

Физиология человека, 2022, T. 48, № 6, стр. 3-31
Нейрофизиологические характеристики придумывания альтернативного использования по данным анализа вызванных потенциалов (ВП) и связанной с событиями синхронизации/десинхронизации ЭЭГ в зависимости от уровня продуктивности и оригинальности испытуемых
Ж. В. Нагорнова 1, *, В. А. Галкин 1, В. А. Васенькина 1, А. В. Грохотова 1, Н. В. Шемякина 1, **
1 ФГБУН Институт эволюционной физиологии и биохимии имени И.М. Сеченова РАН
Санкт-Петербург, Россия
* E-mail: nagornova_zh@mail.ru
** E-mail: shemyakina_n@mail.ru
Поступила в редакцию 22.07.2022
После доработки 02.08.2022
Принята к публикации 05.08.2022
- EDN: CFGYDH
- DOI: 10.31857/S0131164622700163
Аннотация
В ЭЭГ/ВП-исследовании участвовали 44 испытуемых (26 м : 18 ж, средний возраст: 20 ± 1.8 лет), выполнявших модифицированный тест альтернативного использования (“AUT” – придумать оригинальные способы использования объектов) и контрольную к нему задачу – перечислить объекты из заданных категорий. Испытуемым предъявляли 7–8 названий предметов (альтернативное использование) и семь-восемь категорий объектов – на работу с каждым стимулом отводили 10 проб-попыток длительностью 5400 мс. Десятипробные блоки предъявлений творческого и контрольного заданий были рандомизированы между собой (суммарно предъявляли 70–80 творческих и 70–80 контрольных проб). В группе более продуктивных участников (по количеству ответов в задании альтернативного использования) негативность амплитуд при творческом задании N300–N400 была меньше в центральных и теменных областях, а амплитуда P600 была больше в лобных областях при выполнении творческого задания по сравнению с контрольным. В группе менее продуктивных участников в AUT наблюдалась меньшая амплитуда P200 и более негативные амплитуды для N300–N400 компонента в лобных областях при выполнении творческого задания по сравнению с контрольным. Авторы данной статьи предположили, что бóльшая продуктивность связана с активацией семантической сети, в то время как у участников с меньшей продуктивностью задействованы механизмы зрительного внимания, что, вероятно, оказывается менее эффективным для продуктивности в деятельности. В группе с большей продуктивностью в AUT, по сравнению с контрольным заданием, наблюдается десинхронизация ЭЭГ 7–9 Гц в лобных областях на 356–564 мс после предъявления стимула. В группе с низкой продуктивностью различий между творческими и контрольными пробами не наблюдалось. При выполнении творческого задания группой с более высоким уровнем невербальной оригинальности (по тесту Торренса “Эскизы”) выявлена бóльшая десинхронизация ЭЭГ 5–8 Гц после предъявления стимула (до 296 мс) в лобных и теменных областях по сравнению с группой с меньшим уровнем оригинальности. Таким образом, высказано предположение, что продуктивность и оригинальность при выполнении AUT оказывают различный эффект на параметры ВП и вызванной синхронизации/десинхронизации ЭЭГ.
Изучение нейрофизиологических характеристик творческой деятельности реализуется в различных модельных условиях [1, 2]. Обычно под творческой деятельностью понимают способность создавать что-то новое и полезное [3], отклоняться в мышлении от стереотипов [4–6] и находить решения при минимуме доступной информации [7, 8].
В настоящей работе изучались характеристики электроэнцефалограммы (ЭЭГ) и вызванных потенциалов (ВП) выполнения модифицированного задания альтернативного использования [9–10] при субъективной оценке испытуемыми результатов его выполнения.
Тест альтернативного использования (Alternative uses task, AUT) является одним из широко используемых заданий для оценки творческих способностей в психологии [11, 12] и одной из модельных задач в нейрофизиологических исследованиях творчества и дивергентного мышления [13–35] (табл. 1). Считается, что AUT позволяет оценить беглость, оригинальность мышления и является надежным психометрическим инструментом определения творческого потенциала человека [36, 37].
Таблица 1.
Обзор основных работ по изучению нейрофизиологических характеристик выполнения теста альтернативного использования за последние тринадцать лет
| Источник | Испытуемые | Задание AUT | Контрольное задание (либо задача сравнения) |
Методика | Результаты |
|---|---|---|---|---|---|
| (а) N (муж : жен) (b) Средний возраст, лет (c) Группы (d) Тесты разделения |
(а) Инструкция использования (b) N стимулов, примеры (c) Длительность пробы (d) Доп. инструкция |
(а) Задача испытуемому (b) N стимулов, примеры (c) Длительность пробы |
(а) Метод (b) Параметры анализа сигнала (c) Области интереса (d) Контрасты/ сравнения |
||
| [13] | (a) 49 (26 м : 23 ж) (b) 23.1 ± 2.64 (c) – (d) AUT Задания за пределами МРТ-сканера |
(a) Оригинальное (b) 3: карандаш, туфли, очки (c) 2 мин (d) Экспертная оценка оригинальности (1; 5) |
(a1) Дискриминировать
картинку: животное/еду
в зависимости
от условий
предъявления
(запоминание) (a2) Вспомнить: был/нет объект (извлечение из памяти) (b1) 80 (запоминание); клубника (b2) 210 (извлечение); павлин (c) >3 с |
(a) фМРТ
в состоянии покоя (b) Функциональная связность (ФС) (c) Передний/задний гиппокамп и лобная кора (d1) GLMM для оценки взаимосвязи: оригинальности, кол-ва ответов AUT, ВР и точности при запоминании; узнанных объектов; (d2) LME для ФС гиппокампа; оригинальность AUT – непрерывный предиктор |
1. Нет взаимосвязи между:
креативностью и когнитивным контролем;
креативностью и эпизодической памятью 2. Креативность > при меньшей разнице запомненных предметов/правила дискриминации (когнитивный контроль × память) 3. Креативность коррелирует: – положительно с ФС между Hyp и задней частью SFG ЛП, anterior Hyp и anterior MFG ЛП; – отрицательно с ФС между anteriorHyp и anterior SFG ЛП |
| [14] | (а) 24 (12 ж : 12 м) (b) 32.7 ± 9.66 (c) – (d) Тест Струпа |
(a1) Смешное (b) 15 (рэндом с контролем): возд. шарик (c) 15 с |
(a) Обычное (b) 15 (рэндом с творческим): мыло (c) 15 с |
(a) фМРТ (b1) BOLD (b2) gPPI анализ ФС верхней/медиальной лобных извилин ПП (c) Верхняя/средняя лобные извилины ПП, миндалина (d) AUTvs Контроль (тест Струпа – ковариата) |
1. Высокий когнитивный контроль
связан с большей активацией миндалины,
гиппокампа, SFG, MFG ПП во время генерации смешного использования 2. Активация миндалины ЛП при генерации смешного использования положительно коррелирует с количеством идей и обратно связана с активацией лобных зон (SFG, MFG ПП) |
| [15] | (а) 116 (45 м : 69 ж) (b) 21.48 ± 2.05 (c1) Гр. HD (с2) Гр. LD |
(а) Оригинальное (b) – , книга (c) 300 с |
(а) Перечислить
характеристики
предмета (OCT) (c) 300 с |
(а) fNIRS [окси/
дезокси-Hb] (b) IBS – индекс межсубъектной синхронизации (c) 12 ROI (лоб, висок, темя ПП) (d) AUTvsОСТ |
Между группами: 1. Гр. LD беглость AUT > HD 2. Гр. HD гибкость мышления > LD 3. Гр. LD: увеличение IBS в правой угловой извилине и правой соматосенсорной коре > HD |
| [16] | (a) 28 ж (b) 22.71 ± 2.80 (c1) Гр. 15 чел. (>AUT) (с2) Гр. 13 чел. (<AUT) (d1) AUT – деление гр. (d2) IQ; ATTA |
(a) Оригинальное (b) 5: кирпич, бутылка, шина, газета, зонт (c) 2 мин (d) Задание AUT без регистрации ЭЭГ |
(a) Ответить: да/нет: обычная фраза/есть смысл? (b) 132: 3 типа фраз – новые метафоры/ бессмысленные/ буквальные (c) 9.6–10 с |
(a) ЭЭГ (ВП) (b1) Амплитуда N400 (350–500 мс) (b2) Амплитуда поздних компонентов (500–900 мс) (c) Центральные, теменные (d1) Метафора vs бессмысленная фраза vs буквальная фраза; (d2) Группы высокий vs низкий AUT |
В общей группе:
1. Амплитуды N400 бессмысленная фраза = новая метафора > буквальная фраза 2. Амплитуда P500–900 растет в ряду: бессмысленная фраза > новая метафора > > буквальная фраза Гр. высокий AUT: 1. Амплитудa P500–900 > для всех типов фраз vs гр. низкий AUT; 2. Pазница амплитуд: метафора бессмысленные фразы > чем гр. низкий AUT |
| [17] | (a) 29 (8 м : 21 ж) (b) 19.55 ± 1.99 (c) AUT (9 высоко : 9 низкокреативных) |
(a) Оригинальное (b) 30 (c) До 40 с |
(a) Обычное
использование, СTest (b) 30 проб (c) До 40 с |
(a) ЭЭГ, машинное
обучение (b) Spec-CSP (c) θ, α1, α2, β, γ, непрерывные спектры (d1) AUTvs СTest; (d2) Классификация AUT, CTest |
1. При AUT мощность >, чем
в контроле в 8.6–10 и 10–11.5 Гц в F4;
8.6–10 и 10.3–11.8 Гц в Р4 2. Точность 2-классовой классификации AUTvsСTest в среднем 63.9%; > Точность классификации ЭЭГ высоко и низко-креативных участников 82.3% |
| [18] | (а) 36 (17 м : 19 ж) (b) 50.9 ± 13.7 (c1) “Творцы”: 20 (11 м : 9 ж) 51.8 ± 12.48; (c2) Контр. гр.: 16 (6 м : 10 ж), 50 ± 15.04. (d) Опросник достижений |
(a) Новое/необычное (b) 72 предмета (ч/б изображения): кувшин, стул (c) 9 с |
(a) Определить – есть/нет черный
квадрат
поверх изображения (b) 72 предмета (ч/б размытые изображения): кувшин, стул (c) 3 с |
(a) фМРТ (b) BOLD (c) DMN (PreCu, SPL, AG, MPFC, aMTG), ECN (SFG, MFG, IFG) (d) (Творческое задание vs контрольное задание) × × (“Творцы” vs контр. гр.) |
1. По беглости различий
между группами нет 2. В гр. “Творцы” кластеров активации при AUT (ПП: SMG, TTG) <, чем в контр. гр. (ЛП: LG, FP, MFG, ACC, preCu, MFG, HypG; ПП: AG, PCC, FusG) |
| [19] | (a) 93 (41 м : 42 ж) (b) 23.05 ± 3.61 |
(a) Оригинальное (b) 16 рисунков: вилка (c) 25 с (d) Оценка оригинальности ответов (1; 6) шкала |
(a) Понять смысл
2 типов юмористических зарисовок (b) 15 рисунков/тип (c) 30 с (d1) Смешно/ несмешно (1; 6); (d2) Объяснить шутку |
(a) ЭЭГ (b) TRP в α1 (10–12 Гц) (c) Fp1/2, F3/4, F7/8P3/4, P7/P8, O1/2, С3/4 (d1) Сравнение заданий; (d2) Сравнение высокая/низкая оценка AUT |
В каждом из заданий: увеличение мощности α1 в Fp1/2, F3/4, F7/8; уменьшение в P3/4, P7/P8, O1/2 При высокой/низкой оценке AUT у тех, кто оценил высоко: Cинхронизация в Fp1/2 и F3/4<; в С4 >; С3 десинхронизация >; P3/4 десинхронизация >; P7 десинхронизация <; P8 >; O1 десинхронизация <. Синхронизация в AUT в ПП < при понимании юмора |
| [20] | (а) 132 (74 ж : 58 м) (b) 21.23 ± 2.91 (с1) 26 пар Ж–Ж; (c2) 22 пар Ж–M; (c3) 18 пар M–M |
(а) Оригинальное | (a) Описание характеристик (ОСТ) | (а) fNIRS (b) Индивидуальная [окси/дезокси–Hb] (с) Префронтальные и правые височно- теменные области (d) AUTvsОСТ |
Гр. Ж–Ж
при выполнении AUT: 1. > высокие уровни кооперативного поведения и гибкости 2. Повышение IBS в правой задней теменной коре при AUT |
| [21] | (a) 86 (30 м : 56 ж) (b) 23.36 ± 3.55 |
(a) Оригинальное (b) 16: зонт, кирпич, ключ (c) 1 мин (d) Экспертная оценка оригинальности (1; 4) |
(a) Время нахождения ответа делили
на три интервала (b) Сравнивали эти интервалы до нахождения ответа |
(a) ЭЭГ (b) TRP/TRPL в α (10–12 Гц) (c) Лобные и теменно-затылочные (d) Сравнение интервалов поиска ответа |
1. У более креативных участников:
U-образная кривая изменений мощности α
по интервалам и увеличение
функциональной связи между лобными
и теменно-затылочными областями ко второму интервалу 2. У менее креативных – нет различий мощности α между интервалами и увеличение функциональной связи к третьему интервалу |
| [22] | (а1) 20 (10 м : 10 ж) (b) – (c1) 10 – контр. гр. (6 м : 4 ж); (c2) 10 – плацебо (4 м : 6 ж) |
(a) Оригинальное (b) 3: кирпич, скрепка, ботинок (c) >3 мин (d) Оценка оригинальности по частоте ответов. Неоригинальные ответы не засчитывались |
(a1) Умножение
двузначных чисел (b1) Индивидуально (с1) Максимум 10 мин (a2) Назвать как можно больше предметов с определенной функцией (Instances Task, IT) (b2) 3: шестерня, шум, края (c) 3 мин |
(a1) ЭЭГ/sham tDCS
(во время IT) (b1) SVM-классификации состояний и групп; (b2) Мощность ЭЭГ: θ (5–7.5 Гц), α (7.5–12.5 Гц), β (12.5– 30 Гц) и γ (30–60 Гц); кросс-спектр (диапазон/канал) – всего 189 признаков (c2) Лобная зона (d1) Математическое задание vs AUT; (d2) Гр. плацебо vs контр. гр. при AUT |
1. Не было различий
в выполнении творческих задач
между контр. и плацебо группами 2. Классификация математического задания и AUT по данным ЭЭГ – с точностью до 87% 3. Наблюдается плацебо эффект в изменениях ЭЭГ (классификация плацебо и контр. гр. на уровне 92%) |
| [23] | (а) 20 ж (b) 22.0 ± 1.8 (с) Самооценка ориг. ответов (1; 5 – max) |
(a) Оригинальное (b) 30 изображений/ 4 попытки (всего 120 проб); колесо, пенал, часы, стол, кнопка (c) 24 c |
(a) Сравнение 4-х попыток нахождения решения | (a) ЭЭГ (b) TRP α (8–12 Гц) (c) Все отведения (d1) Сравнивали TRP при разных попытках ответа; (d2) Корреляция αTRP с оригинальностью ответа |
1. Рост оригинальности, снижение беглости
с номером попытки 2. α-Десинхронизация на первый ответ и последующая синхронизация (2–4 ответы) во всех зонах интереса 3. Синхронизация в α-диапазоне связана с оригинальностью идей: – в лобных, центральных и височных областях ЛП – в первой фазе AUT; – в центрально-теменных областях ЛП – во всех фазах AUT |
| [24] | (a) 90 (45 м : 45 ж) (b) 23.8 ± 2.3 (с) Гр. tDCS стимуляции (c1) L + R– 30 (15 м : 15 ж); (c2) R–L+ 30 (15 м : 15 ж); (c3) Плацебо 30 (15 м : 15 ж) (d) Compound RAT (СRAT) |
(а) Оригинальное | – | (a) tDCS (b) 1 мА в течение 22 мин (c) Стимуляция iFG, два электрода на пересечении T4–Fz, F8–Cz ПП и T3–Fz, F7–Cz ЛП (d) Межгрупповое сравнение |
1. При активации правой и деактивации левой префронтальной коры беглость AUT
и CRAT повышалась 2. При деактивации левой и активации правой префронтальной коры беглость AUT и CRAT понижалась |
| [25] | (а) 26 мужчин (b) 18.42 ± 0.74 (c1) До стресса; (с2) После стресса (d) MIST |
(a) Оригинальное (b) 30: 15 пред.: 15 для пост. сессий (c) 28 с |
(a) ЭЭГ (b) TRP в α2 (10–12 Гц) (c) – (d1) До и после стресса (MIST); (d2) Ориг. vs неориг. |
1. При выполнении задания до стресса, наблюдалась синхронизация >
> после стресса 2. Ответы до стресса были оценены как более оригинальные 3. TRP выше в оригинальных идеях и более выражен в правом полушарии |
|
| [26] | (а) 46 (37 ж : 9 м) (b) 19.2 ± 0.941 (с) – |
(а) Оригинальное (b) 3 (12 повторений): кирпич, швабра скрепка (c) 25 с |
(а) Перечислить
предметы, категории (b) 3 (12 повторений): страны мира (c) 25 с |
(а) ЭЭГ (ERD) (b) TRP α1 (8–10.5 Гц), α2 (10.5–13 Гц) (c) Лобная, теменно-затылочная зоны (d1) Творческое vs контрольное задания; (d2) Правое vs левое полушария; (d3) Лобная vs теменно-затылочная зоны |
1. Снижение беглости
и увеличение оригинальности идей
при увеличении номера ответа 2. Большее значение мощности α2 в AUTvs КАТЕГОРИИ 3. Увеличение мощности α2 в ЛП по отношению к ПП при большем количестве ответов в AUT 4. Увеличение α1-ритма в ПП при выполнении творческого и контрольного заданий 5. Мощность α1-ритма в лобной зоне ЛП > > чем в теменно-затылочной при творческом задании |
| [5] | (а) 21 (11 ж : 10 м) (b) 20.3 ± 2.18 (с) Remoteness “отдаленность” идей (d) Тест Хейлинга (завершить предложения) |
(а) Оригинальное, предъявление:
“объект – классическое применение” (b) 7 предметов (изображение/ печатное слово/озвучивание); шапка – защищать голову (с) 2 мин 5 с |
(a) Оригинальное,
предъявление только объекта (b) 7 предметов (изображение/печатное слово/озвучивание): шапка, велосипед, зонт (c) 2 мин 5 с |
(a) ЭЭГ (b) TRP (task-related power), мощность α (c) Лобная и височно-теменная зоны (d1) Участники с высоким показателем remoteness vs с низким; (d2) Преодоление стереотипа vs контрольное задание |
1. α-Синхронизация в лобных долях у участников с большими показателями параметра “отдаленности”
при выполнении контрольного задания > чем при преодолении стереотипа 2. α-Десинхронизация в височно-теменной зоне у участников с низким показателем “remoteness” > чем у участников с высоким показателем при преодолении стереотипа 3. ВР при выполнении теста Хейлинга у участников с высоким показателем “отдаленности” < чем у участников с более низкими показателями |
| [27] | (a) 35 (10 м : 25 ж) (b) 22.6 ± 2.02 (c) Гр. по уровню управляющих функций – актуализации (запоминание букв), торможения (Тест Струпа), переключения (Тест “цифра-буква”) |
(a) Оригинальные
(вторые 1.5 мин
задания) (b) 8: карандаш, ключ, часы, ботинки, стол, пуговица, губная помада, мобильный телефон (с) 3 мин |
(а) Оригинальное (первые 1.5 мин задания) | (a) ЭЭГ (b) ERS/ERD в θ(4–8 Гц), α1(8–10 Гц), α2(10–13 Гц), β1(13–20 Гц), β2(20–30 Гц) (c) FP3/4; F3/4; F7/8; FC1/2; FC5/6; C3/4; P3/4; P7/8; T7/8; O1/2 (d) Вторые 1.5 мин (1.6–3 мин) vs первые 1.5 мин (0–1.5 мин) работы со стимулом |
1. В общей группе: снижениебеглости и увеличение оригинальности дивергентного мышления
со временем 2. В группе с высокой переключаемостью – рост оригинальности во вторые 1.5 мин vs первые 1.5 мин 3. В группе с низким торможением – снижение мощности α2 (10–13 Гц) в PF1, F3, F7 в сравнении с первыми 1.5 мин 4. Во вторые 1.5 мин мощность β1 в ПП > ЛП, в первые 1.5 мин такой разницы нет |
| [28] | (а) 16 (4 м : 12 ж) (b) 31.3 |
(а) Оригинальное (b) 11 (изображения): покрышка, губная помада, скрепка Self-pased |
(a) Идентификация
объекта (b) См. лев. столбец (c) 3.6 с |
(а) фМРТ (b) BOLD (c) whole brain анализ (d) “Генерация” vs “Идентификация” с учетом порядкового номера идей |
1. “Генерация” vs “Идентификация” – активация переднего части островка ЛП 2. Положит. корреляция порядка идеи с активацией PreCu/Cu билатерально PСC, ACC; IPL (SMG, AG) – ПП 3. Отриц. корреляция порядка идеи с активацией в ppreSMA, PMC, IFG, IFJ – ЛП |
| [29] | (a) 27 (13 м : 14 ж) (b) 23.11 ± 2.06 (c) – |
(a) Оригинальное (b) 15: когтеточка, катапульта, браслет (c) 60 с |
(a) Характеристики
объекта (OCT) (b) 15: когтеточка, катапульта, браслет (c) 60 с |
(a) ЭЭГ (b) Фазово-амплитудное сопряжение, отн. P (с) θ (4–8 Гц), α1 (8–10 Гц), α2 (10–13 Гц), β1 (13–20 Гц), β2 (20–30 Гц), γ (30–40 Гц) (d) AUTvs OCT в θ-γ и α2-γ |
1. В AUT > OCT: фазово-амплитудное сопряжение ЭЭГ в θ-γ (CP4 и CP6);
α2-γ (FC1 и Cz) 2. Нет различий относительной мощности между заданиями |
| [30] | (а) 35 (11 м : 24 ж) (b) 22.7 |
(а) Oригинальное (b) 15: покрышка (c) 60 с (d1) Оценка идеи “старая” (вспомнил)/ “новая” (придумал); (d2) Эксп. оценка (креативность: 1; 4) |
(а) фМРТ (b1) BOLD (c) whole brain анализ (d1) Ответ vsbaseline; (d2) “Новые” vs “старые”; (d3) Более творческие идеи vs менее творческие |
1. При наличии ответа: активация ЛП: IFG (ПБ 47,46), SFG, PreCG, PostCG.
Активация ПП: гиппокамп, ITG (ПБ 20) Деактивация: ПП: SMG, AG, MTG, STG, PreCu, PCC 2. “Новые” vs “старые”: активация: ЛП: IPС (ПБ 40) SMG 3. Более творческие идеи vs менее творческие: активация ЛП: IFG (orb) (ПБ 47) |
|
| [31] | (a) 20 (11 ж : 9 м) (b) 22.55 ± 2.1 (c) 8.4−8.7 c |
(a) Последовательная оценка необычности/ конгруэнтности предмета и его использования (b) 45 пар: “ботинок – посадить растение” (c) 8.4-8.7 c |
(a) ЭЭГ (ВП) (b) Амплитуды ВП (c) C1, Cz, C2, CP1, CPz, CP2, P1, Pz, P2 (d) Творческие комбинации (необычно и приемлемо) vs бессмысленные (необычно и неприемлемо) vs обычные (обычно и приемлемо) |
1. Амплитуда N400
(300–500 мс) > (более негативна)
для бессмысленных и творческих
комбинаций vs обычные 2. Амплитуда и латентность N400 для творческих vs бессмысленные пары – не отличались 3. Амплитуда компонента 500–900 мс для комбинаций “необычно и приемлемо” (творческие) >, чем для “необычно и неприемлемо” |
|
| [32] | (a) 55 (b) 16.79 ± 0.68 (c) EPQ-R |
(a) Оригинальное (b) 10: кирпич (c) 40 с (d) Эксп. оценка(1; 4) |
(a) Обычное
использование (b) 10 предметов; кирпич (c) 40 с |
(a) ЭЭГ (b) TRP α (7.5–12.5 Гц) (c) От лобных до затылочных (d) Оригинальное vs обычное |
1. Десинхронизация α-ритма в контрольном задании > чем в творческом (все зоны) 2. Снижение мощности α-ритма от передне-лобных до центрально-теменных областей при контрольном задании |
| [33] | (a) 32 (25 ж : 9 м)
танцора (b1) Проф.: 26.93 ± 6.41; (b2) Нач.: 23.94 ± 3.67 (c1) 15 проф. (11 ж); (c2) 17 нач. (14 ж) |
(a) Оригинальное (b) 4: кирпич, банка, носок, ручка (c) 3 мин |
(a) Вообразить оригинальные танцевальные движения/известные движения (вальс) (b) 1 (c) 3 мин |
(a) ЭЭГ (b) TRP α1 (8–10 Гц), α2 (10–12 Гц) (c) От лобных до затылочных (d1) AUTvs контрольное задание; (d2) Профессиональные танцоры vs начинающие; (d3) Правое полушарие vs левое |
1. α1-Синхронизация в лобной зоне
при AUT > чем при воображении
оригинальных движений > чем при воображении вальса 2. Снижение синхронизации α1-ритма от лобных к теменно-затылочным областям ЛП мозга при AUT 3. α1,2-Синхронизация в затылочной зоне ПП > ЛП (при AUT) 4. α2-Синхронизация в центрально-теменной, теменно-височной и теменно-затылочной областях профессиональных танцоров > чем у начинающих |
| [34] | (a) 47 (22 ж : 25 м) (b) 24.09 ± 2.95 |
(a) Оригинальное (b) 8: банка, зонт (c) 28 c (d) Эксп. оценка (5; 1) |
(a1) Назвать типичные характеристики
объекта; (a2) Придумать правдоподобную расшифровку аббревиатуры; (a3) Завершить слово (b) 8: туфли, вешалка (c) 28 c |
(a) ЭЭГ (b) TRP α1 (8–10 Гц), α2 (10–12 Гц) (c) От лобных до затылочных (d1) Творческое vs контрольное задание; (d2) Правое vs левое полушарие; (d3) Высокая оригинальность vs низкая |
1. AUT – α-синхронизацией в лобных и рост синхронизации в теменных областях 2. α2-Синхронизация в центрально-теменной и теменно-затылочной областях ПП у людей с высокой оригинальностью > чем у людей с низкой 3. α1-Синхронизация в ПП при AUT > чем при контрольных |
| Нагор-нова Ж.В. и др., 2022 | (a) 44 (26 м : 18 ж) (b)18–23 (c1) Продуктивные/непродуктивные (ответы AUT); (c2) Более/менее оригинальные (“Эскизы” Торренса); (c3) Более/менее оригинальные (AUT – “Коробки” Торренса) |
(a) Оригинальное,
как никто другой
на вашем месте (b) 7–8 предметов × × (10 попыток): газета, гвоздь, книга, скрепка и т.д. (c) 5400 мс |
(a) Перечислить предметы из категории (К) (b) 8 × 10 (попыток): транспорт, одежда и т.д. (с) 5400 мс |
(a) ЭЭГ (b) ERS/ERD, ВП (с) 15 отведений ЭЭГ (d) AUTvs КАТЕГОРИИ |
1. Группа более продуктивных участников (AUTvs К): 1.1. амплитуда N300–N400 < в центральных и теменных зонах; 1.2. амплитуда P600 > в лобных областях; 1.3. десинхронизация ЭЭГ 7–9 Гц в лобных областях на 356–564 мс после стимула 2. Группа менее продуктивных участников (AUT vs К): 2.1. амплитуда P200 < во всех отведениях; 2.2. амплитуда N300–N400 > в лобных областях 3. Сравнение групп по уровню оригинальности в творческом задании: гр. более оригинальных > десинхронизации ЭЭГ 5–8 Гц после предъявления стимула (до 292 мс) в лобных и теменных зонах по сравнению с гр. менее оригинальных |
Примечание: AG – Аngular Gyrus (ангулярная/угловая извилина); aMTG – anterior Middle Temporal Gyrus (передняя средняя височная извилиня); ATTA – Abbreviated Torrance Test for Adults (сокращенный вариант теста креативности П. Торренса); AUT – Alternative Uses Task (тест альтернативного использования); DMN – Default Mode Network (сеть пассивного режима работы мозга); ECN – Executive Control Network (сеть исполнительного контроля мозга); ERS/ERD – event-related synchronization/desynchronization (связанная с событием (вызванная) синхронизация/десинхронизация); fNIRS – functional Near-Infrared Spectroscopy (функциональная спектроскопия в ближней инфракрасной области); HD – high educational diversity (испытуемые в паре специализировались в разных естественных/социальных науках); IBS – Interpersonal Brain Synchronisation (показатель корреляции или когерентности сигналов мозга между взаимодействующими участниками); IFG – inferior frontal gyrus (нижняя лобная извилина), GLMM – general linear mixed model (обобщенная линейная смешанная модель); LME – linear mixed-effects (модель смешанных эффектов); LD – low diversity education (оба испытуемых в паре специализировались в естественных\социальных науках); MFG – Middle Frontal Gyrus (средняя лобная извилина); MIST – Montreal Imaging Stress Task (монреальский тест визуализации стресса); MPFC – Medial Prefrontal Cortex (медиальная префронтальная кора); OCT – Object Characteristics Task (описание характеристик предмета); PAC – Phase-to-Amplitude Coupling (фазо-амплитудная связь, при которой амплитуда высокочастотного сигнала модулируется фазой низкочастотных колебаний); PreCu – Precuneus (предклинье); RAТ – The Remote Associates Test (тест отдаленных ассоциаций Медника); RBF – radial; basis function (радиальная базисная функция); ROI – Region of Interest (зоны интереса); self-paced – переход к следующему стимулу осуществляется по произвольному нажатию на кнопку; SFG – Superior Frontal Gyrus (верхняя лобная извилина); SPL – Superior Parietal Lobule (верхняя теменная долька); SVM – Support Vector Machine (метод опорных векторов); tDCS – transcranial Direct Current Stimulation (транскраниальная микрополяризация); TRP (Task-Related Power [35]) (связанная с заданием мощность, сопоставляется с предстимульным интервалом); TRPL – Task-Related Phase-Locking (изменение связанной с заданием функциональной связи); Δ, θ, α1, α2, β1, β2, γ – дельта, тета, альфа-1, альфа-2, бета-1, бета-2, гамма-диапазоны ЭЭГ соответственно; Альфа индекс – процент времени в которое наблюдается α-ритм; ВП – вызванные потенциалы; ВР – время реакции; ЛП – левое полушарие; ПП – правое полушарие; фМРТ – функциональная магнитно-резонансная томография; ЭЭГ – электроэнцефалография.
В приводимом исследовании испытуемым давалось десять попыток придумать использование одного и того же предмета. Ряд исследователей рассматривают такой вариант инструкций, как связанный с ростом оригинальности ответов во времени [23, 27], а также c ростом и сложности задачи.
Можно предположить, что в ситуации субъективного оценивания собственных ответов как “инсайтные/не инсайтные/трудно определиться” вероятность повышения оригинальности ответов во времени присутствует, но инсайтные ответы могут возникать и при первых попытках выполнения задания в условиях инструкции быть оригинальным.
Исследование осуществлялось с использованием метода вызванных потенциалов и анализа изменений связанной с событиями синхронизации/десинхронизации ЭЭГ при выполнении АUT в сравнении с контрольной задачей – внутри групп испытуемых с разным уровнем продуктивности (по количеству ответов в задании AUT) и оригинальности (согласно субтесту Торренса “Эскизы”), а также при межгрупповом сравнении выполнения заданий с учетом уровня оригинальности испытуемых.
Ожидалось, что при анализе связанной с событиями синхронизации/десинхронизации ЭЭГ мы выявим эффекты изменений в α- и, возможно, в θ-диапазонах ЭЭГ, как весьма чувствительных к выполнению творческих задач, в том числе связанных с заданием альтернативного использования [38–40].
Так, в исследованиях C. Martindale и D. Hines было показано, что высокие баллы в заданиях альтернативного использования соответствовали высокому индексу α-ритма (зоны интереса – правая теменная и затылочная области мозга) испытуемых при выполнении заданий AUT, RAT (The Remote Associates Test – тест отдаленных ассоциаций Медника) и интеллектуального теста по сравнению с испытуемыми с низкими баллами по AUT. В целом это подтверждалось и более поздними работами [39]. При характеристике творческих состояний через уровень активации коры получалось [38, 41], что при реализации творческой деятельности он ниже (т.е., α-ритма, в основном больше), чем в состояниях интеллектуальной деятельности. При этом в исследовании [40] отмечались бóльшие значения δ-, θ- и β-частот при выполнении задачи дивергентного мышления (AUT) в сравнении с задачами конвергентного мышления (вычисления в уме).
Дискуссия о роли θ- и α-частот в творческом мышлении сохраняется (табл. 1) [42–46]. Увеличение θ-частот обсуждается в целом в творческой деятельности [43], творческом письме [47], при эскизировании (больший процент частот 5–6 Гц в лобных, 6–7 Гц в теменных зонах коры vs контрольное рисование [46]) и интуитивном мышлении (особенно во фронтальных областях), связанном с загрузкой памяти и когнитивным контролем [48]. Увеличение θ-активности также связывают с механизмами ассоциативной памяти и когнитивного контроля [49, 50].
Заданий с AUT в парадигме ВП и связанной с событиями синхронизации/десинхронизации ЭЭГ не так много (табл. 1). Косвенным образом в ранних работах [51] было показано, что более креативные испытуемые характеризуются большей амплитудой и вариабельностью компонентов N100 и P300 в oddball тестах, что авторами объяснялось большей гибкостью процессов внимания, возможно, отличающей более креативных испытуемых.
Можно предположить, что выполнение AUT в сравнении с контрольной задачей может быть, в нашем случае, связано с различиями в поздних когнитивных компонентах ВП. Зонами же отличий станут теменно-затылочные и лобные области коры соответственно. Исследования по картированию мозговой активности описывают вовлечение лобной, теменной коры и зон мозга, соотносимых с сетью пассивной работы мозга и др., в творческое мышление в целом [14, 28, 52–57]. Было показано, что при транскранильной стимуляции правой нижней лобной извилины и деактивации левой префронтальной коры креативность повышалась (AUT, RAT), при обратном действии – понижалась [24].
МЕТОДИКА
В исследовании принимали участие 44 здоровых испытуемых обоих полов с нормальным или корректированным зрением: 26 мужчин, 18 женщин (18–23 года), студенты 1–3 курса вузов г. Санкт-Петербурга.
Психологическое исследование. Перед ЭЭГ/ВП-исследованием испытуемые выполняли ряд психологических заданий для оценки: когнитивных (прогрессивные матрицы Равена, 1936), вербальных творческих (RAT [58] в адаптации [59]) и невербальных творческих способностей (субтест Торренса “Незавершенные эскизы” по [60]), а также задание на выявление полезависимости/поленезависимости (тест “Включенные фигуры” [61]). Также испытуемые в бумажном варианте выполняли вербальный субтест Торренса “Альтернативное использование картонных коробок” по [60]. Это было, своего рода, тренировочное задание перед психофизиологическим исследованием с выполнением модифицированного AUT.
Процедура психофизиологического исследования. Испытуемые выполняли два типа задач внутри одной регистрации ЭЭГ/ВП: творческое задание (модифицированный тест альтернативного использования, AUT, [10]) и контрольное к нему задание (КАТЕГОРИИ). В творческом задании необходимо было придумать оригинальное использование какого-либо предмета (газета, кирпич, скрепка и т.д). В контрольном задании КАТЕГОРИИ испытуемому было необходимо перечислить предметы из предложенной категории (мебель, посуда, транспорт и т.д.). Пример организации отдельной пробы представлен на рис. 1. В каждой пробе в период обдумывания испытуемый должен был, в случае придуманного ответа, мысленно его проговорить и нажать на кнопку, а после предъявления знака вопроса, разрешающего ответ – озвучить ответ и оценить стратегию его возникновения (“инсайтная”, “логическая”, “трудно сказать”). Ответы и оценки испытуемых фиксировали. Для анализа использовали все пробы заданий AUT, заданные инструкцией быть оригинальным. При этом, чтобы снизить дисперсию данных в контрольной задаче – ответы, которые оценивали в задании КАТЕГОРИИ как инсайтные, исключали из анализа.
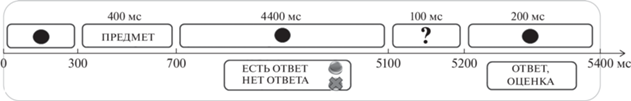
Стимулы были написаны 48 черным шрифтом (Times New Roman) на белом фоне. Суммарно испытуемым предъявляли около 160 проб, задания (АЛЬТЕРНАТИВНОЕ ИСПОЛЬЗОВАНИЕ и КАТЕГОРИИ) были организованы в блоки по 9–10 проб-попыток на один стимул и перемежались между собой во время предъявления. В начале каждого блока задания предъявляли зрительную инструкцию. Порядок начала выполнения заданий рандомизировали между испытуемыми. Перед выполнением исследования с каждым испытуемым проводили тренировочную сессию. Самоотчет испытуемых после исследования включал оценку субъективной сложности творческого и контрольного заданий по 10-балльной шкале.
Регистрация ЭЭГ. ЭЭГ регистрировали монополярно от 15 отведений (Fpz, F7, F3, Fz, F4, F8, C3, C4, T5, P3, Pz, P4, T6, O1, O2) 32-канального энцефалографа ООО Мицар, в полосе пропускания 0.53–30 Гц, частота дискретизации 500 Гц. Референтом служил объединенный ушной электрод, заземляющий электрод располагали в передне-центральном отведении, сопротивление электродов не превышало 5 кОМ, режекторный фильтр соответствовал 45–55, 905–105 Гц. Анализ ЭЭГ проводили в полосе 1.6–30 Гц. Для регистрации и анализа данных использовали программный пакет WinEEG (Пономарев В.А., Кропотов Ю.Д., № государственной регистрации 2 001 610 516 от 08.05.2001), для предъявления стимулов – программу RSYTASK (В.А. Пономарев, 2001).
Предобработка ЭЭГ-данных. Для удаления глазодвигательных артефактов использовали метод пространственной фильтрации путем обнуления соответствующих независимых компонент ЭЭГ [62–64]. Из анализа исключали фрагменты ЭЭГ, содержащие медленные волны (0–2 Гц с амплитудой выше 50 мкВ) и быстрые волны (25–35 Гц с амплитудой выше 35 мкВ), общая фильтрация ЭЭГ исключала фрагменты, содержащие волны с амплитудой больше 150 мкВ. Далее проводили визуальный анализ данных для удаления оставшихся глазодвигательных и мышечных артефактов.
Вычисление и анализ ВП. В творческом и контрольном заданиях для расчета ВП учитывали только те пробы, где испытуемый давал ответ (с нажатием кнопки), т.е. ответы, которые испытуемые обдумывали и о которых сигнализировали намеренно, без ложных нажатий и задержек. Для каждого испытуемого и каждого отведения рассчитывали индивидуальные усредненные ВП в окне от –300 до 2000 мс от начала предъявления стимула для отдельных типов стимулов (задание AUT, задание КАТЕГОРИИ – придумать использование/перечислить объекты из категории). Временны́е интервалы и зоны интереса для статистического анализа определяли по полумаксимуму разностной волны между ВП AUT и контрольного задания внутри групп и по полумаксимуму разностной волны для одних и тех же заданий в межгрупповом сравнении.
Вычисление и анализ связанной с событиями синхронизации/десинхронизации ЭЭГ. Расчет вызванной синхронизации/десинхронизации ЭЭГ производили на безартефактных пробах в интервале [–300; 2000 мс] относительно предъявления стимула. Вызванную мощность рассчитывали от 2 до 30 Гц с шагом в 0.25 Гц, с использованием вейвлет преобразования Морле [65]. Ширина материнского вейвлета составила 5 циклов. ВП при расчете связанной с событиями синхронизации/десинхронизации вычитали. Вычисляли усредненные по группам проб оценки спектральной мощности для каждого испытуемого, каждого электрода и каждой группы проб по отдельности.
Статистический анализ данных. Амплитуды ВП и показатели вызванной синхронизации/десинхронизации в группах более и менее креативных/продуктивных участников сравнивали с использованием дисперсионного анализа для повторных измерений (RM-ANOVA) для факторов ЗАДАНИЕ (АЛЬТЕРНАТИВНОЕ ИСПОЛЬЗОВАНИЕ/КАТЕГОРИИ) и ЗОНА (15 отведений ЭЭГ) и их взаимодействий. Межгрупповые сравнения амплитуд ВП и показателей вызванной синхронизации/десинхронизации проводили при помощи RM-ANOVA для внутригруппового фактора ЗОНА (15 отведений ЭЭГ) и межгруппового фактора УРОВЕНЬ ОРИГИНАЛЬНОСТИ/ПРОДУКТИВНОСТИ (2 группы) и их взаимодействий. Разделение групп участников на подгруппы проводили при помощи кластерного анализа (метод К-средних).
С использованием критерия Спирмена рассчитывали корреляции процента ответов в творческом и контрольном задании (показатели продуктивности деятельности) и результатов психологических тестов.
Межгрупповые сравнения времени принятия решения и субъективной сложности заданий проводили с использованием критерия Манна-Уитни, попарное сравнение времени ответа между творческим и контрольным заданием осуществляли при помощи критерия Вилкоксона.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Поведенческие и психологические результаты в общей группе испытуемых. Общая характеристика группы и результаты психологического тестирования, а также поведенческие показатели выполнения заданий приведены в табл. 2.
Таблица 2.
Характеристика общей группы испытуемых и поведенческие данные
| Название теста/показателя | Медиана | 25 квартиль | 75 квартиль |
|---|---|---|---|
| Прогрессивные матрицы Равена (IQ) | 112 | 104 | 120 |
| Оригинальность невербального творческого мышления (субтест Торренса “Эскизы”), шкала от 0 до 10 | 5.3 | 3.9 | 6.2 |
| Оригинальность конвергентного творческого мышления (тест отдаленных ассоциаций Медника), шкала от 0 до 1 | 0.68 | 0.57 | 0.85 |
| Оригинальность дивергентного творческого мышления (субтест “Альтернативное использование”), шкала от 0 | 4 | 2 | 6 |
| Время нахождения решения в AUT, мс* | 2219 | 1937 | 2412 |
| Время нахождения решения в задаче КАТЕГОРИИ, мс* | 1724 | 1424 | 2155 |
| Процент ответов в задаче AUT, %** | 56 | 49 | 73 |
| Процент ответов в задаче КАТЕГОРИИ, %** | 87 | 82 | 94 |
При сравнительном рассмотрении показателя “оригинальность” в трех психологических тестах творческого мышления: невербального субтеста Торренса “Эскизы” (оценка по базе ответов группы О.М. Разумниковой, г. Новосибирск), теста отдаленных ассоциаций Медника (RAT, в адаптации [59]) и субтеста Торренса “Альтернативное использование картонных коробок” [66] – не было выявлено корреляции по данному параметру. Можно предположить независимость показателей оригинальности вербального и невербального творческого мышления, и относительную независимость оригинальности ассоциативного и дивергентного мышления. При этом внутри каждого теста значимо коррелировали показатели беглости и оригинальности (Rs от 0.6 до 0.73).
Процент/количество ответов в AUT в рамках психофизиологического исследования положительно коррелировали с оригинальностью (Rs = = 0.37, t(N–2) = 2.4, p < 0.03); а также с беглостью (Rs = 0.35, t(N–2) = 2.3, p < 0.03) в тесте Медника и в “бумажном” тренировочном субтесте Торренса – “Альтернативное использование картонных коробок” (Rs = 0.33, t(N–2) = 2.21, p < 0.04).
Также было выявлено, что, чем выше поленезависимость, тем больше ответов давал человек в задании AUT (Rs = 0.4, t(N–2) = 2.7, p < 0.01) и КАТЕГОРИИ (Rs = 0.3, t(N–2) = 2.0, p < 0.04).
Результаты ЭЭГ/ВП-исследования в общей группе испытуемых. AUT vs КАТЕГОРИИ. В условиях инструкции к творческому заданию – “быть оригинальными” – не выявлено различий при нахождении ответа в творческом задании (AUT), по сравнению с контрольным (КАТЕГОРИИ) в общей группе испытуемых (44 чел.) по данным связанной с событиями синхронизации/десинхронизации ЭЭГ.
При этом метод ВП оказался более чувствительным и различия ВП наблюдались как в более ранних, так и в более поздних компонентах при сравнении AUT vs КАТЕГОРИИ (табл. 3, рис. 2).
Таблица 3.
Различия вызванных потенциалов (ВП) в сравнении творческого и контрольного заданий в общей группе испытуемых
| Интервал, мс, ~ компонент ВП |
Статистические эффекты | Зоны различий | Физиологические эффекты |
|---|---|---|---|
| 196–236, P200 | F(1,43) = 6.7, p < 0.02 | F7, F3, Fz, F4, F8, C3, C4, T5, P3, Pz, P4, T6, O1, O2 | Тв < K |
| 372–424, N300(400) | ЗИ: F(1,43) = 12.6, p < 0.001 | Теменные области: T5, P3, Pz, P4, T6, O1, O2 | Тв > K |
| 632–708, P600 | ЗИ: F(1,43) = 6.6, p < 0.02 | Лобные области: Fpz, F7, F3, Fz, F4, F8 | Тв > K |
Рис. 2.
Вызванные потенциалы (ВП) при нахождении ответа в творческом задании AUT (черная линия) и контрольном задании КАТЕГОРИИ (серая линия) в общей группе участников (n = 44). AUT – тест альтернативного использования. Fpz–O2 – электроды; по оси х – время (мс), по оси y – амплитуда ВП (мкВ). Вертикальные пунктирные линии – начало/конец предъявления стимула (длительность интервала – 400 мс). * – компонент значимых различий между состояниями.
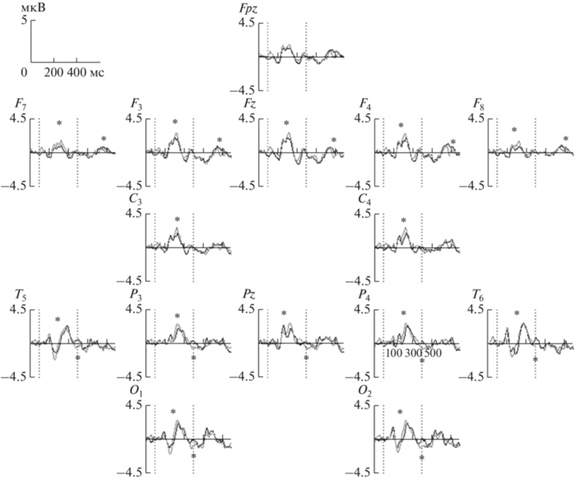
Как можно видеть на рис. 2 амплитуда компонента P200 в лобных областях была ниже при нахождении ответа в творческом задании, а в затылочных областях можно отметить несколько большую латентность компонента P200 в творческом задании. Негативный компонент N300 был меньше по амплитуде в творческом задании, а амплитуда позднего позитивного компонента Р600 – выше при нахождении ответа в творческом задании.
Подгруппы испытуемых по результатам поведенческих и психологических данных. Так как в двух психологических тестах дивергентного творческого мышления (субтесты Торренса “Эскизы” и “Альтернативное использование картонных коробок”) нами не были получены корреляции между показателями оригинальности, можно предположить, что исследуемая в тестах с разной модальностью “оригинальность” характеризует разные аспекты этого параметра и может отразиться в разных особенностях физиологических реакций.
Таким образом, для дальнейшего анализа общая группа участников была разделена на подгруппы по: (i) оригинальности невербального мышления (субтест Торренса “Эскизы”, классический и часто встречающийся вариант разделения испытуемых на подгруппы); (ii) оригинальности вербального дивергентного мышления (субтест Торренса – “Альтернативное использование картонных коробок”); (iii) продуктивности творческого и нетворческого мышления (количество ответов) в физиологическом исследовании при выполнении теста альтернативного использования (AUT) и контрольного задания.
Анализ ЭЭГ/ВП-данных в группах испытуемых с разным уровнем вербальной и невербальной оригинальности. Разделение на подгруппы по уровню оригинальности. Общая группа участников для оценки физиологических показателей была независимо разделена на подгруппы исходя из: (i) оригинальности невербального мышления (оцененного на основании субтеста Торренса “Эскизы”); (ii) оригинальности вербального дивергентного мышления (субтест Торренса “Альтернативное использование картонных коробок”). Для разделения использовали кластерный анализ методом K-средних (отдельно для индекса невербальной оригинальности и отдельно для индекса вербальной оригинальности). Таким образом, общая группа была дважды разделена на подгруппы, условно обозначенные как “высоко оригинальные” и “низко оригинальные”. В табл. 4 приведены оценки оригинальности и количество человек для данных подгрупп. У двоих испытуемых отсутствовали оценки психологических тестов, и они были исключены из последующего рассмотрения.
Таблица 4.
Показатели оригинальности творческого мышления в группах низко- и высоко-оригинальных участников
| Показатель | Низко оригинальные: M (25–75Q) (n) | Высоко оригинальные: M (25–75Q) (n) |
|---|---|---|
| Оригинальность невербального мышления (субтест Торренса “Эскизы”) | 3.15 (2.6–4.3) (n = 17) | 6.0 (5.3–6.8) (n = 25) |
| Оригинальность дивергентного мышления (субтест Торренса “Альтернативное использование”) | 2.5 (2–4) (n = 24) | 6 (5–8) (n = 18) |
Физиологические данные. Внутри и межгрупповых различий ВП при выполнении задания AUT и КАТЕГОРИИ испытуемыми с разным уровнем оригинальности в вербальном и невербальном дивергентных психологических творческих заданиях – выявлено не было.
Более чувствительным в данном случае оказался метод связанной с событием синхронизации/десинхронизации ЭЭГ.
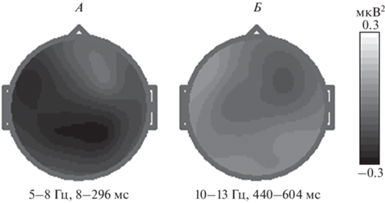
Были выявлены: (i) межгрупповые различия при решенииAUTв частотном диапазоне 5–8 Гц на интервале до 296 мспосле предъявления стимула: F(1,40) = 6.0, p < 0.02 (эффект фактора УРОВЕНЬ НЕВЕРБАЛЬНОЙ ОРИГИНАЛЬНОСТИ). Мощность в данном диапазоне была ниже в группе с большей невербальной оригинальностью. Зоны различий: F7, F3, Fz, C3, Pz, P4, T6, O2 (рис. 3, А); (ii) в группе с меньшей оригинальностью ВЕРБАЛЬНОГО ДИВЕРГЕНТНОГО ТВОРЧЕСКОГО МЫШЛЕНИЯ наблюдаются меньшие значения мощности в AUT vs КАТЕГОРИИ в частотном диапазоне 10–13 Гц на интервале 440–604 мс после предъявления стимула (эффект фактора ЗАДАНИЕ: F(1,22) = 5.7, p < 0.03. Зоны различий: Fpz, F3, Fz, F4, F8, C3, C4, P3, Pz, P4 (рис. 3, Б)).
Рис. 3.
Разностные топограммы мощности электроэнцефалограммы (ЭЭГ) при нахождении решения в творческом задании (АЛЬТЕРНАТИВНОЕ ИСПОЛЬЗОВАНИЕ). А – различия между группами с высокой и низкой оригинальностью невербального мышления в диапазоне частот 5–8 Гц, в интервале 8–296 мс после предъявления стимула; Б – различия между нахождением ответа в творческом (тест альтернативного использования – AUT) vs контрольном задании (КАТЕГОРИИ) в частотном диапазоне 10–13 Гц на интервале 440–604 мс после предъявления стимула в группе с низкой оригинальностью вербального творческого мышления.
Анализ ЭЭГ/ВП-данных в группах испытуемых с разным уровнем продуктивности. Для разделения группы участников на подгруппы согласно результатам выполнения задания в физиологическом исследовании использовали процент ответов в творческом (AUT) и контрольном (КАТЕГОРИИ) задании. Разделение по продуктивности деятельности проводилось на основании двух переменных – процента ответов в творческом задании и процента ответов в контрольном задании. Количество возможных подгрупп для разделения определяли визуально при помощи иерархического кластерного анализа (метод Варда), как количество ветвей на середине максимального расстояния объединения. При данном подходе было выявлено, что группа испытуемых может быть разделена на 2 кластера. В дальнейшем для кластеризации использовали метод K-средних и обозначили выделенные группы как более- и менее- продуктивных участников (табл. 5).
Таблица 5.
Процент ответов в творческом и контрольном задании в физиологическом исследовании в группах более и менее продуктивных участников
| Показатель | Менее продуктивные (n = 24): M (25–75Q) | Более продуктивные (n = 24): M (25–75Q) |
|---|---|---|
| Процент ответов в контрольном задании (КАТЕГОРИИ) | 83 (74–87) | 94 (89–97) |
| Процент ответов в творческом задании (AUT) | 49 (44–55) | 74 (69–80) |
Сравнения ВП в группах участников с большей и меньшей продуктивностью при выполнении задания в физиологическом исследовании (AUT vs КАТЕГОРИИ). В группе более продуктивных участников наблюдаются различия ВП при сравнении AUT vs КАТЕГОРИИ в следующих интервалах: (i) 360−408 мс: эффект фактора ЗАДАНИЕ: F(1,19) = 5.0, p < 0.05, зоны отличий – Fpz, F3, Fz, C3, C4, T5, P3, Pz, P4, T6, O1, O2 – на данном интервале амплитуда при нахождении ответа в творческом задании была выше, чем при нахождении ответа в контрольном задании. Данные различия, вероятно, связаны с амплитудой и латентностью компонента N300, более “негативном” в контрольном задании. (ii) 616–680 мс: эффект взаимодействия факторов ЗАДАНИЕ на ЗОНА: F(14,266) = 5.7, e(G-G) = = 0.26, p < 0.001, зоны отличий – Fpz, F7, F3, Fz, F4, F8, C3 – на данном интервале амплитуда при нахождении ответа в творческом задании была выше, чем при нахождении ответа в контрольном задании. Эти различия соотносятся с большей амплитудой положительного компонента P600 в творческом задании относительно контрольного (рис. 4, А).
Рис. 4.
Вызванные потенциалы (ВП) при нахождении ответа в творческом задании (черная линия) и контрольном задании (серая линия) в группе более продуктивных участников (А) и менее продуктивных участников (Б). Fpz–O2 – электроды; по оси х – время (мс), по оси y – амплитуда ВП (мкВ). Вертикальные пунктирные линии – время предъявления стимула (400 мс). Знак * обозначает интервал значимых различий между состояниями.
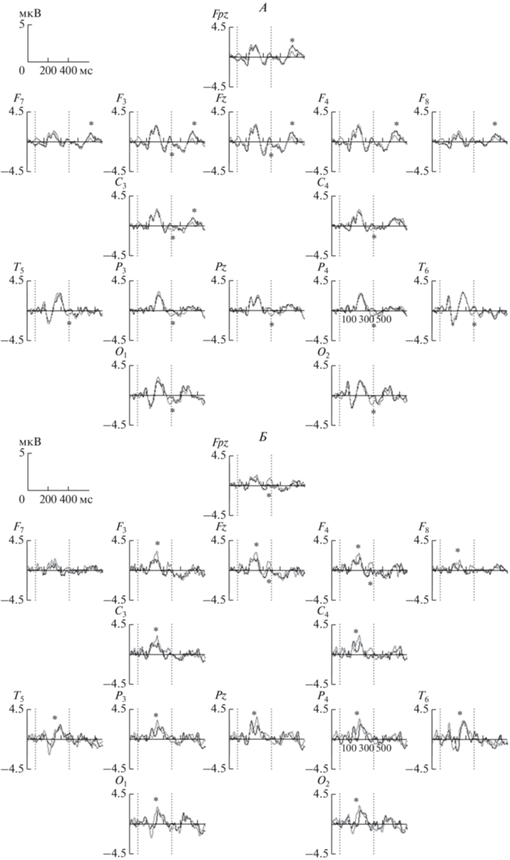
В группе менее продуктивных участников различия ВП при сравнении AUT vs КАТЕГОРИИ наблюдаются в следующих интервалах: (i) 192–232 мс: эффект фактора ЗАДАНИЕ: F(1,23) = 6.9, p < 0.02, зоны отличий – F3–O2 – на данном интервале амплитуда при нахождении ответа в творческом задании была ниже, чем при нахождении ответа в контрольном задании. Эти различия соотносятся с меньшей амплитудой положительного компонента P200 в творческом относительно контрольного задания. (ii) 360–408 мс: эффект взаимодействия факторов ЗАДАНИЕ на ЗОНА: F(14,322) = 3.5, e(G–G) = 0.17, p < 0.05, зоны отличий – Fpz, Fz, F4, T5. На данном интервале амплитуда в лобных областях была выше при выполнении контрольного задания по сравнению с творческим, что, по-видимому, может быть связано с большей латентностью отрицательных и положительных пиков ВП в творческом задании по сравнению с контрольным в данной группе участников (рис. 4, Б).
Сравнения связанной с событиями синхронизации/десинхронизации ЭЭГ в группах участников с большей и меньшей продуктивностью в физиологическом исследовании. В группе более продуктивных участников наблюдается значимый эффект взаимодействия факторов ЗАДАНИЕ на ЗОНА на показатели вызванной мощности ЭЭГ во временнóм интервале 356–564 мс после предъявления стимула в диапазоне частот 7–9 Гц: F(14,266) = = 3.9, e(G–G) = 0.28, p < 0.01. В лобных и центральных зонах – Fpz, F7, F3, Fz, F4, С3 наблюдается десинхронизация (уменьшение мощности) ЭЭГ, а в отведениях P4 и O2 – синхронизация ЭЭГ при нахождении ответа в творческом задании по сравнению с контрольным (рис. 5).
Рис. 5.
Разностные частотно-временны́е карты и топограмма различий (диапазон частот 7–9, интервал 356–564 мс после предъявления стимула) между нахождением ответов в творческом задании в сравнении с нахождением неинсайтных ответов в контрольном задании. Fpz-C3 – электроды; по оси х – время (мс), вертикальными пунктирными линиями обозначен временнóй интервал предъявления стимула (400 мс); по оси y – анализируемые частоты (2–30 Гц); цветовая шкала указывает на разностную мощность ЭЭГ: черный цвет – снижение мощности/десинхронизация; белый цвет – увеличение мощности/синхронизация.
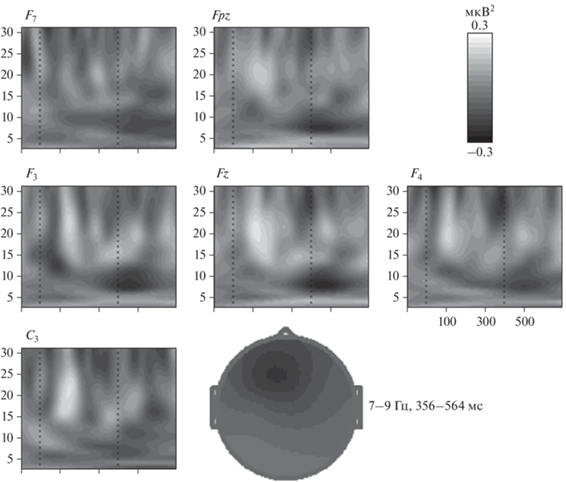
В группе менее продуктивных участников значимых различий, вызванной синхронизации/десинхронизации ЭЭГ при нахождении ответов в творческом задании по сравнению с контрольным, не выявлено.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Психологические данные, поведенческие результаты и данные самоотчетов. В целом, в исследовании приняла участие группа молодых взрослых с высокими показателями когнитивных способностей (медиана IQ = 112), вполне успешно справившаяся с заданиями и адекватно оценившая свои результаты выполнения (оригинальность ответов – 5 [25–75%: 4–6]). Было выяснено, что между более и менее продуктивными испытуемыми отсутствовали отличия в оценке субъективной сложности задания и в оценке оригинальности собственных ответов. Cложность творческого задания – 7 [25–75%: 6–8], контрольного задания – 5 [25–75%: 4–6], в общей группе оценивалась, как разная (T = 6.5, Z = 2.9, p < 0.004).
Творческие задания, в отличие от контрольных, включают в себя поиск в широком ассоциативном поле, дивергентное мышление, оценку и выбор ответа, что заведомо предполагает большую когнитивную нагрузку и необходимость больших усилий для решения данных задач [67]. Ранее в наших исследованиях был выявлен специфический эффект творческого характера деятельности в отличие от влияния сложности задания на биоэлектрическую активность мозга [68, 69]. В психологических и психофизиологических исследованиях показано, что самооценка усилий, приложенных к выполнению заданий выше у тех участников, которые давали больше ответов [70]. В исследовании [67] было показано, что более творческие участники, определенные согласно опроснику творческих достижений, тратят больше усилий (объективно измеряемых с помощью реокардиографии) при выполнении заданий на дивергентное мышление (альтернативное использование) и достигают лучших результатов, чем менее творческие участники. В нашем исследовании между группами более и менее продуктивных участников различий субъективных оценок сложности творческого задания не было, также как и корреляции между количеством ответов и самооценкой сложности задания, что, может быть, связано с рандомизацией блоков заданий – когда творческая деятельность сменялась когнитивным заданием по перечислению категорий и наоборот. А субъективная сложность задания “Альтернативное использование картонных коробок”, выполняемого в блоке психологического тестирования в течение 3-х мин, отрицательно коррелировала с количеством ответов, данных участниками в физиологическом исследовании.
Из рассматриваемых личностных предикторов творческой деятельности в нашем исследовании можно выделить показатель поленезависимости. При этом поленезависимость (большие коэффициенты, свидетельствующие о поленезависимости) была положительно взаимосвязана с общей вербальной беглостью – количеством ответов как в творческом, так и в контрольном заданиях.
Данные ВП. Оценка влияния продуктивности деятельности. Анализ ВП оказался более чувствительным для выделения различий между творческим и контрольным заданием в общей группе испытуемых, чем анализ связанной с событием синхронизации/десинхронизации ЭЭГ. Вместе с тем разделение на подгруппы участников выявило различия ВП только при разделении по продуктивности деятельности, но не оригинальности.
Наиболее ранние различия ВП в общей группе испытуемых были связаны с меньшей амплитудой компонента P200 (196–236) при восприятии стимула в творческом задании по сравнению с контрольным. Компонент P200 часто рассматривается как показатель активации внимания, в том числе во время чтения, при манипулировании контекстом [71] и специфически связанный с семантическими процессами при чтении [72]. На амплитуду данного компонента может влиять частотность слова [73, 74]: амплитуда P200 ниже на более частое слово, а также контекст [75]: амплитуда P200 увеличивается в эмоциональном контексте. В нашем исследовании амплитуда P200 была ниже при нахождении ответа в творческом задании по сравнению с контрольным – в общей группе испытуемых и в группе испытуемых с меньшей продуктивностью деятельности. У испытуемых, нашедших большее количество ответов, амплитуды P200 в творческом и контрольном задании – не различались. Мы можем предположить, что участники с низкой продуктивностью деятельности задействовали больше ресурсов внимания при выполнении задания, чем участники с высокой продуктивностью. Различия в поздних компонентах ВП – N300 (N400) и P600, также весьма специфичны для групп с низкой и высокой продуктивностью и влияют на результаты при оценке ВП в общей группе. В группе с низкой продуктивностью наблюдается бóльшая негативность компонента N300 в лобных зонах – при решении творческой задачи, в то время как в группе с высокой продуктивностью негативность компонента N400 больше в центральных и теменных областях коры при решении контрольной задачи, а в лобных областях выше амплитуда позднего позитивного компонента P600.
В литературе обсуждается различие между лобным и теменным компонентом N400, где лобный компонент FN400 рассматривается как отражение распознавания слова на основе знакомости или же он функционально идентичен центро-теменному компоненту N400, отражающему семантическую обработку [76, 77]. В нашем исследовании данные компоненты дифференцировали группы менее и более продуктивных участников. Лобный компонент FN400 был более негативным в творческом задании в группе с низкой продуктивностью, а центрально-теменной компонент N400 был более выражен в контрольном задании (перечислить объекты из категории) в группе с большей продуктивностью – вероятно, в связи с активацией семантической сети категориального идентификатора.
Необходимо подчеркнуть, что большинство исследований ВП построены на основе восприятия речевого материала – слов и предложений с буквальным и метафорическим смыслом. При этом отмечается бóльшая негативность N400 при восприятии метафор в сравнении с буквальным значением или при восприятии оригинального/бессмысленного способа употребления предмета [16, 31], но амплитуда этой негативности модулируется знаниями воспринимающего [78]. В целом, N400 интерпретируется как показатель больших усилий по извлечению семантической информации и интеграции новых концепций, представленных в метафорических или бессмысленных фразах [79]. В нашем исследовании при генерации, а не при восприятии ответов, бóльшая амплитуда компонента N400 в контрольном задании, вероятно, связана с активацией памяти и семантических сетей предъявленной категории. Группа более продуктивных участников давала значимо больше ответов и в контрольном задании (94% от всех проб).
В группе более продуктивных участников при выполнении творческого задания (AUT) выделялся лобный компонент P600, амплитуда которого, кроме того, положительно коррелировала с процентом ответов в творческом задании (рис. 4, A) в общей группе испытуемых. Амплитуды поздних компонентов ВП (P600) были чувствительны к индивидуальным различиям креативности в исследовании [16], а также различались между бессмысленным (неподходящим) и осмысленным (подходящим) способом употребления предмета при его оценке [31] и оценке новизны идеи [80]. В нашем исследовании бóльшая амплитуда P600, возможно, связана с оценкой испытуемыми собственных идей – их новизны и употребимости, при этом на данном этапе (около 600 мс), мы можем говорить о первичной оценке, так как время внутренней вербализации ответа и нажатия на кнопку в творческом задании было около 2219 мс.
В общей группе испытуемых компонент P600 в творческом задании также выделяется, как и в группе более продуктивных участников (рис. 2), что важно для дальнейших сравнений с исследованиями без разделения групп.
Анализ связанной с событиями синхронизации/ десинхронизации ЭЭГ. Оценка продуктивности и оригинальности творческого мышления. При анализе связанной с событиями синхронизации/десинхронизации различия между творческим и контрольным заданием выявлялись только при разделении группы по показателям оригинальности или продуктивности и в ряде сравнений данных групп. Все различия были связаны с десинхронизацией ЭЭГ при нахождении ответа в творческом задании (придумать альтернативное использование предмета).
В группе более продуктивных участников наблюдаются различия между творческим и контрольным заданием в лобных областях в частотном диапазоне 7–9 Гц на интервале 356–564 мс после предъявления стимула. Десинхронизация в низкочастотном α1-диапазоне свидетельствует о большей активации лобных областей при нахождении решения в творческом, по сравнению с контрольным, задании. Связанная с событием десинхронизация отмечается, как при создании идей альтернативного использования [23], их оценке [81] решении инсайтных задач [82]. В ряде работ подчеркивается, что нахождение творческого решения (более отдаленного, оригинального и т.п.) сопровождается синхронизацией ЭЭГ в α-диапазонах [5, 33, 34], однако в зависимости от стадии творческого процесса [21]. В нашем исследовании на придумывание идеи давалось сравнительно мало времени, и оно не контролировалось самим участником, что в целом может вызывать большее напряжение и активацию. То есть, творческая деятельность в нашей модельной ситуации требовала быстрого включения в деятельность и принятия решения, что характеризовалось паттерном активации у более продуктивных участников.
Можно отметить разное вовлечение активации лобных и теменных структур при нахождении решения в творческом задании, по сравнению с контрольным при разделении участников по продуктивности и оригинальности вербального дивергентного мышления. А именно: нахождение решения в творческом задании при сравнении с контрольным в группе с большей продуктивностью сопровождается описанной выше десинхронизацией в θ- и низкочастотном α1-диапазонах ЭЭГ (7–9 Гц на интервале 356–564 мс) в лобных областях с максимумом различий мощности в левом полушарии (рис. 5). А в группе с меньшей оригинальностью вербального дивергентного мышления – десинхронизацией в высокочастотном α2-диапазоне (10–13 Гц на интервале 440–604 мс) с максимумом различий мощности в лобных областях правого полушария (рис 3, Б). Возможно, в группе с меньшей оригинальностью вербального творческого мышления при нахождении ответа в физиологическом исследовании активировалась жесткая смысловая сеть способов обычного употребления предмета (вовлекая области дорсолатеральной префронтальной коры), для преодоления которой требовалась бы синхронизация в высокочастотном α2-диапазоне [17, 21, 34]. Следует подчеркнуть, что данные группы (выделенные по продуктивности и оригинальности) частично совпадают, т.е. участник мог быть высокопродуктивным (давал много ответов на творческое задание в физиологическом исследовании), но низкооригинальным по данным психологического исследования.
Особенности невербального творческого мышления также отражаются в процессе поиска ответа в творческом задании: в группе с высокой оригинальностью невербального творческого мышления на ранних этапах восприятия стимула в творческом задании (до 296 мс) наблюдается бóльшая десинхронизация в θ-диапазоне 5–8 Гц в теменных областях и лобных областях левого полушария. Возможно, данные ранние отличия связаны с включением зрительных представлений и активацией мультимодальных сенсорных областей в группы с большими невербальными творческими способностями.
ЗАКЛЮЧЕНИЕ
В настоящей работе представлены новые данные, позволяющие оценить как ранние, так и более поздние этапы поиска ответа в творческом задании альтернативного использования по параметрам ВП и связанной с событиями синхронизации/десинхронизации ЭЭГ. Выявлено, что индивидуальные показатели продуктивности деятельности (количества ответов в творческом и контрольном задании) отражаются в характеристиках ВП при нахождении ответа в задании альтернативного использования. В группе с большей продуктивностью нахождение ответа в творческом задании характеризовалось различиями семантических компонентов ВП: меньшей амплитудой негативного компонента N300–N400 в центральных и теменных отведениях и большей амплитудой компонента P600 в лобных областях по сравнению с контрольным заданием. В группе с меньшей продуктивностью в творческом задании наблюдалась меньшая амплитуда компонента P200 и бóльшая негативность N300–N400 в лобных областях. Эти различия могут отражать активацию семантических сетей при поиске ответа у продуктивных участников и меньшую эффективность когнитивного контроля в группе с низкой продуктивностью.
Анализ связанной с событиями синхронизации/десинхронизации ЭЭГ выявил влияние как продуктивности деятельности, так и оригинальности мышления на корреляты нахождения способов альтернативного использования: высокая оригинальность невербального творческого мышления приводила к большей активации теменных областей на этапе восприятия стимула (до 296 мс) в творческом задании, что, вероятно, связано с механизмами зрительного представления и воображения. Низкая оригинальность вербального творческого мышления отражалась в десинхронизации ЭЭГ в α2-диапазоне (10–13 Гц) на 440–604 мс в лобных областях правого полушария при поиске ответа в творческом задании по сравнению с контрольным, что может рассматриваться как фиксация на более распространенном (неоригинальном) способе использования объекта. В то время как бóльшая продуктивность деятельности была связана с десинхронизацией в α1-диапазоне (7–9 Гц) в лобных областях левого полушария в сопоставимом временнóм интервале 356–564 мс, свидетельствующей об активации вербальной семантической сети.
Стоит отметить, что все выделенные отличия имеют достаточно раннюю латентность (до 1000 мс) и отражают основные этапы восприятия стимула и поиска решения.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Этическим комитетом Института эволюционной физиологии и биохимии им. И.М. Сеченова РАН (Санкт-Петербург).
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа поддержана РНФ (грант № 22-28-02073).
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Список литературы
Fink A., Benedek M., Grabner R.H. Creativity meets neuroscience: experimental tasks for the neuroscientific study of creative thinking // Methods. 2007. V. 42. № 1. P. 68.
Dietrich A., Kanso R. A review of EEG, ERP, and neuroimaging studies of creativity and insight // Psychol. Bull. 2010. V. 136. № 5. P. 822.
Runco M.A., Jaeger G.J. The standard definition of creativity // Creat. Res. J. 2012. V. 24. № 1. P. 92.
Шемякина Н.В., Данько С.Г., Нагорнова Ж.В. и др. Динамика спектров мощности и когерентности ритмических компонентов ЭЭГ при решении вербальной творческой задачи преодоления стереотипа // Физиология человека. 2007. Т. 33. № 5. С. 14. Shemyakina N.V., Danko S.G., Nagornova Zh.V. et al. Changes in the power and coherence spectra of the EEG rhythmic components during solution of a verbal creative task of overcoming a stereotype // Human Physiology. 2007. V. 33. № 5. P. 524.
Camarda A., Salvia É., Vidal J. et al. Neural basis of functional fixedness during creative idea generation: An EEG study // Neuropsychologia. 2018. V. 118. Pt A. P. 4.
Shemyakina N.V., Nagornova Z.V. EEG “Signs” of Verbal Creative Task Fulfillment with and without Overcoming Self-Induced Stereotypes // Behav. Sci. (Basel). 2019. V. 10. № 1. P. 17.
Sternberg R.J. The Nature of Creativity // Creat. Res. J. 2006. V. 18. № 1. P. 87.
Bechtereva N.P. The usefulness of psychophysiology in the maintenance of cognitive life // Int. J. Psychophysiol. 2009. V. 73. № 2. P. 83.
Christensen P.R., Guilford J.P., Merrifield P.R., Wilson R.C. Alternate Uses / Sheridan Psychological Service: Beverly Hills, 1960.
Guilford J.P. The nature of human intelligence. Mc-Graw-Hill, 1967. 538 p.
Plucker J.A., Makel M.C. Assessment of creativity / The Cambridge handbook of creativity. Publisher: Cambridge University Press, 2010. P. 48.
Barbot B., Hass R.W., Reiter-Palmon R. Creativity assessment in psychological research: (Re)setting the standards // Psychology of Aesthetics, Creativity, and the Arts. 2019. V. 13. № 2. P. 233.
Hao X., Geng F., Wang T. et al. Relations of Creativity to the Interplay Between High-order Cognitive Functions: Behavioral and Neural Evidence // Neuroscience. 2021. V. 473. P. 90.
Bitsch F., Berger P., Fink A. et al. Antagonism between brain regions relevant for cognitive control and emotional memory facilitates the generation of humorous ideas // Sci. Rep. 2021. V. 11. № 1. P. 10685.
Lu K., Qiao X., Yun Q., Hao N. Educational diversity and group creativity: Evidence from fNIRS hyperscanning // NeuroImage. 2021. V. 243. P. 118564.
Abraham A., Rutter B., Hermann C. Conceptual expansion via novel metaphor processing: an ERP replication and extension study examining individual differences in creativity // Brain Lang. 2021. V. 221. P. 105007.
Stevens C.E., Zabelina D.L. Classifying Creativity: Applying Machine Learning Techniques to Divergent Thinking EEG Data // NeuroImage. 2020. V. 219. P. 116990.
Chrysikou E.G., Jacial C., Yaden D.B. et al. Differences in Brain Activity Patterns During Creative Idea Generation Between Eminent and Non-eminent Thinkers // NeuroImage. 2020. V. 220. P. 117011.
Perchtold-Stefan C.M., Papousek I., Rominger C. et al. Humor comprehension and creative cognition: Shared and distinct neurocognitive mechanisms as indicated by EEG alpha activity // NeuroImage. 2020. V. 213. P. 116695.
Lu K., Teng J., Hao N. Gender of partner affects the interaction pattern during group creative idea generation // Exp. Brain Res. 2020. V. 238. № 5. P. 1157.
Rominger C., Papousek I., Perchtold C.M. et al. Creativity is associated with a characteristic U-shaped function of alpha power changes accompanied by an early increase in functional coupling // Cogn. Affect. Behav. Neurosci. 2019. V. 19. № 4. P. 1012.
Petersen T., Puthusserypady S. Assessing tDCS Placebo Effects on EEG and Cognitive Tasks / 41st Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC). 2019. P. 4509.
Agnoli S., Zanon M., Mastria S. et al. Predicting response originality through brain activity: An analysis of changes in EEG alpha power during the generation of alternative ideas // NeuroImage. 2020. V. 207. P. 116385.
Hertenstein E., Waibel E., Frase L. et al. Modulation of creativity by transcranial direct current stimulation // Brain Stimul. 2019. V. 12. № 5. P. 213.
Wang X., Duan H., Kan Y. et al. The creative thinking cognitive process influenced by acute stress in humans: an electroencephalography study // Stress. 2019. V. 22. № 4. P. 472.
Kraus B., Cadle C., Simon-Dack S. EEG alpha activity is moderated by the serial order effect during divergent thinking // Biol. Psychol. 2019. V. 145. P. 84.
Wang M., Hao N., Ku Y. et al. Neural correlates of serial order effect in verbal divergent thinking // Neuropsychologia. 2017. V. 99. P. 92.
Heinonen J., Numminen J., Hlushchuk Y. Default Mode and Executive Networks Areas: Association with the Serial Order in Divergent Thinking // Plos One. 2016. V. 11. № 9. P. e0162234.
Marmpena M., Dimitriadis S.I., Thakor N., Bezerianos A. Phase to amplitude coupling as a potential biomarker for creative ideation: An EEG study / 38th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC). 2016. P. 383.
enedek M., Jauk E., Fink A. et al. To create or to recall? Neural mechanisms underlying the generation of creative new ideas. // NeuroImage. 2014. V. 88. № 100. P. 125
Kröger S., Rutter B., Hill H. et al. An ERP study of passive creative conceptual expansion using a modified alternate uses task // Brain Res. 2013. V. 1527. P. 189.
Jauk E., Benedek M., Neubauer A.C. Tackling creativity at its roots: Evidence for different patterns of EEG alpha activity related to convergent and divergent modes of task processing // Int. J. Psychophysiol. 2012. V. 84. № 2. P. 219.
Fink A., Graif B., Aljoscha C. Brain correlates underlying creative thinking: EEG alpha activity in professional vs. novice dancers // NeuroImage. 2009. V. 46. № 3. P. 854.
Fink A., Grabner R.H., Benedek M. et al. The creative brain: Investigation of brain activity during creative problem solving by means of EEG and FMRI // Hum. Brain Mapp. 2009. V. 30. № 3. P. 734.
Pfurtscheller G. Quantifikation of ERD and ERS in the time domain / Event-related desynchronization // Handbook of Elektroencephalic and Clinical Neurophysiology. 1999. V. 6. P. 89.
Guilford J.P. Creativity // American Psychologist. 1950. V. 5. P. 444.
Runco M.A., Acar S. Divergent thinking as an indicator of creative potential // Creat. Res. J. 2012. V. 24. № 1. P. 66.
Martindale C., Hines D. Creativity and cortical activation during creative, intellectual and EEG feedback tasks // Biol. Psychol. 1975. V. 2. № 3. P. 91.
Martindale C., Hines D., Mitchell L. et al. EEG alpha asymmetry and creativity // Pers. Individ. Differ. 1984. V. 5. P. 77.
Mölle M., Marshall L., Wolf B. et al. EEG complexity and performance measures of creative thinking // Psychophysiology. 1999. V. 36. № 1. P. 95.
Martindale C., Armstrong J. EEG complexity and performance measures of creative thinking // Psychophysiology. 1999. V. 36. № 1. P. 94.
Jausovec N., Jausovec K. EEG activity during the performance of complex mental problems // Int. J. Psychophysiol. 2000. V. 36. № 1. P. 73.
Razumnikova O.M. Creativity related cortex activity in the remote associates task // Brain Res. Bull. 2007. V. 73. № 1–3. P. 96.
Gruzelier J. A theory of alpha/theta neurofeedback, creative performance enhancement, long distance functional connectivity and psychological integration // Cogn. Process. 2009. V. 10. № 1. P. 101.
Karakaş S. A review of theta oscillation and its functional correlates // Int. J. Psychophysiol. 2020. V. 157. P. 82.
Шемякина Н.В., Потапов Ю.Г., Нагорнова Ж.В. Динамика частотного ЭЭГ во время эскизирования в экологических условиях и выполнения невербальных творческих задач профессиональным художником: лонгитюдное case study // Физиология человека. 2022. Т. 48. № 5. С. 26. Shemyakina N.V., Potapov Y.G., Nagornova Zh.V. Dynamics of EEG Frequency Structure During Sketching in Ecological Conditions and Non-Verbal Creative Tasks Fulfillment by a Professional Artist: Case Study // Human Physiology. 2022. V. 48. № 5. P. 506.
Cruz-Garza J.G., Sujatha Ravindran A., Kopteva A.E. et al. Characterization of the Stages of Creative Writing With Mobile EEG Using Generalized Partial Directed Coherence // Front. Hum. Neurosci. 2020. V. 14. P. 577651.
Williams C.C., Kappen M., Hassall C.D. et al. Thinking theta and alpha: Mechanisms of intuitive and analytical reasoning // Neuroimage. 2019. V. 189. P. 574.
Eisma J., Rawls E., Long S. et al. Frontal midline theta differentiates separate cognitive control strategies while still generalizing the need for cognitive control // Sci. Rep. 2021. V. 11. № 1. P. 14641.
Herweg N.A., Solomon E.A., Kahana M.J. Theta Oscillations in Human Memory // Trends Cogn Sci. 2020. V. 24. № 3. P. 208.
Kwiatkowski J., Vartanian O., Martindale C. Creativity and speed of mental processing // Empir. Stud. Arts. 1999. V. 17. № 2. P. 187.
Bechtereva N.P. Korotkov A.D., Pakhomov S.V. et al. PET study of brain maintenance of verbal creative activity // Int. J. Psychophysiol. 2004. V. 53. № 1. P. 11.
Flaherty A.W. Frontotemporal and dopaminergic control of idea generation and creative drive // J. Comp. Neurol. 2005. V. 493. № 1. P. 147.
Beaty R.E. Creativity and the default network: A functional connectivity analysis of the creative brain at rest // Neuropsychologia. 2014. V. 64. P. 92.
Beaty R.E. The neuroscience of musical improvisation // Neurosci. Biobehav. Rev. 2015. V. 51. P. 108.
Bitsch F., Berger P., Nagels A. et al. The role of the right temporo-parietal junction in social decision-making // Hum. Brain Mapp. 2018. V. 39. № 7. P. 3072.
Wertz C.J., Chohan M.O., Flores R.A., Jung R.E. Neuroanatomy of creative achievement // Neuroimage. 2020. V. 209. P. 116487.
Mednick T., Mednick F.M. Creative Thinking and Level of Intelligence // J. Creat. Behav. 1967. V. 1. P. 428.
Воронин А.Н., Галкина Т.В. Диагностика вербальной креативности (адаптация теста Медника) / Методы психологической диагностики. М.: “Институт психологии РАН”, 1994. № 2. С. 40.
Туник Е.Е. Тест Торренса / Диагностика креативности. СПб.: ИМАТОН, 1998. 170 с.
Witkin H.A., Oltman P.K., Raskin E., Karp S.A. A manual for the embedded figures tests / Consulting Psychologists Press, 1971. 32 p.
Vigario R.N. Extraction of ocular artefacts from EEG using independent component analysis // Electroencephalogr. Clin. Neurophysiol. 1997. V. 103. № 3. P. 395.
Jung T.P., Makeig S., Westerfield M. et al. Removal of eye activity artifacts from visual event-related potentials in normal and clinical subjects // Clin. Neurophysiol. 2000. V. 111. № 10. P. 1745.
Терещенко Е.П., Пономарев В.А., Кропотов Ю.Д., Мюллер А. Сравнение эффективности различных методов удаления артефактов морганий при анализе количественной электроэнцефалограммы и вызванных потенциалов // Физиология человека. 2009. Т. 35. № 2. С. 124. Tereshchenko E.P., Ponomarev V.A., Kropotov Yu.D., Müller A. Comparative efficiencies of different methods for removing blink artifacts in analyzing quantitative electroencephalogram and event-related potentials // Human Physiology. 2009. V. 35. № 2. P. 241.
Tallon-Baudry C., Bertrand O. Oscillatory gamma activity in humans and its role in object representation // Trends Cogn. Sci. 1999. V. 4. № 3. P. 151.
Туник Е.Е. Диагностика креативности. Тест Е. Торренса / Адаптированный вариант. СПб.: Речь, 2006. С. 176.
Silvia P.J., Beaty R.E., Nusbaum E.C. et al. Creative motivation: creative achievement predicts cardiac autonomic markers of effort during divergent thinking // Biol. Psychol. 2014. V. 102. P. 30.
Shemyakina N.V., Nagornova Zh.V., Danko S.G. What do we measure – creativity or task complexity? // Psychophysiology. 2009. V. 46. № 1. P. 51.
Данько С.Г., Шемякина Н.В., Нагорнова Ж.В., Старченко М.Г. Сравнение воздействия субъективной сложности и творческого характера вербальной деятельности на спектральные показатели мощности ЭЭГ // Физиология человека. 2009. Т. 35. № 3. С. 132. Danko S.G., Shemyakina N.V., Nagornova Zh.V., Starchenko M.G. Comparison of the effects of the subjective complexity and verbal creativity on EEG spectral power parameters // Human Physiology. 2009. V. 35. № 3. P. 381.
von Helversen B., Gendolla G.H.E., Winkielman P. et al. Exploring the hardship of ease: Subjective and objective effort in the ease-of-processing paradigm // Motiv. Emot. 2008. V. 32. № 1. P. 1.
Yang C.L., Zhang H., Duan H., Pan H. Linguistic focus promotes the ease of discourse Integration processes in reading comprehension: evidence from event-related potentials // Front. Psychol. 2019. V. 9. P. 2718.
Landi N., Perfetti C.A. An electrophysiological investigation of semantic and phonological processing in skilled and less-skilled comprehenders // Brain Lang. 2007. V. 102. № 1. P. 30.
Dambacher M., Kliegl R., Hofmann M., Jacobs A.M. Frequency and predictability effects on event-related potentials during reading // Brain Res. 2006. V. 1084. № 1. P. 89.
Lee H., Lee Y., Tae J., Kwon Y. Advantage of the go/no-go task over the yes/no lexical decision task: ERP indexes of parameters in the diffusion model // PLoS One. 2019. V. 14. № 7. P. e0218451.
Donahoo S.A., Pfeifer V., Lai V.T. Cursed concepts: new insights on combinatorial processing from ERP correlates of swearing in context // Brain Lang. 2022. V. 226. P. 105079.
Bridger E.K., Bader R., Kriukova O. et al. The FN400 is functionally distinct from the N400 // NeuroImage. 2012. V. 63. № 3. P. 1334.
Stróżak P., Abedzadeh D., Curran T. Separating the FN400 and N400 potentials across recognition memory experiments // Brain Res. 2016. V. 1635. P. 41.
Jończyk R., Kremer G.E., Siddique Z., van Hell J.G. Engineering creativity: Prior experience modulates electrophysiological responses to novel metaphors // Psychophysiology. 2020. V. 57. № 10. P. e13630.
Rutter B., Kröger S., Hill H. et al. Can clouds dance? Part 2: an ERP investigation of passive conceptual expansion // Brain Cogn. 2012. V. 80. № 3. P. 301.
Tang S., Jia L., Ren J. et al. Reactive control in evaluating appropriately or inappropriately novel ideas: Evidence from electrophysiological measures // Psychophysiology. 2022. V. 59. № 7. P. e14010.
Rataj K., Nazareth D.S., van der Velde F. Use a Spoon as a Spade? Changes in the Upper and Lower Alpha Bands in Evaluating Alternate Object Use // Front. Psychol. 2018. V. 9. P. 1941.
Sandkühler S., Bhattacharya J. Deconstructing insight: EEG correlates of insightful problem solving // PLoS One. 2008. V. 3. № 1. P. e1459.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека