 | 8.10 ± 0.4 | 5.36 ± 0.2 | [32] |
| Лыжные гонки | mVL | 5.42 ± 0.3 | 6.80 ± 0.3 | 6.81 ± 0.3 | 6.22 ± 0.5 | ||||||
| Спортивная ходьба | mGS | – | 10 | 27 | 171 | 66.7 | 7.71 ± 0.8 | 8.20 ± 1.9 | – | – | [33] |
| Бег | mGS | – | 6 | 20 | 176 | 63 | 7.63 ± 1.8 | 9.11 ± 0.3 | – | – | |
| Бег | mGS | 68 | 8 | 27 ± 3 | 178 ± 2 | 68.8 ± 5 | 4.66 ± 0.7 | 5.76 ± 1.7 | – | – | [34] |
| Велоспорт (шоссе) | mVL | 71 | 16 | 26 | 178 | 71 | 6.20 | 6.19 | 6.16 | 6.2 | [35] |
| Бег | mGS | 70.5 | 5 | 20 ± 1 | 178 ± 3 | 65 ± 4 | 7.34 ± 0.8 | 7.69 ± 0.5 | 7.69 ± 0.5 | [36] | |
| Лыжные гонки | mTB | 75.4 | 10 | 19 ± 4 | 183 ± 6 | 74 ± 6 | 5.68 ± 1.2 | 8.56 ± 3.1 | 8.98 ± 2.2 | 5.55 ± 0.4 | [37] |
| Лыжные гонки (конек) | mVL | 70.5 | 5 | 20 ± 1 | 178 ± 3 | 65 ± 4 | 6.07 ± 1.2 | 6.22 ± 0.9 | 6.28 ± 1.1 | 5.52 ± 0.7 | |
| Конькобежцы | mVL | – – |
20 14 |
18.9 | 181 ± 5 181 |
74.5 ± 3.8 | 5.63 ± 1.1 | 6.02 ± 1.7 | – | – | [38] |
| Велоспорт | mVL | 19 ± 1 | 71 | 5.76 ± 0.9 | 5.37 ± 1.0 | – | – | [39] | |||
| Велоспорт, триатлон | mVL | 63.2 ± 2 | 14 | 25 ± 6 | 182 | 76.5 | 5.94 ± 1.3 | 5.97 ± 1.3 | – | – | [40] |
| Лыжные гонки | mTB | 66 ± 3 | 8 | 24 ± 4 | – | 79 ± 7 | 7.62 ± 2.6 | 8.81 ± 2.1 | – | – | [41] |
| mVL | 8.97 ± 1.9 | 8.21 ± 1.4 | |||||||||
| Х ± SD | 70.4 ± 5 | 10 ± 5 | 22.7 ± 3 | 180 ± 3 | 71.8 ± 5 | 6.44 ± 1.1 | 7.30 ± 1.2 | 7.23 ± 1.1 | 5.75 ± 0.4 | ||
Примечание: mTB – m. Triceps Bbrachii. Остальные обозначения см. табл. 1.
Таблица 3.
Сводные данные по средним значениям МПК, возраста, длины и массы тела, площади поперечного сечения для мышечных волокон различных типов менее нагружаемых мышц у спортсменов циклических видов спорта
| Специализация спортсменов | Мышца | МПК, мл/кг/мин | n | Возраст, лет | Длина тела, см | Масса тела, кг | Площадь поперечного сечения мышечного волокна, мкм2 × 103 | Источник | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| тип I | тип II | тип IIА | тип IIХ | ||||||||
| Лыжные гонки, бегуны, велоспорт | mVL | 77.9 | 6 | 24 | – | 73 | 4.6 ± 0.1 | 5.13 ± 0.2 | 5.34 ± 0.2 | 4.81 ± 0.2 | [9] |
| Бег | mVL | 68.5 | 26 | 22 | 177 | 64 | 5.45 ± 1.6 | 6.48 ± 2.0 | – | – | [12] |
| Бег | mVL | 67 ± 4 | 18 | – | 176 | 64 | 4.29 ± 1.0 | 5.07 ± 1.3 | – | – | [13] |
| Велоспорт | m. delt | 68.2 ± 2 | 4 | 24 | 182 | 74.5 | 5.47 ± 0.2 | 7.34 ± 0.3 | – | – | [16] |
| Бег | mVL | 72.4 ± 2 | 8 | 23 | 177 | 69.5 | 4.93 ± 0.4 | 5.56 ± 0.3 | – | – | |
| Плавание | mVL | 79.9 ± 1 | 5 | 21 | 181 | 78.3 | 4.95 ± 0.2 | 6.93 ± 0.3 | – | – | |
| Лыжные гонки, бег, велоспорт | mVL | 64.8 ± 7.1 | 6 | 29.7 ± 6 | – | 69.1 ± 5 | 4.57 ± 1.5 | 5.07 ± 1.9 | – | – | [18] |
| Бег | mVL | – | 3 | – | – | – | 4.61 ± 1.2 | 4.77 ± 1.2 | 5.22 ± 0.2 | 3.67 | [19] |
| Бег | mVL | 60 | 8 | 28 | 174 | 65 | 4.94 ± 1.1 | 4.59 ± 1.3 | 4.56 ± 1.3 | 4.67 ± 1.8 | [22] |
| Бег | mVL | – | 9 | 25 ± 5 | 180.2 | 68.1 | 5.80 ± 1.4 | 5.85 ± 1.8 | – | – | [23] |
| m. delt | 4.64 ± 1.3 | 5.60 ± 1.2 | – | – | |||||||
| Гребля на каяках | mVL | 9 | 23 ± 4 | 184.7 | 80.9 | 5.17 ± 1.2 | 7.43 ± 1.2 | – | – | ||
| Бег | mVL | – | 6 | 26 | 177 | 66.5 | 4.30 ± 0.5 | 4.85 ± 0.5 | – | – | [29] |
| Спортивная ходьба | mVL | – | 10 | 27 | 171 | 66.7 | 6.98 ± 1.7 | 6.07 ± 1.4 | – | – | [33] |
| Бег | mVL | – | 6 | 20 | 176 | 63 | 9.00 ± 2.5 | 8.68 ± 2.6 | – | – | |
| Бег | mVL | 68.5 | 23 | 21 | 170 | 59.5 | 6.82 ± 2.4 | 6.25 ± 2.3 | – | – | [42] |
| Плавание (длинные дистанции) | mVL | – | 8 | 18 | 184 | 71 | 4.20 ± 0.1 | 4.92 ± 0.2 | – | – | [43] |
| Плавание (средние дистанции) | mVL | – – |
7 | 17 | 178 | 74 | 4.41 ± 0.6 | 5.16 ± 0.9 | – | – | |
| Бег | mVL | 7 | 22 | – | – | 6.29 ± 0.2 | 5.67 ± 0.1 | – | – | [44] | |
| Бег (марафон) | mVL | – | 5 | 29.6 ± 4 | – | – | 4.78 ± 1.6 | 4.50 ± 1.4 | 4.48 ± 1.4 | 4.59 ± 1.4 | [45] |
| Х ± SD | 70.3 ± 6 | 9 ± 6 | 23 ± 4 | 178.1 ± 4 | 69 ± 6 | 5.31 ± 1.2 | 5.80 ± 1.1 | 4.90 ± 0.4 | 4.44 ± 0.5 | ||
Примечание: обозначения см. табл. 1.
Таблица 4.
Сводные данные по средним значениям МПК, возраста, длины и массы тела, площади поперечного сечения для мышечных волокон различных типов у спортсменов силовых видов спорта
| Специализация спортсменов | Мышца | МПК, мл/кг/мин | n | Возраст, лет | Длина тела, см | Масса тела, кг | Площадь поперечного сечения мышечного волокна, мкм2 × 103 | Источник | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| тип I | тип II | тип IIА | тип IIХ | ||||||||
| Бодибилдинг, пауэрлифтинг | mTB | – | 7 | 26.4 | – | 98 | 4.77 ± 1.1 | 7.87 ± 1.1 | – | – | [1] |
| Бодибилдинг | mVL | – | 3 | 25 | 177 | 84 | 5.40 | 7.10 | – | – | [2] |
| m. delt | – | 4.40 | 6.70 | – | – | ||||||
| Бодибилдинг | mBB | – | 8 | 33 | 173 | 95 | 7.53 ± 0.6 | 11.40 ± 1.0 | – | – | [3] |
| Тяжелая атлетика | mVL | 47.9 ± 6.4 | 8 | 20.3 ± 4 | – | 72.8 ± 18 | 4.55 ± 0.1 | 7.43 ± 0.2 | – | – | [18] |
| Тяжелая атлетика | mVL | – | 4 | – | – | – | 4.43 ± 1.3 | 7.06 ± 1.1 | 7.29 ± 0.3 | 7.02 ± 0.2 | [19] |
| Тяжелая атлетика | m. delt | 40.1 ± 6.1 | 4 | 25 | 171 | 81.3 | 6.04 ± 0.6 | 9.74 ± 0.5 | [21] | ||
| Тяжелая атлетика | mVL | 43.7 | 7 | 24 | 174 | 86 | 5.29 ± 0.6 | 8.27 ± 1.9 | 9.11 ± 2.1 | 6.74 ± 0.6 | [22] |
| Тяжелая атлетика | mVL | – | 7 | 25 | 172.4 | 86.6 | 5.06 ± 0.8 | 8.91 ± 1.8 | – | – | [23] |
| Тяжелая атлетика | m.delt | – | 5.01 ± 0.7 | 8.45 ± 1.3 | – | – | |||||
| Бодибилдинг, пауэрлифтинг | mVL | – | 13 | 26 | 180 | 95 | 6.70 ± 1.5 | 9.85 ± 2.0 | – | – | [25] |
| Бодибилдинг | mBB | – | 4 | 25 ± 2 | 185 ± 2 | 98 ± 3 | 6.83 ± 1.0 | 10.57 ± 1.1 | 11.0 ± 1.0 | 9.83 ± 1.3 | [26] |
| Бодибилдинг | mBB | – | 5 | 32.8 ± 2 | 171.3 ± 2 | 89.6 ± 2 | 8.17 ± 0.6 | 12.50 ± 1.3 | – | – | [28] |
| Пауэрлифтинг | m. trap | – | 10 | 27.7 ± 8 | – | – | 5.70 ± 0.9 | 8.04 ± 1.8 | 8.00 ± 2.1 | 8.24 ± 1.4 | [30] |
| Тяжелая атлетика | mVL | – | 6 | 27 ± 2 | 174 | 95 | 6.33 ± 0.7 | 8.42 ± 0.7 | 8.74 ± 0.7 | 2.65 ± 1.6 | [46] |
| Пауэрлифтинг | mVL | – | 10 | 27 ± 7 | – | 98 ± 21 | 4.77 ± 1.2 | 7.28 ± 1.2 | 7.31 ± 1.3 | 7.23 ± 0.9 | [47] |
| Бодибилдинг | mVL | – | 8 | 26.1 ± 4.2 | 178 ± 5 | 91 ± 6 | 6.27 ± 1.5 | 8.77 ± 1.5 | 9.64 ± 1.7 | 7.62 ± 1.3 | [48] |
| Тяжелая атлетика | mVL | 32.6 | 1 | 22 | – | 121.5 | 5.63 ± 1.6 | 8.78 ± 1.5 | 9.62 ± 1.3 | 8.54 ± 1.8 | [49] |
| Х ± SD | 41.1 ± 7 | 6 ± 3 | 26 ± 3 | 175 ± 4 | 91 ± 11 | 5.72 ± 1.1 | 8.71 ± 1.5 | 8.84 ± 1.3 | 7.22 ± 2.1 | ||
Примечание: mTB – m. Triceps Brachii. Остальные обозначения см. табл. 1.
Таблица 5.
Сводные данные по средним значениям МПК, возраста, длины и массы тела, площади поперечного сечения для мышечных волокон различных типов у спортсменов скоростно-силовых видов спорта
| Специализация спортсменов | Мышца | МПК, мл/кг/мин | n | Возраст, лет | Длина тела, см | Масса тела, кг | Площадь поперечного сечения мышечного волокна, мкм2 × 103 | Источник | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| тип I | тип II | тип IIА | тип IIХ | ||||||||
| Легкая атлетика (спринт) | mGS | – | 2 | 19.5 | 181 | 71.5 | 5.88 | 6.03 | – | – | [4] |
| Прыжки в длину | mGS | – | 2 | 29 | 183 | 77 | 4.72 | 6.52 | – | – | |
| Метание копья | mGS | – | 3 | 25.3 | 176 | 83.6 | 5.59 | 5.77 | – | – | |
| Толкание ядра | mGS | – | 4 | 26.8 | 198 | 129.0 | 7.70 | 9.48 | – | – | |
| Толкание ядра | mGS | – | 8 | 25 ± 2 | 191 | 125 | 6.37 ± 0.5 | 6.44 ± 0.7 | – | – | [5] |
| Хоккей | mVL | 55.3 ± 1 | 12 | 21 | – | 77 | 5.56 ± 0.3 | 6.44 ± 0.5 | 6.50 ± 0.4 | 6.12 ± 0.5 | [6] |
| Борьба | mVL | – | 8 | 22.1 ± 4 | 173 | 75 | 5.70 ± 2.1 | 7.01 ± 2.0 | – | – | [23] |
| m. delt | – | 4.50 ± 1.0 | 7.13 ± 2.0 | – | – | ||||||
| Велоспорт, спринт | mVL | 65 | 10 | 22 | 177 | 76 | 7.04 ± 3.4 | 8.25 ± 1.3 | 9.34 ± 0.8 | 7.3 ± 0.8 | [24] |
| Футбол | mVL | – | 11 | 18.1 ± 1 | – | – | 6.29 ± 1.6 | 7.78 ± 1.9 | 7.92 ± 2.0 | 7.47 ± 1.9 | [27] |
| Волейбол | mVL | – | 10 | 21.4 | 188 | 81 | 4.56 ± 0.3 | 5.89 ± 0.5 | – | – | [29] |
| Легкая атлетика (спринт) | mVL | – | 7 | 24 | 183 | 74.9 | 6.50 ± 1.1 | 6.16 ± 2.0 | – | – | [33] |
| mGS | – | 7.01 ± 1.0 | 7.65 ± 1.5 | – | – | ||||||
| Прыжки в высоту | mVL | – | 20 | 22 | 183 | 67 | 7.14 ± 2.0 | 7.74 ± 1.5 | – | – | |
| mGS | – | 6.46 ± 2.5 | 7.70 ± 2.8 | – | – | ||||||
| Плавание (спринт) | mVL | – | 5 | 19.2 ± 1 | 181 ± 3 | 74 ± 4 | 4.48 ± 0.5 | 5.64 ± 0.6 | – | – | [43] |
| Легкая атлетика (спринт) | mVL | – | 12 | 22 | – | – | 5.38 ± 0.2 | 5.61 ± 0.1 | – | – | [44] |
| Легкая атлетика (спринт) | mVL | – | 5 | 23.6 ± 2 | – | – | 5.00 ± 2.7 | 6.78 ± 3.0 | 7.31 ± 3.1 | 5.94 ± 3.0 | [45] |
| Плавание (спринт) | m. delt | 62.2 | 12 | 19 | 180.3 | 74.2 | 5.15 ± 0.1 | 6.64 ± 0.1 | – | – | [50] |
| Толкание ядра | mVL | – | 2 | – | – | – | 4.68 ± 0.3 | 7.27 ± 1.2 | – | – | [51] |
| Легкая атлетика (спринт) | mVL | – | 16 | 24.3 ± 1 | 178 | 77 | 4.30 ± 0.2 | 4.70 ± 0.1 | 4.70 ± 0.1 | 3.45 ± 0.4 | [52] |
| Футбол | mVL | – | 14 | 25 | 179 | 75 | 4.86 ± 0.4 | 5.53 ± 0.5 | 5.67 ± 0.4 | 5.12 ± 1.1 | [53] |
| Метание молота | mVL | – | 6 | 26 | 185 | 116 | 5.79 ± 0.7 | 7.53 ± 1.5 | 7.70 ± 1.2 | 6.56 ± 2.0 | [54] |
| Легкая атлетика (спринт) | mVL | – | 1 | – | – | – | 6.01 ± 1.0 | 5.79 ± 0.6 | 6.25 ± 0.6 | 5.30 ± 1.0 | [55] |
| Х ± SD | 60.8 ± 5 | 8 ± 5 | 23 ± 3 | 182 ± 6 | 83 ± 19 | 5.69 ± 1.0 | 6.82 ± 1.2 | 6.92 ± 1.5 | 5.91 ± 1.3 | ||
Примечание: обозначения см. табл. 1.
Для каждой группы рассчитывали средние и стандартные отклонения ППСМВ каждого подтипа по средним значениям ППСМВ из каждой работы. Выборки каждой группы были проверены на нормальное распределение по критерию Колмогорова-Смирнова. В связи с ненормальностью распределения для большинства групп, статистическую значимость различий средних рассчитывали по непараметрическому критерию Манна-Уитни при уровне значимости p < 0.05, с использованием статистического пакета StatPlus (Analyst Soft Inc., США) версия 7. Поскольку изучаемые категории испытуемых могли значительно различаться по массе тела, что может влиять на результаты сопоставления ППСМВ между этими группами, отдельно, только для тех работ, где были опубликованы антропометрические данные, значения ППСМВ были приведены к массе тела. Для сравнения ППСМВ “наиболее нагружаемой” мышцы (mGS) с “менее нагружаемой” (mVL) у одних и тех же СВ (бег на длинные и средние дистанции, спортивная ходьба) дополнительно были выбраны и проанализированы соответствующие данные из 14 статей.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В табл. 1–5 представлены средние данные ППСМВ для исследований, включенных в анализ ППСМВ. Пустые ячейки указывают на отсутствие информации.
У спортсменов силовых видов спорта значения ППСМВ типа II и подтипа IIА выше относительно аналогичных показателей для спортсменов скоростно-силовых видов спорта (p < 0.001 и p = = 0.043) и для “наиболее нагружаемых” мышц у СВ (p = 0.007 и p = 0.038); ППСМВ типа IIХ выше у СС, чем в “наиболее нагружаемых” мышцах СВ (p = 0.042). Не было отличий в ППСМВ типов II, IIА и IIХ у спортсменов скоростно-силовых видов спорта и СВ в “наиболее нагружаемых” мышцах (p > 0.05). ППСМВ МВ типа I “наиболее нагружаемых” мышц у СВ выше относительно силовых видов спорта (р = 0.038), скоростно-силовых видов спорта (p = 0.027), контроля (p < < 0.001) и “менее нагружаемых” мышц у СВ (p = = 0.001). В “наиболее нагружаемых” мышцах ППСМВ типа IIA у СВ были выше типа IIX (p = = 0.009). У СВ ППСМВ всех подтипов (IIA и IIX) были выше в “наиболее нагружаемых” мышцах, чем в “менее нагружаемых” мышцах (p = 0.019 и р = 0.367 соответственно).
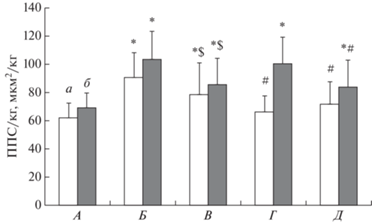
На рис. 1 представлены средние значения ППСМВ, приведенные к массе тела. Относительные значения ППСМВ типа I в “наиболее нагружаемых” мышцах у СВ были выше относительно ППСМВ в силовых и скоростно-силовых видах спорта (89.2 ± 18 против 63.9 ± 13 и 71.7 ± ± 17 мкм2/кг, соответственно, p < 0.05). Относительная ППСМВ типа II “наиболее нагружаемых” мышц у СВ была выше относительно скоростно-силовых видов спорта (102.7 ± 21 против 84.0 ± 18 мкм2/кг, p < 0.05), но не различалась относительно силовых видов спорта (97.3 ± 18 мкм2/кг, p > 0.05).
Рис. 1.
Средние значения площади поперечного сечения мышечных волокон (ППСМВ) типов I (а) и II (б), приведенные к массе тела у спортсменов различных групп видов спорта и контроля. А – контроль, все мышцы, Б – наиболее нагружаемые мышцы у спортсменов, тренирующих выносливость (СВ), В – менее нагружаемые мышцы у СВ, Г и Д – наиболее нагружаемые мышцы у спортсменов, соответственно силовых (Г) и скоростно-силовых (Д) видов спорта. Различия достоверны (p < 0.05): * – отличия от контрольной группы; # – отличия от наиболее нагружаемых мышц СВ; $ – отличия наиболее нагружаемых мышц от менее нагружаемых у СВ.
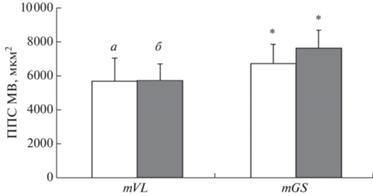
На рис. 2 представлены значения ППСМВ обоих типов в мышцах mVL и в mGS у бегунов. Значения ППСМВ типа I (р = 0.02) и типа II (р = = 0.007) были выше в mGS относительно mVL.
Рис. 2.
Средние значения площади поперечного сечения мышечных волокон (ППСМВ) в musculus vastus lateralis (mVL, n = 12) и musculus gastrocnemius (mGS, n = 9) у высококвалифицированных легкоатлетов. а – Тип I, б – тип II. n – число исследований, из которых взяты средние значения ППСМВ. Различия достоверны при p < 0.05. * – отличие от mVL при p < 0.05.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
По опубликованным в литературе данным в исследовании сравнивалась ППСМВ разного типа у спортсменов, тренирующих выносливость, силовые, скоростно-силовые способности, и нетренированных лиц.
Было показано, что ППСМВ типа I в “наиболее нагружаемых” мышцах у спортсменов, тренирующих выносливость (6.44 ± 1.1 мкм2 × 103) выше, чем ППСМВ этого типа у спортсменов силовых (5.72 ± 1.1 мкм2 × 103), скоростно-силовых (5.69 ± 1.0 мкм2 × 103) видов спорта и контроля (4.61 ± 0.8 мкм2 × 103). Особенно заметными различия становятся, если привести ППСМВ к массе тела (рис. 1). Более того, ППСМВ волокон типа II, приведенная к массе тела у спортсменов, тренирующих выносливость, также оказалась высокой: не различалась с ППСМВ этого типа у спортсменов, тренирующих силу, и была выше, чем у спортсменов скоростно-силовых видов спорта (рис. 1).
Таким образом, полученные нами результаты не согласуются с популярной в литературе точкой зрения, что СВ должны иметь относительно небольшую ППСМВ спортсменов других специализаций, имеющих такую же массу тела. Вывод о целесообразности относительно низкой ППСМВ у спортсменов, тренирующих выносливость, основан: на данных об обратной зависимости между ППСМВ и их окислительным потенциалом у млекопитающих, включая человека [56, 57]; на низких значениях ППСМВ, экспериментально подтвержденных в некоторых исследованиях [13, 14]; на модели A. Krogh, согласно которой небольшой диаметр МВ облегчает диффузию кислорода к потребителям внутри МВ [58]. Однако, также неоднократно высказывались и другие гипотезы о механизмах и факторах адекватного снабжения МВ кислородом, из которых нельзя сделать заключение о необходимости небольшого диаметра МВ [59, 60].
Обсуждая причины большого размера МВ обоих типов у спортсменов, тренирующих выносливость, можно предположить, что к увеличению размера приводит длительная нагрузка, которую определенные мышцы испытывают при многолетнем использовании определенной локомоции в качестве основного тренировочного и соревновательного упражнений. В этой логике хорошо объясняется высокая относительная ППСМВ типа I. Действительно, в силу “правила размера” E. Henneman [61], медленно сокращающиеся МВ рекрутируются уже при относительно низкой степени напряжения мышц и, по этой причине, могут играть основную роль в генерировании силы при выполнении длительных циклических (т. н. “аэробных”) упражнений. Кроме того, упражнения с такой интенсивностью составляют 70–90% от всего тренировочного объема у большинства СВ и, в среднем, составляют 2–2.5 ч в день, включая дни отдыха и отпуск [62, 63]. Следовательно, факторы активации синтеза структур органелл мышечных клеток, связанные с длительностью активного состояния и/или механической нагрузкой на МВ типа I при выполнении самой локомоции, могут быть причиной их относительно высокой гипертрофии. Кроме этого, данные настоящей работы позволяют считать, что те же факторы в полной мере действуют в отношении МВ типа IIA, поэтому в “наиболее нагружаемых” мышцах СВ ППСМВ была существенно выше, чем МВ типа IIX тех же мышц. Следовательно, если предположение о влиянии больших объемов циклической работы, как главной причины увеличения ППСМВ справедливо, то МВ типа IIA оказываются объектом тренировочного (механического) воздействия у СВ в не меньшей степени, чем МВ типа I.
Однако объяснить данные ряда исследований [9, 13, 16, 18, 19, 22, 23, 29, 43, 45] (табл. 3), в которых получены относительно небольшая ППСМВ у СВ, относительно ППСВМ у СВ, полученных в других исследованиях [4, 10, 16, 33, 36, 41] (табл. 2), на наш взгляд, удается, если оценивать ППСМВ, разделив основные мышцы, участвующие в локомоции, на “наиболее” и “менее нагружаемые” по механической нагрузке. Действительно, нами получена существенная разница ППСМВ всех типов (I, II, IIA и IIX) между “наиболее нагружаемыми” и “менее нагружаемыми” мышцами у СВ (15, 21, 32 и 23%, соответственно) (табл. 2 и 3). Та же закономерность проявилась при исследовании спортсменов одной специализации. Например, у высококвалифицированных легкоатлетов ППСМВ mGS, которую мы рассматриваем как “наиболее нагружаемую”, была существенно выше ППСМВ “менее нагружаемой” mVL (рис. 2). На основании этого факта, а также с учетом относительно небольшой доли собственно силовых упражнений в общей тренировочной нагрузке можно выдвинуть гипотезу, что у спортсменов, тренирующих выносливость, эффект повышения ППСМВ всех типов в “более нагружаемых” мышцах может достигаться, вероятнее всего, за счет механических факторов, действующих в течение нескольких лет регулярных тренировок.
ЗАКЛЮЧЕНИЕ
Результаты настоящего исследования позволяют сделать вывод, что многолетняя спортивная тренировка выносливости приводит не только к повышению окислительного потенциала МВ, как это многократно показано в ряде исследований, но и к увеличению ППСМВ типа I, относительно СС и ССС, а МВ типа IIA относительно ССС. При этом наиболее выраженная гипертрофия наблюдается в мышцах, несущих основную механическую нагрузку по сравнению с такими же активными, но “менее нагружаемыми” мышцами.
Финансирование работы. Работа выполнена в рамках государственного задания ФГБУ ФНЦ ВНИИФК № 777-00026-22-00 (тема № 001-22/5).
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
MacDougall J., Sale D., Elder G., Sutton J. Muscle ultrastructural characteristics of elite powerlifters and bodybuilders // Eur. J. Appl. Physiol. Occup. Physiol. 1982. V. 48. № 1. P. 117.
Tesch P., Larsson L. Muscle hypertrophy in bodybuilders // Eur. J. Appl. Physiol. Occup. Physiol. 1982. V. 49. № 3. P. 301.
Alway S., Grumbt W., Gonyea W., Stray-Gundersen J. Contrasts in muscle and myofibers of elite male and female bodybuilders // J. Appl. Physiol. 1989. V. 67. № 1. P. 24.
Costill D., Daniels J., Evans W. Skeletal muscle enzymes and fiber composition in male and female track athletes // J. Appl. Physiol. 1976. V. 40. № 2. P. 149.
Coyle E., Bell S., Costill D., Fink W. Skeletal muscle fiber characteristics of world class shot-putters // Res. Q. 1978. V. 49. № 3. P. 278.
Green H., Thomson J., Daub W. et al. Fiber composition, fiber size and enzyme activities in vastus lateralis of elite athletes involved in high intensity // Eur. J. Appl. Physiol. Occup. Physiol. 1979. V. 41. № 2. P. 109.
Alway S., Macdougall J., Sale D. et al. Functional and structural adaptations in skeletal-muscle of trained athletes // J. Appl. Physiol. 1988. V. 64. № 3. P. 1114.
Rodríguez L., López-Rego J., Calbet J. et al. Effects of training status on fibers of the musculus vastus lateralis in professional road cyclists // Am. J. Phys. Med. Rehabil. 2002. V. 81. № 9. P. 651.
Ingjer F. Capillary supply and mitochondrial content of different skeletal muscle fiber types in untrained and endurance-trained men. A histochemical and ultrastructural study // Eur. J. Appl. Physiol. Occup. Physiol. 1979. V. 40. № 3. P. 197.
Harber M., Trappe S. Single muscle fiber contractile properties of young competitive distance runners // J. Appl. Physiol. 2008. V. 105. № 2. P. 629.
Trappe S., Costill D., Fink W., Pearson D. Skeletal muscle characteristics among distance runners: a 20-yr follow-up study // J. Appl. Physiol. 1995. V. 78. № 3. P. 823.
Kohn T., Essén-Gustavsson B., Myburgh K. Do skeletal muscle phenotypic characteristics of Xhosa and Caucasian endurance runners differ when matched for training and racing distances? // J. Appl. Physiol. 2007. V. 103. № 3. P. 932.
Kohn T., Essén-Gustavsson B., Myburgh K. Specific muscle adaptations in type II fibers after high-intensity interval training of well-trained runners // Scand. J. Medi. Sci. Sports. 2010. V. 21. № 6. P. 765.
Trappe S., Harber M., Creer A. et al. Single muscle fiber adaptations with marathon training // J. Appl. Physiol. 2006. V. 101. № 3. P. 721.
Hakkinen K., Alen M., Kraemer W. et al. Neuromuscular adaptations during concurrent strength and endurance training versus strength training // Eur. J. Appl. Physiol. 2003. V. 89. № 1. P. 42.
Gollnick P., Armstrong R., Saubert C. et al. Enzyme activity and fiber composition in skeletal muscle of untrained and trained men // J. Appl. Physiol. 1972. V. 33. № 3. P. 312.
Lundberg T., Feuerbacher J., Sünkeler M. et al. The effects of concurrent aerobic and strength training on muscle fiber hypertrophy: a systematic review and meta-analysis // Sports Med. 2022. https://doi.org/10.1007/s40279-022-01688-x
Edstrom L., Ekblom B. Differences in sizes of red and white muscle fibers in vastus lateralis of musculus quadriceps femoris of normal individuals and athletes. Relation to physical performance // Stand. J. Clin. Lab. Invest. 1972. V. 30. № 2. P. 175.
Prince F., Hikida R., Hagerman F. Human muscle fiber types in power lifters, distance runners and untrained subjects // Pflugers Arch. 1976. V. 363. № 1. P. 19.
Costill D., Fink W., Pollock M. Muscle fiber composition and enzyme activities of elite distance runners // Med. Sci. Sports. 1976. V. 8. № 2. P. 96.
Burke E., Cerny F., Costill D., Fink W. Characteristics of skeletal muscle in competitive cyclists // Med. Sci. Sports. 1977. V. 9. № 2. P. 109.
Staron R., Hikida R., Hagerman F. et al. Human skeletal muscle fiber type adaptability to various workloads // J. Histochem. Cytochem. 1984. V. 32. № 2. P. 146.
Tesch P., Karlsson J. Muscle fiber types and size in trained and untrained muscles of elite athletes // J. Appl. Physiol. 1985. V. 59. № 6. P. 1716.
Macková E., Melichna J., Havlíčková L. et al. Skeletal muscle characteristics of sprint cyclists and nonathletes // Int. J. Sports Med. 1986. V. 7. № 5. P. 295.
Tesch P., Thorsson A., Essén-Gustavsson B. Enzyme activities of FT and ST muscle fibers in heavy-resistance trained athletes // J. Appl. Physiol. 1989. V. 67. № 1. P. 83.
Klitgaard H., Zhou M., Richter E. Myosin heavy chain composition of single fibres from m. biceps brachii of male body builders // Acta Physiol. Scand. 1990. V. 140. № 2. P. 175.
Kuzon W., Rosenblatt J., Huebel S. et al. Skeletal muscle fiber type, fiber size and capillary supply in elite soccer players // Int. J. Sports Med. 1990. V. 11. № 2. P. 99.
Alway S.E., Grumbt W.H., Stray-Gundersen J. et al. Effects of resistance training on elbow flexors of highly competitive bodybuilders // J. Appl. Physiol. 1992. V. 72. № 4. P. 1512.
Sleivert G., Backus R., Wenger H. Neuromuscular differences between volleyball players, middle distance runners and untrained controls // Int. J. Sports Med. 1995. V. 16. № 6. P. 390.
Kadi F., Eriksson A., Holmner S. et al. Cellular adaptation of the trapezius muscle in strength-trained // Histochem. Cell Biol. 1999. V. 111. № 3. P. 189.
Terzis G., Stattin B., Holmberg H. Upper body training and the triceps brachii muscle of elite cross country skiers // Scand. J. Med. Sci. Sports. 2006. V. 16. № 2. P. 12.
Ørtenblad N., Nielsen J., Boushel R. The muscle fiber profiles, mitochondrial content, and enzyme activities of the exceptionally well-trained arm and leg muscles of elite cross-country skiers // Front. Physiol. 2018. V. 9. P. 1031.
Boros-Hatfaludy S., Fekete G., Apor P. Metabolic enzyme activity patterns in muscle biopsy samples in different athletes // Eur. J. Appl. Physiol. Occup. Physiol. 1986. V. 55. № 3. P. 334.
Coggan A., Spina R., Rogers M. Histochemical and enzymatic characteristics of skeletal muscle in master athletes // J. Appl. Physiol. 1990. V. 68. № 5. P. 1896.
Sjøgaard G. Muscle morphology and metabolic potential in elite road cyclists during a season // Int. J. Sports Med. 1984. V. 5. № 5. P. 250.
Harber M., Gallagher P., Creer A. et al. Single muscle fiber contractile properties during a competitive season in male runners // Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004. V. 287. № 3. P. 1124.
Mygind E. Fibre characteristics and enzyme levels of arm and leg muscles in elite cross-country skiers // Scand. J. Med. Sci. Sports. 2007. V. 5. № 2. P. 76.
Ahmetov I., Druzhevskaya A., Lyubaeva E. et al. The dependence of preferred competitive racing distance on muscle fibre type composition and ACTN3 genotype in speed skaters // Exp. Physiol. 2011. V. 96. № 12. P. 1302.
Aagaard P., Andersen J., Bennekou M. Effects of resistance training on endurance capacity and muscle fiber composition in young top-level cyclists // Scand. J. Med. Sci. Sports. 2011. V. 21. № 6. P. 298.
Mitchell E., Martin N., Bailey S., Ferguson R. Critical power is positively related to skeletal muscle capillarity and type I muscle fibers in endurance trained individuals // J. Appl. Physiol. 2018. V. 125. № 3. P. 737.
Gejl K., Hvid L., Andersson E. et al. Contractile properties of MHC I and II fibers from highly trained arm and leg muscles of cross-country skiers // Front. Physiol. 2021. V. 12. P. 682943.
Mitsuzono R., Katsuta S., Kanao Y. Relationship between muscle fiber composition, capillary supply, oxidative enzyme activity and aerobic work capacity in middle-and long-distance runners // Japan. J. Physical Fitness Sports Med. 1986. V. 35. № 4. P. 182.
Gerard E., Caiozzo V., Rubin B. et al. Skeletal muscle profiles among elite long, middle and short distance swimmers // Am. J. Sports Med. 1986. V. 14. № 1. P. 77.
Sadoyama T., Masuda T., Miyata H., Katsuta S. Fibre conduction velocity and fibre composition in human vastus lateralis // Eur. J. Appl. Physiol. Occup. Physiol. 1988. V. 57. № 6. P. 767.
Sjöström M., Johansson C., Lorentzon R. Muscle pathomorphology in m. quadriceps of marathon runners. Early signs of strain disease or functional adaptation? // Acta Physiol. Scand. 1988. V. 132. № 4. P. 537.
Fry A.C., Schilling B.K., Staron R.S. et al. Muscle fiber characteristics and performance correlates of male olympic-style weightlifters // J. Strength Cond. Res. 2003. V. 17. № 4. P. 746.
Eriksson A., Kadi F., Malm C. et al. Skeletal muscle morphology in power-lifters with and without anabolic steroids // Histochem. Cell Biol. 2005. V. 124. № 2. P. 167.
Kesidis N., Metaxas T.I., Vrabas I.S. et al. Myosin heavy chain isoform distribution in single fibres of bodybuilders // Eur. J. Appl. Physiol. 2008. V. 103. № 5. P. 579.
Staron R.S., Hagerman F.C., Hikida R.S. The effects of detraining on an elite power lifter // Neurol. Sci. 1981. V. 51. № 2. P. 247.
Fitts R.H., Costill D.L., Gardetto P.R. Effect of swim exercise training on human muscle fiber function // J. Appl. Physiol. 1989. V. 66. № 1. P. 465.
Billeter R., Jostarndt-Fögen K., Günthör W. et al. Fiber type characteristics and myosin light chain expression in a world champion shot putter // Int. J. Sports Med. 2003. V. 24. № 3. P. 203.
Korhonen M.T., Cristea A., Alén M. et al. Aging, muscle fiber type, and contractile function in sprint-trained athletes // J. Appl. Physiol. 2006. V. 101. № 3. P. 906.
Andersen J.L., Klitgaard H., Bangsbo J. et al. Myosin heavy chain isoforms in single fibres from m. vastus lateralis of soccer players: effects of strength-training // Acta Physiol. Scand. 1994. V. 150. № 1. P. 21.
Terzis G., Spengos K., Kavouras S. et al. Muscle fiber type composition and body composition in hammer throwers // J. Sports Sci. Med. 2010. V. 9. № 1. P. 104.
Trappe S., Luden N., Minchev K. et al. Skeletal muscle signature of a champion sprint runner // J. Appl. Physiol. 2015. V. 118. № 12. P. 1460.
Malisoux L., Francaux M., Theisen D. What do single-fiber studies tell us about exercise training? // Med. Sci. Sports Exerc. 2007. V. 39. № 7. 1051.
Van Wessel T., de Haan A., van der Laarse W.J. et al. The muscle fiber type-fiber size paradox: hypertrophy or oxidative metabolism? // Eur. J. Appl. Physiol. 2010. V. 110. № 4. P. 665.
Krogh A. The number and distribution of capillaries in muscle with calculations of the oxygen pressure head necessary for supplying the tissue // J. Physiol. 1918. V. 52. P. 409.
Wagner P.D. CrossTalk proposal: Diffusion limitation of O2 from microvessels into muscle does contribute to the limitation of VO2max // J. Physiol. 2015. V. 593. № 17. P. 3757.
Hirai D.M., Colburn T.D., Craig J.C. Skeletal muscle interstitial O2 pressures: bridging the gap between the capillary and myocyte // Microcirculation. 2019. V. 26. № 5. P. e12497.
Henneman E., Somjen G., Carpenter D. Functional significance of cell size in spinal motoneurons // J. Neurophysiol. 1965. V. 28. P. 560.
Myakinchenko E.B., Kriuchkov A.S., Adodin N.V. et al. The annual periodization of training volumes of international-level cross-country skiers and biathletes // Int. J. Sports Physiol. Perform. 2020. V. 15. P. 1181.
Torvik P., Solli G., Sandbakk Ø. The training characteristics of world-class male long-distance cross-country skiers // Front. Sports Act. Living. 2021. V. 3. P. e641389.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека