

Физиология человека, 2023, T. 49, № 1, стр. 28-41
Особенности нейрофункциональной организации рабочей памяти для базовых характеристик зрительного пространства у мужчин и женщин
Е. С. Михайлова 1, *, **, А. Б. Кушнир 1, Н. Ю. Мошникова 1
1 ФГБУН Институт высшей нервной деятельности и нейрофизиологии РАН
Москва, Россия
* E-mail: esmikhailova@mail.ru
** E-mail: mikhailovaes@gmail.com
Поступила в редакцию 30.04.2022
После доработки 09.06.2022
Принята к публикации 24.06.2022
- EDN: APBDJU
- DOI: 10.31857/S0131164622600379
Аннотация
В группе 38 испытуемых (19 женщин) в задаче рабочей памяти (РП) обнаружены половые различия нейронных механизмов сопоставления ориентационных характеристик зрительного пространства. У мужчин бо̀льшая по сравнению с женщинами чувствительность амплитуды ранней затылочно-височной негативности N150 к совпадению/несовпадению текущей и удерживаемой в памяти ориентаций говорит об эффективном раннем детектировании изменений ориентаций. Более высокая амплитуда компонента Р200 затылочно-височных связанных с событием потенциалов и центральной позитивности 400–500 мс у мужчин по сравнению с женщинами рассматривается как показатель более высокого уровня селективного внимания к ориентациям и потенциальной возможности удержания в памяти бо̀льшего объема информации о пространственных характеристиках среды. В работе установлена статистически значимая связь времени реакции выполнения задачи РП и оценок стратегий навигационного поведения по данным опросников. Предполагается, что эффективная зрительно-пространственная РП на ориентации является одним из важных биологических факторов, лежащих в основе более успешной навигационной стратегии, учитывающей глобальные пространственные ориентиры.
Пол является одним из важных биологических факторов, определяющих особенности нейрофункциональной организации как перцептивных функций, так и когнитивных процессов разной степени сложности [1, 2]. Зрительно-пространственные способности являются той частью поведения человека, где различия между мужчинами и женщинами проявляются наиболее отчетливо. Преимущество мужчин экспериментально подтверждено как для базовых характеристик зрительного восприятия, таких как острота, порог контрастной чувствительности, оценка направления движения, ориентационная чувствительность [3], так и для более сложных действий. Например, в тесте трехмерного ментального вращения превосходство мужчин остается неизменно высоким на протяжении десятков лет [4]. Успешность выполнения этого теста положительно коррелирует с выполнением других пространственных задач, например, скоростью выучивания сложных маршрутов на картах [5] и эффективностью обучения навигации в виртуальном лабиринте [6]. Существуют убедительные экспериментальные подтверждения тесной связи механизмов ментального вращения и зрительно-пространственной рабочей памяти (РП) [7]. Понимание нейрофизиологических основ половых различий зрительной РП может предоставить сведения для более обоснованного понимания гендерной специфичности зрительно-пространственного поведения. Интерес к РП на ориентационные характеристики зрительной среды определяется результатами наших, ранее проведенных исследований, выполненных с регистрацией вызванных потенциалов (ВП) при идентификации базовых и наклонных ориентаций [8–10], а также при моделировании распределенных внутримозговых дипольных источников компонентов ВП. Было показано, что на ранних стадиях обработки информации (компоненты Р100 и N150 ВП) у мужчин, по сравнению с женщинами, значимо выше уровень корковой активации в затылочной, височной и теменной областях коры.
Половые различия в пространственном поведении обнаружены у различных видов млекопитающих как в естественных, так и лабораторных условиях. Самки и самцы не только по-разному используют пространство, но привлекают для навигационного поведения разную информацию [5, 11, 12]. Так, например, самки диких кенгуровых крыс более чувствительны к отдельным уникальным характеристикам объектов, а самцы чаще обращаются к геометрии окружающего пространства [11, 13]. По данным [14], при обучении крыс в радиальном лабиринте у самцов производительность снижалась при изменении геометрии помещения, а у самок – при изменении положения отдельных объектов в помещении. Сходные результаты были получены в исследованиях на людях [15–17].
Для понимания нейронных механизмов половых различий поведения в пространстве интересна модель когнитивной карты, предложенная L.F. Jacobs и F. Schenk [18]. Эта модель “параллельных карт” (“parallel map model”) постулирует, что когнитивная карта представляет собой интеграцию двух параллельных компонентов, которые построены с использованием двух различных классов сигналов: сигналов направления (“directional cues”) и позиционных сигналов (“positional cues”). Эти два вида карт поддерживаются различными субобластями гиппокампа и связанными с ними структурами головного мозга [19, 20]. К сигналам направления относятся градиентные сенсорные сигналы, такие как запах и свет, географическая информация, геометрические подсказки и отдаленные ориентиры, которые хоть и описывают окружающую среду, но не предоставляют точной информации о местоположении. Позиционные сигналы – это ближайшие точечные объекты, которые могут предоставить более точную информацию о расположении. Пространственное представление об окружающей среде формируется путем объединения локальных позиционных карт в более крупные карты, построенные на основе информации о направлениях. Таким образом, ориентационная чувствительность является важной составляющей навигационного поведения и может определять его стратегию.
В настоящей работе исследовали функциональную организацию РП на ориентации у мужчин и женщин с привлечением метода связанных с событием потенциалов (ССП). Применение “n-back” модели РП мы рассматривали как более чувствительный тест для оценки половых различий этой функции по сравнению с использованной нами ранее моделью отставленного сопоставления образца с эталоном.
Задача “n-back” считается валидным тестом при исследовании нейронных коррелятов РП и, в частности, ее гендерной специфики в связи с возможностью изменять объем удерживаемой в памяти информации и доступом к высокоуровневым процессам управления, например, обновлению содержания памяти [21]. Кроме того, мы анализировали половые различия в использовании разных пространственных стратегий по данным заполнения опросников, чтобы выяснить, существует ли связь между этими стратегиями и половыми различиями выполнения задачи РП на ориентации в исследуемой группе испытуемых.
МЕТОДИКА
В исследовании принимали участие 38 испытуемых (19 мужчин и 19 женщин) c нормальным или скорректированным до нормы зрением, без неврологических или психиатрических расстройств. Средний возраст мужчин 24.5 ± 0.9 лет, женщин – 24.1 ± 0.9 лет. Большинство испытуемых (29 чел.) были в возрастном диапазоне от 19 до 26 лет, 9 чел. – от 27 до 33 лет. Все испытуемые имели высшее или незаконченное высшее образование. Большинство из них (35 чел.) имели естественно-научное, медицинское или гуманитарное образование, и только у трех человек (2 мужчин и 1 женщина) было техническое образование. Исследование проводили в утреннее и дневное время c 9 до 15 ч.
Стимулы представляли собой прямоугольные контрастные решетки из горизонтальных (0 град), вертикальных (90 град) и наклонных (45 град) черных линий (рис. 1, A), представленных на светло-сером фоне (1 цикл/град).
Рис. 1.
Условия эксперимента. А – зрительные стимулы – ориентационные решетки горизонтальной, вертикальной и наклонной (45 град) ориентации. Б – схема расположения 128 регистрирующих электродов на поверхности головы. Цифрами обозначены кластеры электродов, в которых проводили анализ компонентов связанных с событием потенциалов (ССП): 1 – лобный, 2 – центральный, 3 – теменной, 4 – височный и 5 – затылочный. В – временнáя последовательность событий в одном блоке эксперимента – сличение текущего (тестового) и удерживаемого в памяти (эталонного) стимулов.
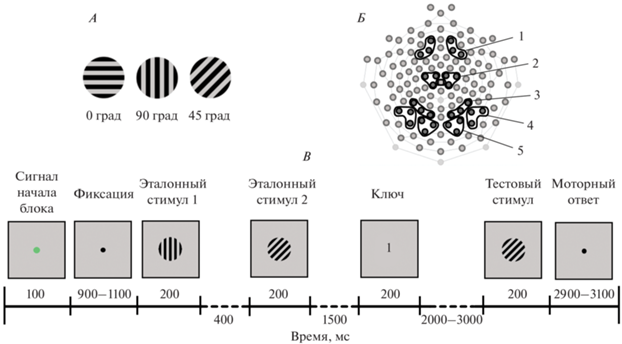
Процедура эксперимента. Во время исследования испытуемый сидел в удобном кресле в затемненной звукозаглушенной комнате на расстоянии 120 см от экрана монитора Dell E1911c (диагональ 17 дюймов, частота обновления экрана 60 Гц). Размер стимула на экране монитора составлял 6 угл. град. Освещенность на уровне глаз испытуемого – 3 лк. Горизонтальные и вертикальные края монитора были закрыты круглой апертурой. На рис. 1, Б представлена временнáя схема эксперимента. Каждый блок состоял из следующих событий: зеленая точка длительностью 100 мс, при появлении который испытуемый должен был сосредоточиться и фиксировать взгляд в центре экрана; черная фиксационная точка, длительность которой в случайном порядке изменялась от 1500 до 1700 мс; первый эталонный стимул (200 мс), интервал 400 мс, второй эталонный стимул (200 мс); после периода удержания информации (1500 мс) на экране появлялся “ключ” – цифра 1 или 2, сигнализирующая, с каким из эталонов необходимо сравнивать тестовый стимул. После “ключа” через интервал времени 2000–3000 мс на экране появлялся тестовый стимул (200 мс). Интервал между двумя последовательными блоками в серии варьировал от 2900 до 3100 мс. Общая длительность эксперимента – от 21 до 24 мин в зависимости от длительности варьирующих по времени интервалов между стимулами. Предъявление стимулов и регистрацию поведенческих ответов испытуемого проводили с помощью программы E-Prime 2.0 (Psychology Software Tools, Inc., США).
Испытуемому предъявляли 144 пары стимулов, в 50% случаев тестовая ориентация совпадала с эталонной, в 50% случаев – не совпадала. “Ключи” 1 и 2 использовали с равной вероятностью. В сценарий входили 4 варианта сочетания эталонного и тестового стимулов и “ключа”: 1) “ключ 1” и совпадение первого эталонного и тестового стимулов; 2) “ключ 2” и совпадение второго эталонного и тестового стимулов; 3) “ключ 1” и несовпадение первого эталонного и тестового стимулов; 4) “ключ 2” и несовпадение второго эталонного и тестового стимулов. Последовательность совпадений и несовпадений была псевдорандомизированной.
Испытуемого просили ответить на вопрос, совпадают или не совпадают тестовая и соответствующая “ключу” эталонная ориентации. При совпадении испытуемый должен был нажать на клавишу “1” выносной клавиатуры, при несовпадении – на клавишу “2”. С каждым испытуемым перед экспериментом проводили тренировочную серию для ознакомления с экспериментальной задачей и обучения моторному навыку. В качестве стимулов использовали изображения букв. Выносную клавиатуру располагали перед испытуемым. Нажатие осуществляли в удобной для него манере: указательными пальцами обеих рук или указательным и средним пальцем одной руки. В ходе исследования делали перерыв 5–10 мин для отдыха испытуемого, проверки и коррекции сопротивления электродов.
Все принимавшие участие в эксперименте испытуемые заполняли опросники для оценки стратегии навигационного поведения (Lawton’s Wayfinding Strategy Scale) [22]. Определяли (в баллах) использование в навигационном поведении двух различных стратегий: 1) стратегии с использованием метрических характеристик и глобальных пространственных ориентиров (“ориентационная стратегия”, которая в статье [22] обозначена как Orientation strategy) и 2) стратегии с использованием топографии ближних меток на местности (“стратегия маршрута”, Route strategy).
Регистрация и анализ ССП. Регистрацию электроэнцефалограммы (ЭЭГ) проводили с помощью 128-канального электроэнцефалографа (Electrical Geodesics Inc., США) с использованием шлема Hydro Cel Geodesic Sensor Net. Размер шлема подбирали индивидуально с учетом окружности головы испытуемого. Частота квантования сигнала составляла 500 Гц. Согласно рекомендации производителя электроэнцефалографа сопротивление электродов не превышало 50 кОм. При записи в качестве референтного электрода использовали сенсор, расположенный в отведении вертекс (Cz). Данные ЭЭГ обрабатывали off-line в программе NetStation 4.5.4 (Electrical Geodesics Inc., США). При off-line обработке ЭЭГ в качестве референтного выбирали усредненный электрод. В ходе препроцессинга фильтрованные (0.5–45 Гц) записи ЭЭГ каждого испытуемого сегментировали на отрезки длительностью 1300 мс, содержащие интервалы 300 мс до предъявления тестового стимула и 1000 мс после, и программно исключали реализации с глазными движениями, амплитуда которых превышала 80 µV. Далее, при дополнительном визуальном анализе удаляли фрагменты записи с двигательными, глазодвигательными и электромиографическими артефактами. Для каждого испытуемого по безартефактным отрезкам ЭЭГ проводили усреднение ССП на тестовый стимул только для правильных ответов. В группе мужчин усреднение проводили по 39.7 ± 2.4 реализациям, в группе женщин – по 44.0 ± 2.1. Коррекцию базовой линии проводили по 300 мс отрезку ЭЭГ до предъявления эталонного стимула. После усреднения для каждого испытуемого было получено по два усредненных ССП – для совпадения и несовпадения эталонной и тестовой ориентаций.
Амплитуду компонентов индивидуальных ССП измеряли в программе NetStation 4.5.4. Для каждого компонента замеряли адаптивный минимум/ максимум: программно регистрировали среднее значение амплитуды в 4-миллисекундном интервале (2 мс до и 2 мс после пика компонента). При статистической обработке анализировали амплитуду компонентов ССП, усредненную по кластерам электродов, расположенных над затылочной, височной, теменной, центральной и лобной областями коры. Кластеры схематично приведены на рис. 1, Б. Измеряли амплитуду следующих компонентов ССП. В затылочном, височном, теменном кластерах – Р100 (временнóе окно 60–120 мс), N150 (115–200 мс), Р200 (200–250 мс), N270 (250-320 мс) и P300 (310–370 мс). В центральном – N90 (60–125 мс), Р150 (125–200 мс), N200 (190–250 мс), а для позднего позитивного отклонения измеряли среднюю амплитуду в интервале от 400 до 500 мс. Такой способ измерения амплитуды поздних компонентов ССП рекомендован в [23]. Во фронтальном кластере измеряли пиковые амплитуды N90 (60–125 мс), Р150 (125–200 мс), N200 (190–250 мс) и P250 (250–320 мс), N320 (310–380 мс) и P400 (340–420 мс). В окончательный анализ ССП включены данные 31 чел. (16 мужчин и 15 женщин), ЭЭГ-записи которых не содержали большого количества артефактов при высокой степени представленности всех компонентов.
Статистический анализ амплитуд компонентов индивидуальных ССП, усредненных по группам электродов, и поведенческих показателей (время реакции (ВР) и точность) проводили с использованием дисперсионного анализа ANOVA RM (метод повторных измерений). При внутригрупповых post-hoc сопоставлениях для поправки на множественные сравнения использовали Тьюки-тест (Tukey test). Для межгрупповых сопоставлений использовали T-test. Для определения связи между поведенческими характеристиками и оценками по шкалам стратегий навигации использовали коэффициент корреляции по Пирсону с учетом нормальности их распределения (по критерию Шапиро-Уилка) и принадлежности этих оценок к категории величин, оцениваемых в интервальной шкале.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Анализ поведенческих характеристик выполнения задачи сличения тестовой и эталонной ориентаций. Проведен дисперсионный анализ RM ANOVA (метод повторных измерений) ВР и точности (вероятность правильных ответов) с учетом факторов Условие (совпадение и несовпадение эталонной и тестовой ориентаций) и Пол. Установлено, что ВР больше при несовпадении эталонной и тестовой ориентаций, эффект Условия: F(1.36) = 41.06, p = 0.0001, $\eta _{{\text{p}}}^{{\text{2}}}$ = 0.053. Влияние Пола статистически незначимо, хотя приведенные средние по группам значениям ВР свидетельствуют о тенденции к более низким значениям у мужчин (725.68 ± 39.53 и 844.41 ± 43.35 мс при совпадении и несовпадении, соответственно) по сравнению с женщинами (810.56 ± 55.67 и 952.15 ± 73.25 мс). Для точности не было выявлено значимого влияния фактора Пола. И у мужчин, и у женщин точность выполнения задания достаточно высокая: в группе мужчин – 0.95 ± 0.02, в группе женщин – 0.91 ± 0.02. Эти результаты проиллюстрированы диаграммами на рис. 2, А.
Рис. 2.
Поведенческие характеристики выполнения задачи сопоставления тестовой и эталонной ориентаций в группах мужчин и женщин. А – усредненные по группам время реакции (ВР) и точность. Цифрами обозначены: 1 – черные закрашенные столбики – значения в группе мужчин при совпадении эталонной и тестовой ориентаций, 2 – черные заштрихованные – значения в группе мужчин при несовпадении, 3 – серые закрашенные – значения в группе женщин при совпадении, 4 – серые заштрихованные – значения в группе женщин при несовпадении. Приведены средние по группе значения и ошибки среднего. Достоверность различий: *** – p < 0.005 (критерий Тьюки). Б – графики зависимости ВР (мс) выполнения задания от оценок по шкалам стратегий навигации (а, б – стратегия “ориентирования”; в, г – стратегия “маршрута”) в группах мужчин (а, в) и женщин (б, г). Цифрами обозначены условия: 1 – совпадения эталонной и тестовой ориентаций и 2 – несовпадения эталонной и тестовой ориентаций.
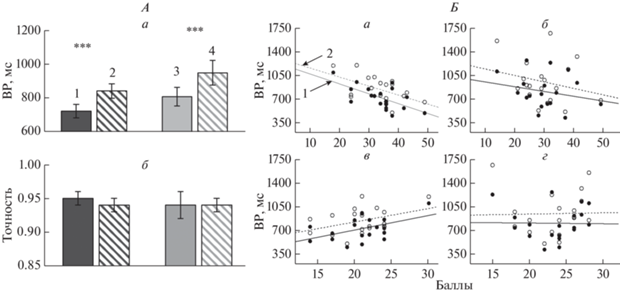
В объединенной группе испытуемых показаны бо́льшие оценки для стратегии “ориентирования” (31.86 ± 1.27) по сравнению со стратегией “маршрута” (22.13 ± 0.63), Т = 6.82, df = 36, p = = 0.0001. Для стратегии “ориентирования” средние показатели выше у мужчин (33.95 ± 1.76) по сравнению с женщинами (29.79 ± 1.76), Т = 1.62, df = 36, p = 0.10; для стратегии “маршрута” средние показатели выше у женщин (23.32 ± 0.86) по сравнению с мужчинами (20.95 ± 0.86), Т = –1.95, df = 36, p = 0.06, но эти межгрупповые различия статистически незначимы.
Проведен регрессионно-корреляционный анализ поведенческих характеристик выполнения задачи сличения эталонной и тестовой ориентаций и оценок стратегий навигации. В группе мужчин коэффициенты корреляции Пирсона между ВР и оценками стратегии “ориентирования” были отрицательными, т.е., при более высоких баллах для стратегии “ориентирования” задание выполнялось быстрее, чем при более низких. Коэффициенты корреляции, рассчитанные отдельно для трех эталонных ориентаций (горизонталь, вертикаль, наклон), располагались в диапазоне от –0.502 до –0.644 (0.001 < р < 0.05). Для значений ВР, усредненных по трем эталонным ориентациям, эта зависимость описывалась уравнением линейной регрессии y = 1226.37 – 14.75x, r = = –0.63, p = 0.004 (для совпадения эталона и теста), и y = 1301.47 – 13.46x, r = –0.53, p = 0.02 (для несовпадения). Эта зависимость представлена на рис. 2, Б, фрагмент а. ВР выполнения задания (для совпадения эталона и теста) положительно коррелировала с оценками по шкале стратегии “маршрута” (r = 0.474, p = 0.04), рис. 2, Б, фрагмент в. То есть, чем выше значения по шкале стратегия “маршрута”, тем медленнее выполняется задание. В группе женщин значимые корреляции между оценками шкал навигации и ВР выполнения задания отсутствовали, что проиллюстрировано на рис. 2, Б, фрагменты б и г.
Анализ ССП при выполнении задачи сличения тестовой и эталонной ориентаций. Результаты дисперсионного анализа амплитуды компонентов ССП представлены в табл. 1. Усредненные по группам мужчин и женщин ССП на тестовые стимулы и диаграммы средних амплитуд компонентов ССП в каудальных и передне-центральных кластерах представлены на рис. 3 и 4.
Таблица 1.
Результаты дисперсионного анализа амплитуды компонентов связанных с событием потенциалов (ССП)
| Кластер электродов | Волна | Фактор | df | F | p | $\eta _{{\text{p}}}^{2}$ |
|---|---|---|---|---|---|---|
| Фронтальный | N90 | Условие × Полушарие | 1.29 | 3.88 | 0.05 | 0.118 |
| Условие × Полушарие × Пол | 1.29 | 7.13 | 0.012 | 0.197 | ||
| N200 | Полушарие | 1.29 | 5.60 | 0.025 | 0.162 | |
| Условие × Полушарие× Пол | 1.29 | 11.46 | 0.002 | 0.283 | ||
| P200 | Условие × Полушарие × Пол | 1.29 | 4.14 | 0.05 | 0.125 | |
| N320 | Условие × Пол | 1.29 | 8.25 | 0.008 | 0.22 | |
| Центральный | N90 | Условие × Полушарие × Пол | 1.29 | 9.15 | 0.005 | 0.24 |
| P400–500 | Пол | 1.29 | 6.36 | 0.02 | 0.18 | |
| Височный | N150 | Условие | 1.29 | 3.99 | 0.05 | 0.121 |
| Условие × Пол | 1.29 | 4.12 | 0.05 | 0.124 | ||
| P200 | Пол | 1.29 | 3.98 | 0.05 | 0.121 | |
| Полушарие × Пол | 1.29 | 4.02 | 0.05 | 0.122 | ||
| Теменной | N270 | Условие | 1.29 | 5.87 | 0.05 | 0.168 |
| P300 | Условие | 1.29 | 9.93 | 0.005 | 0.255 | |
| Полушарие | 1.29 | 4.56 | 0.05 | 0.136 | ||
| Затылочный | N150 | Условие | 1.29 | 8.60 | 0.01 | 0.229 |
| Условие × Полушарие | 1.29 | 4.00 | 0.05 | 0.121 | ||
| Условие × Пол | 1.29 | 5.69 | 0.05 | 0.164 | ||
| P200 | Пол | 1.29 | 3.94 | 0.05 | 0.12 |
Рис. 3.
Половые различия связанных с событием потенциалов (ССП) на тестовый стимул в теменном, височном и затылочном кластерах электродов в задаче рабочей памяти на ориентации. А – усредненные по группам мужчин и женщин ССП правого и левого полушарий для условий совпадения и несовпадения эталонной и тестовой ориентаций теменных (а), височных (б) и затылочных (в) областей. Слева приведены данные по левому полушарию, справа – по правому. Цифрами обозначены: 1 – черная сплошная линия – ССП в группе мужчин при совпадении ориентаций, 2 – черная пунктирная – ССП в группе мужчин при несовпадении, 3 – серая сплошная – ССП в группе женщин при совпадении, 4 – серая пунктирная – ССП в группе женщин при несовпадении. Вертикальной линией обозначен момент предъявления тестового стимула. Обозначены пики компонентов Р90, N150, P200, N270 и P300. Б – средние значения амплитуды компонентов ССП в теменном (а), височном (б) и затылочном (в) кластерах правого полушария при совпадении и несовпадении эталонной и тестовой ориентаций в группах мужчин и женщин. Цифрами обозначены: 1 – черные закрашенные столбики – средние амплитуды в группе мужчин при совпадении ориентаций, 2 – черные заштрихованные – средние амплитуды в группе мужчин при несовпадении, 3 – серые закрашенные – средние амплитуды в группе женщин при совпадении, 4 – серые заштрихованные – средние амплитуды в группе женщин при несовпадении. Приведены средние по группе значения и ошибки среднего. Достоверность различий: * – p < 0.05 (критерий Тьюки).
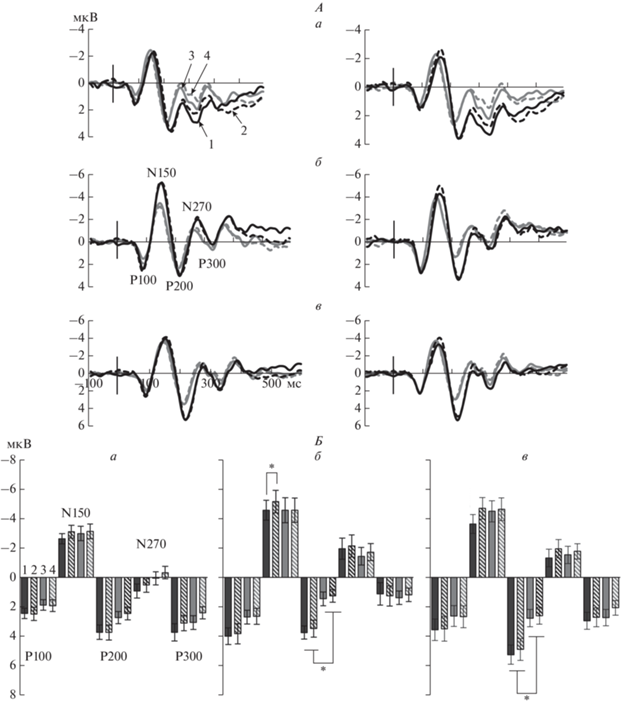
Рис. 4.
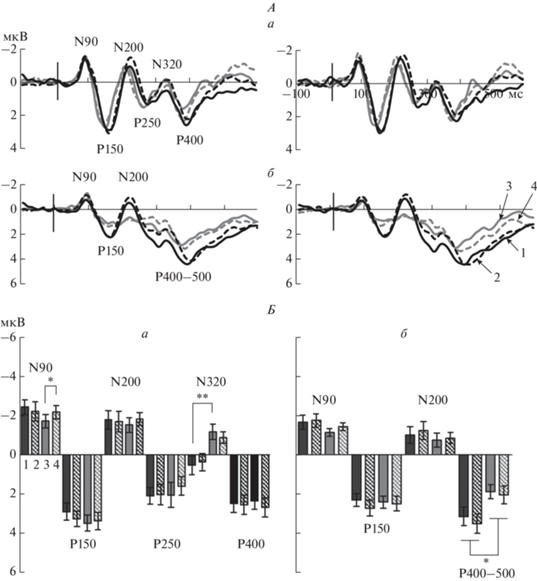
Половые различия связанных с событием потенциалов (ССП) на тестовый стимул во фронтальном и центральном кластерах электродов в задаче рабочей памяти на ориентации. А – усредненные по группам мужчин и женщин ССП правого и левого полушарий для условий совпадения и несовпадения эталонной и тестовой ориентаций во фронтальном (а) и центральном (б) кластерах электродов. Слева приведены данные по левому полушарию, справа – по правому. Обозначены пики компонентов N90, P150, N200, P250, N320, P400 и Р400–500. Б – средние значения амплитуды компонентов ССП во фронтальном (а) и центральном (б) кластерах правого полушария при совпадении и несовпадении эталонной и тестовой ориентаций в группах мужчин и женщин. Достоверность различий: * – p < 0.05, ** – p < 0.01 (критерий Тьюки). Остальные обозначения см. рис. 3.
В затылочном кластере электродов связанные с полом эффекты выявлены для амплитуды компонентов N150 и P200. Для амплитуды N150 они проявлялись как значимый эффект Условия (p = = 0.006) в виде бо̀льшей амплитуды N150 при несовпадении тестовой и эталонной ориентаций по сравнению с их совпадением. Влияние Условия зависело от Пола (Условие × Пол, р = 0.027): только в группе мужчин амплитуда N150 больше при несовпадении эталонного и тестового стимулов по сравнению с совпадением (p = 0.003). Близкое к значимому взаимодействие Условие × × Полушарие (р = 0.053) проявлялось как наличие значимых различий между условиями совпадения и несовпадения только в правом полушарии (p = 0.0002) и как бо̀льшая амплитуда N150 в правом полушарии по сравнению с левым (p = = 0.0002) при условии совпадения эталонной и тестовой ориентаций.
Для амплитуды компонента Р200 эффект Пола значим (р = 0.029). Межгрупповое сопоставление амплитуды Р200, усредненной через два Условия и два Полушария, показало более высокую амплитуду в группе мужчин по сравнению с женщинами (T = 2.29, df = 29, p = 0.029). На рис. 3, А, в приведены усредненные по группам мужчин и женщин ССП затылочного кластера правого и левого полушарий для условий совпадения и несовпадения тестового и эталонного стимулов. Отчетливо видно превышение амплитуды Р200 в группе мужчин по сравнению с женщинами. Этот эффект проиллюстрирован на диаграммах средних значений амплитуды Р200 в группах мужчин и женщин (рис. 3, Б, в).
В височном кластере электродов связанные с полом различия обнаружены для амплитуды компонентов N150 и P200. Для компонента N150 основной эффект Условия близок к значимому (р = = 0.053). Влияние Условия было неодинаково в группах мужчин и женщин, что подтверждается взаимодействием Условие × Пол (р = 0.05). По результатам post-hoc сопоставлений, только в группе мужчин есть значимое превышение амплитуды N150 при несовпадении эталонного и тестового стимулов относительно их совпадения (р = 0.03). Сходно с затылочным, в височном кластере электродов амплитуда компонента Р200 выше в группе мужчин по сравнению с женщинами (эффект Пола значим, р = 0.012). Межгрупповое сопоставление амплитуды Р200, усредненной через два Условия и два Полушария, показало более высокую амплитуду в группе мужчин по сравнению с женщинами (T = 2.67, df = 29, p = 0.012). Этот эффект проявляется в правом полушарии (Полушарие × Пол, р = 0.052), что видно на ССП (рис. 3, А, б) и диаграммах (рис. 3, Б, б). В правом полушарии амплитуда Р200 выше в группе мужчин по сравнению с женщинами, что подтверждено post-hoc сопоставлением (p = 0.03).
В теменном кластере электродов значимые влияния Пола как в виде основного эффекта, так и в виде его взаимодействия с другими факторами не выявлены. Обнаружен эффект Условия для амплитуды компонентов N270 (р = 0.02) и P300 (р = 0.023) в виде бо̀льшей амплитуды при совпадении по сравнению с несовпадением эталонной и тестовой ориентаций. Амплитуда компонента Р300 выше в правом полушарии, что соответствует эффекту Полушария (р = 0.01). Сходно с затылочными и височными кластерами, в теменном амплитуда компонента Р200 больше в группе мужчин, но различия не достигают уровня значимости (рис. 3, А, а; рис. 3, Б, а).
В центральном кластере электродов для ранней негативности N90 показано тройное взаимодействие Условие × Полушарие × Пол (р = 0.005), что проявлялось как правополушарное преобладание амплитуды N90 при условии совпадения эталонной и тестовой ориентации (р = 0.04) в группе женщин. Основной эффект Пола (р = 0.02) выявлен для амплитуды медленной позитивности в интервале 400–500 мс после начала стимула. Более высокая амплитуда этой позитивности у мужчин по сравнению с женщинами видна на усредненных ССП (рис. 4, А, б) и на диаграммах средней амплитуды в этом интервале (рис. 4, Б, б). Межгрупповые различия амплитуды позитивности в окне 400–500 мс, усредненной через два Условия и два Полушария, значимы (T = 2.52, df = 29, p = 0.02).
Во фронтальном кластере электродов влияние Пола обнаружено как двойные или тройные взаимодействия. Для амплитуды N90 взаимодействие Условие × Полушарие × Пол (p = 0.012) проявлялось как более высокая амплитуда при несовпадении по сравнению с совпадением эталонной и тестовой ориентаций (p = 0.02) в правом полушарии в группе женщин, но не в группе мужчин. Для амплитуды N200 тройное взаимодействие Условие × Полушарие × Пол (p = 0.002) проявлялось в виде преобладания активности правого полушария в ситуации совпадения эталона и теста (р = 0.02) у мужчин и в ситуации несовпадения (р = 0.003) у женщин. Для амплитуды N320 обнаружено двойное взаимодействие Условие × Пол (p = 0.008). Компонент N320 выше у женщин по сравнению с мужчинами при условии совпадения эталонной и тестовой ориентаций (p = 0.005).
Для понимания возможной поведенческой значимости обнаруженных половых различий амплитуды компонентов ССП, зарегистрированных в постцентральных зрительных областях, мы провели корреляционный анализ по Спирмену амплитуды этих компонентов и ВР, как показателя эффективности выполнения задачи. В объединенной группе значимая корреляция получена только для амплитуды Р200 в ССП правого полушария при совпадении эталонного и тестового стимулов: r = –0.39, р = 0.03. При анализе в группах значимые корреляции получены только в группе мужчин. Самый высокий уровень отрицательной связи с ВР показан для амплитуды Р200 в каудальных кластерах правого полушария (условие совпадения эталона и теста): r = –0.67, р = 0.004 и r = –0.68, р = 0.004 в затылочном и височном кластерах, соответственно. В группе женщин значимые корреляции амплитуды компонента Р200 и ВР отсутствовали.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В проведенном нами исследовании установлено, что нейронные механизмы обнаружения различий между текущей и кратковременно удерживаемой в РП ориентациями у мужчин и у женщин различны.
Анализ ССП выявил половые различия в зрительной области коры, где локализованы нейронные структуры, ответственные за восприятие и анализ ориентаций. Они относились к раннему этапу переработки информации – временнóму окну компонента N150 и проявлялись как взаимодействие Пол × Условие. Только в группе мужчин амплитуда N150 оказалась выше при несовпадении по сравнению с совпадением. Аналогичный результат показан для амплитуды компонента N150 в височной коре правого полушария.
По данным ряда исследований при выполнении задач зрительной РП процессы, разворачивающиеся в диапазоне N150, отражают не только восприятие зрительного сигнала, но и его удержание и извлечение из памяти [24–26]. В настоящей работе мы не только подтвердили результат о значимости ранних процессов в зрительных областях коры для обнаружения изменения текущей ориентации относительно удерживаемой в памяти [27], но дополнили его сведениями о связи с полом. Можно полагать, что преимущество мужчин в раннем детектировании несовпадения текущей и удерживаемой в памяти ориентаций связано с особенностями восприятия этого вида зрительной информации. Как показано нами ранее, в задаче дискриминации ориентаций у мужчин по сравнению с женщинами выше амплитуда ранних компонентов ВП в проекционных областях, шире область локализации их дипольных источников, захватывающая затылочную, теменную и височные области коры [8, 9]. Более высокая чувствительность зрительной коры мозга мужчин к ориентационным характеристикам подтверждается данными функциональной магнитно-резонансной томографии, зарегистрированной при выполнении задачи идентификации ориентаций линий [28].
Для амплитуды компонента Р200 дисперсионный анализ выявил основной эффект пола. В затылочном и височном кластерах электродов амплитуда Р200 была значимо выше в группе мужчин по сравнению с женщинами. Что касается функциональной роли компонента Р200, то многие исследователи связывают увеличение Р200 с реакцией на нецелевые стимулы [29–31], в связи с чем компонент Р200 интерпретируется как отражение процесса подавления нерелевантной информации [32, 33]. Так, например, в работе [32] показано, что при выполнении задачи поиска целевого стимула (полоска определенной пространственной ориентации и цвета) в присутствии в зрительном поле дистракторов (полоски других ориентаций и цветов) амплитуда Р200 зависит от числа характеристик, которые отличают дистрактор от значимого стимула. Именно за счет избирательного внимания к этим характеристикам осуществляется подавление незначимой информации и выбор кандидатов для последующего сопоставления с целевым стимулом. Важность событий в затылочной коре в интервале Р200 для имплицитного обнаружения различий между стандартным и измененным иероглифом описана в работе [34]. Данные ряда исследований указывают на значимость Р200 как показателя сопоставления информации от сенсорных входов с сохраненной в памяти информацией [29, 35].
В ряде работ компонент Р200 рассматривается как возможный нейрофизиологический маркер такого свойства РП, которое в англоязычной литературе обозначается как “capacity”, что может соответствовать терминам “способность”, “вместимость”. Так, в работе [29] характеристики Р200 ССП теменных областей коры зависели не только от совпадения/несовпадения текущих и ранее прослушанных цифр, но и от количества удерживаемых в памяти знаков. Аналогичные результаты представлены и в других работах [32, 36].
Компонент Р200 связывают также со способностью эффективно управлять вниманием. Это убедительно показано в работе [37], в которой при выполнении задачи разделения значимых и незначимых ориентационных решеток испытуемые, играющие в видео-игры, демонстрировали бо̀льшую амплитуду компонента Р200 в центральной области коры по сравнению с неигроками, что авторы связывают с высоким уровнем контроля внимания и восприятия как результатом тренировок. Корреляции Р200 и внимания обсуждают и другие авторы. Например, в задаче обнаружения различий двух последовательных стимулов увеличение объема вводимой информации приводило к росту амплитуды Р200 в затылочных и височных областях, но без снижения производительности РП [36]. Предполагается, что отсутствие поведенческого ухудшения может быть результатом регулируемого сверху [35, 38] привлечения дополнительных ресурсов внимания для поддержания эффективности выполнения задачи РП, что и отражается в увеличении амплитуды Р200. Сходный эффект описан в работе [39].
Таким образом, полученный в нашей работе результат о бо̀льшей амплитуде Р200 в ССП каудальных областей коры у мужчин можно рассматривать как показатель более высокого уровня селективного внимания к ориентационным свойствам зрительной среды. Кроме того, бо̀льший по амплитуде компонент Р200 в каудальных областях коры у мужчин по сравнению с женщинами может быть проявлением гендерно-обусловленной специфики селективного внимания [40].
Повышение амплитуды у мужчин по сравнению с женщинами обнаружено также для медленной позитивной волны, развивающейся в центральных отведениях; различия значимы в интервале от 400 до 500 мс. Позитивное отклонение в центрально-теменных областях с пиковой латентностью около 300 мс и позже – один из наиболее частых феноменов, регистрируемых в широком спектре когнитивных задач и обозначаемый традиционно как Р300 [41]. Амплитуда Р300 вариабельна и может быть максимальной в диапазоне от 200–250 до 800 мс. Согласно литературе, семейство Р300 включает в себя три накладывающихся друг на друга позитивных отклонения: Р3а с пиком около 250 мс, Р3b с пиком около 350 мс и позитивная медленная волна. Все они в равной степени зависят от вероятности появления стимула, но по-разному чувствительны к характеристикам релевантного стимула и имеют разную связь со вниманием [41]. Одним из важных факторов, влияющим на латентность пика и длительность волны считается трудность задания [41–43], которая модулируется, например, числом стимулов или набором значимых характеристик релевантного сигнала.
В литературе представлены немногочисленные и противоречивые сведения о половых различиях Р300 [44–47]. Причины неоднозначности данных о половых различиях Р300 подробно обсуждаются в работе [44].
Что касается зрительной РП, то Р300 считается важным индикатором кодирования и извлечения информации из памяти. В этом отношении показательна работа [42], в которой авторы, исследуя хронометрию извлечения информации из памяти, обнаружили, что при более сложном задании отставленного сопоставления в интервале компонента P3b можно выделить два временны́х промежутка. При этом поздний подкомпонент в бо̀льшей степени связан с операциями поиска информации в памяти, особенно в сложных задачах, которые не могут быть решены на основании обнаружения простого сходства. В исследованиях с категоризацией объектов, авторы обращают внимание на позднее (после 400 мс) позитивное отклонение в центрально-теменных отведениях. Его связывают с операцией сравнения объекта с другими представителями категории, хранящимися в памяти, и считают индексом доступа к хранилищу [48, 49].
Для понимания нашего результата о более высокой амплитуде позитивности 400–500 мс в группе мужчин по сравнению с женщинами также важны сведения о положительной связи Р300 с оценками “общих когнитивных способностей” по Векслеру [50], а также с индивидуальными оценками “объема памяти”, как общего когнитивного ресурса, не зависящего от материала [51]. Люди с бо̀льшей емкостью РП, как правило, демонстрируют большую P300 при кодировании информации. Исходя из изложенного выше, можно предположить, что известное для мужского пола преимущество выполнения зрительно-пространственных задач, в том числе задач РП, может проявляться как усиление позитивности в диапазоне Р300, не обязательно связанное с непосредственной деятельностью, а отражающее “способность” к выполнению этого класса задач. В основе таких более широких возможностей РП у мужчин может лежать гендерно-зависимая специфичность нейросетей РП [52–54].
Анализ поведенческих показателей (ВР и точность) не обнаружил значимых различий между мужчинами и женщинами. В обеих группах ВР при несовпадении эталонной и тестовой ориентаций выше, чем при их совпадении, что соответствует литературным данным [55, 56].
Интересным и важным результатом представляется связь ВР выполнения задачи сличения ориентаций с оценками стратегий навигационного поведения. Более высокие оценки по шкале стратегии “ориентирования”, опирающейся на учет метрических характеристик и глобальных пространственных ориентиров, соответствуют более быстрому выполнению задания. Напротив, более высокие оценки по шкале стратегии “маршрута”, учитывающей топографию ближних меток на местности, соответствуют более медленному выполнению задачи. Важно, что эта зависимость четко проявляется только в группе мужчин.
Для обсуждения полученных данных интересны сведения, приведенные в обзоре [57], о биологических и социально-культурных факторах половых различий ориентационного поведения, а также о связи этих различий с разными стратегиями решения навигационных задач. Известно, что мужчины полагаются на глобальные контрольные точки и конфигурационные средовые характеристики, тогда как женщины используют локальные ориентиры и “стратегию маршрута”, соблюдая инструкции, как добраться от места до места [58]. Некоторые авторы считают, что различия между мужчинами и женщинами не связаны с лучшей ориентацией у мужчин, а обусловлены разными стратегиями [59]. В исследовании [59] участников попросили пройти по четырем неизвестным направлениям. Местоположение каждого пункта назначения определялось, следуя инструкциям. В инструкции “на основе Евклидовых характеристик” необходимо указать направления (например, север или запад) и метрические расстояния (например, 100 м). В “ориентировочной” инструкции просили указать заметный ориентир (например, фиолетовые двери) и эгоцентрические (вправо или влево) направления поворота. Обнаружено, что мужчины лучше всего справляются с навигационными задачами, когда их предоставляют в евклидовых координатах, тогда как женщины справляются лучше, когда им предоставляется информация об ориентирах. По данным исследования [15], в ситуации, когда доступны лишь локальные ориентиры, половые различия не обнаруживаются. Мужчины, предпочитающие конфигурационные стратегии, при необходимости используют и локальные ориентиры, т.е., они могут менять стратегии, адаптируясь к доступной информации из окружающей среды. Женщины, предпочитающие использовать локальные ориентиры, испытывают больше трудностей при перестройке стратегий.
Полученный нами результат о связи психофизических характеристик выполнения задачи сличения ориентаций в модели РП с оценками стратегий навигации, находится в соответствии с представлениями, что поиск пути является специфической нагрузкой на зрительно-пространственную РП [60]. Поэтому люди с высокими баллами в тесте Корси, оценивающим краткосрочную зрительно-пространственную память, имеют оптимальную память на описание маршрута [61]. Более того, авторы исследования [62], используя регрессионный подход, обнаружили, что результативность зрительно-пространственной РП прогнозирует успешность навигационного поведения.
ЗАКЛЮЧЕНИЕ
В группе 38 испытуемых (19 женщин) в модели РП обнаружены половые различия нейронных механизмов сопоставления ориентационных характеристик зрительного пространства. Анализ ССП показал, что у мужчин амплитуда ранней негативности в затылочной и височной областях коры чувствительна к фактору совпадение/несовпадение текущей и удерживаемой в памяти ориентаций, что говорит об эффективном раннем детектировании изменений ориентационных характеристик в этой группе. Повышение амплитуды компонента Р200 в затылочно-височной области у мужчин по сравнению с женщинами в контексте литературных данных [36] свидетельствует о более высоком уровне селективного внимания к основным пространственным ориентирам. Повышение амплитуды центральной позитивности 400–500 мс у мужчин по сравнению с женщинами указывает на потенциальную возможность удержания в РП бо̀льшего объема информации об ориентационных свойствах. В работе выявлена статистически значимая связь ВР выполнения задачи РП на ориентации и оценок стратегий навигационного поведения по данным анкетирования. Более высокие оценки по шкале стратегии “ориентирования”, опирающейся на учет метрических характеристик и глобальных пространственных ориентиров, соответствуют более быстрому выполнению задания сличения ориентаций. Напротив, более высокие оценки по шкале стратегии “маршрута”, учитывающей топографию ближних меток на местности, соответствуют более медленному выполнению задания. При раздельном анализе в группах мужчин и женщин, вышеописанные зависимости проявлялись только у мужчин. Таким образом, в настоящей работе экспериментально подтверждено, что одним из важных биологических факторов, лежащих в основе более успешной стратегии навигационного поведения, учитывающей глобальные пространственные характеристики, является эффективная зрительно-пространственная РП на базовые характеристики зрительной среды.
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены локальным биоэтическим комитетом Института высшей нервной деятельности и нейрофизиологии РАН (Москва). Протокол № 1 от 15 января 2020 г.
Информированное согласие. Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Финансирование работы. Работа поддержана средствами государственного бюджета по государственному заданию Министерства образования и науки РФ на 2021–2023 гг. Электрофизиологические исследования выполнены на базе Центра коллективного пользования научным оборудованием для функционального картирования мозга на базе ИВНД и НФ РАН (Москва).
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией данной статьи.
Вклад авторов в публикацию. Е.С. Михайлова и Н.Ю. Мошникова планировали исследование, программировали экспериментальные серии в программе E-Prime. Н.Ю. Мошникова и А.Б. Кушнир создавали библиотеку изображений, провели эксперименты, делали иллюстрации. Е.С. Михайлова, Н.Ю. Мошникова и А.Б. Кушнир обработали и проанализировали полученные данные. Е.С. Михайлова написала статью. Все авторы внесли свой вклад и одобрили окончательный вариант рукописи.
Список литературы
Cahill L. Why sex matters for neuroscience // Nat. Rev. Neurosci. 2006. V. 7. № 6. P. 477.
Voyer D., Voyer S.D., Saint-Aubin J. Sex differences in visual-spatial working memory: A meta-analysis // Psychon. Bull. Rev. 2017. V. 24. № 2. P. 307.
Shaqiri A., Roinishvili M., Grzeczkowski L. et al. Sex-related differences in vision are heterogeneous // Sci. Rep. 2018. V. 8. № 1. P. 7521.
Voyer D., Voyer S., Bryden M.P. Magnitude of sex differences in spatial abilities: A meta-analysis and consideration of critical variables // Psychol. Bull. 1995. V. 117. № 2. P. 250.
Galea L.A., Kimura D. Sex differences in route-learning // Pers. Individ. Dif. 1993. V. 14. № 1. P. 53.
Moffat S.D., Hampson E., Hatzipantelis M. Navigation in a virtual maze: sex differences and correlation with psychometric measures of spatial ability in humans // Evol. Hum. Behav. 1998. V. 19. P. 73.
Christie G.J., Cook C.M., Ward B.J. et al. Mental rotational ability is correlated with spatial but not verbal working memory performance and P300 amplitude in males // PLoS One. 2013. V. 8. № 2. P. e57390.
Крылова М.А., Изъюров И.В., Герасименко Н.Ю. и др. Моделирование источников компонентов зрительных вызванных потенциалов человека в задаче определения ориентации отрезков линий // Журн. высш. нерв. деят. им. И.П. Павлова. 2015. Т. 65. № 6. С. 685. Krylova M.A., Izyurov I.V., Gerasimenko N.Yu. et al. [The Modeling of Human Visual ERPs Sources in the Task of Line Orientation Identification] // Zh. Vyssh. Nerv. Deiat. im. I.P. Pavlova. 2015. V. 65. № 6. P. 685.
Михайлова Е.С., Герасименко Н.Ю., Крылова М.А. и др. Механизмы ориентационной чувствительности зрительной системы человека. Сообщение II. Корковые механизмы ранних этапов переработки информации об ориентации линий // Физиология человека. 2015. Т. 41. № 3. С. 5. Mikhailova E.S., Gerasimenko N.Yu., Krylova M.A. et al. Mechanisms of orientation sensitivity of human visual system: Part II. Neural patterns of early processing of information about line orientation // Human Physiology. 2015. V. 41. № 3. P. 229.
Михайлова Е.С., Герасименко Н.Ю., Кушнир А.Б. Психофизические и нейрофизиологические характеристики оценки наклонных ориентаций у мужчин и женщин // Сенсорные системы. 2020. Т. 34. № 4. С. 283. Mikhailova E.S., Gerasimenko N.Yu., Kushnir A.B. Psychophysical and neurophysiological characteristics of the assessment of tilted orientations in men and women // Neurosci. Behav. Physiol. 2021. V. 51. № 6. P. 820.
Barkley C.L., Jacobs L.F. Sex and species differences in spatial memory in food-storing kangaroo rats // Anim. Behav. 2007. V. 73. № 2. P. 321.
Gaulin S.J.C., Fitzgerald R.W. Sex-differences in spatial ability – an evolutionary hypothesis and test // Am. Nat. 1986. V. 127. P. 74.
Langley C.M. Spatial memory in the desert kangaroo rat (Dipodomys deserti) // J. Comp. Psychol. 1994. V. 108. № 1. P. 3.
Williams C.L., Barnett A.M., Meck W.H. Organizational effects of early gonadal secretions on sexual differentiation in spatial memory // Behav. Neurosci. 1990. V. 104. № 1. P. 84.
Sandstrom N.J., Kaufman J., Huettel S.A. Males and females use different distal cues in a virtual environment navigation task // Brain Res. Cogn. Brain Res. 1998. V. 6. № 4. P. 351.
Jones C.M., Healy S.D. Differences in cue use and spatial memory in men and women // Proc. R. Soc. B Biol. Sci. 2006. V. 273. № 1598. P. 2241.
Kelly D.M., Bischof W.F. Reorienting in images of a three-dimensional environment // J. Exp. Psychol. Hum. Percept. Perform. 2005. V. 31. № 6. P. 1391.
Jacobs L.F., Schenk F. Unpacking the cognitive map: the parallel map theory of hippocampal function // Psychol. Rev. 2003. V. 110. № 2. P. 285.
Kemp A., Manahan-Vaughan D. The hippocampal CA1 region and dentate gyrus differentiate between environmental and spatial feature encoding through long-term depression // Cereb. Cortex. 2008. V. 18. № 4. P. 968.
Chai X.J., Jacobs L.F. Sex differences in directional cue use in a virtual landscape // Behav. Neurosci. 2009. V. 123. № 2. P. 276.
Lejbak L., Crossley M., Vrbancic M. A male advantage for spatial and object but not verbal working memory using the n-back task // Brain Cogn. 2011. V. 76. № 1. P. 191.
Lawton C.A., Kallai J. Gender differences in wayfinding strategies and anxiety about wayfinding: A cross-cultural comparison // Sex Roles. 2002. V. 47. № 9–10. P. 389.
Chen C.-C., Kuo J.-C., Wang W.-J. Distinguishing the visual working memory training and practice effects by the effective connectivity during n-back tasks: A DCM of ERP study // Front. Behav. Neurosci. 2019. V. 13. P. 84.
Pasternak T., Greenlee M.W. Working memory in primate sensory systems // Nat. Rev. Neurosci. 2005. V. 6. № 2. P. 97.
Postle B.R. Working memory as an emergent property of the mind and brain // Neuroscience. 2006. V. 139. № 1. P. 23.
Agam Y., Sekuler R. Interactions between working memory and visual perception: an ERP/EEG study // Neuroimage. 2007. V. 36. № 3. P. 933.
Михайлова Е.С., Герасименко Н.Ю., Славуцкая А.В. Сенсорные механизмы ранней дискриминации ориентаций в модели зрительной рабочей памяти // Журн. высш. нерв. деят. им. И.П. Павлова. 2019. Т. 69. № 5. С. 577. Mikhailova E.S., Gerasimenko N.Yu., Slavutskaya A.V. Sensory mechanisms in early orientation discrimination in a model of visual working memory // Neurosci. Behav. Physiol. 2020. V. 50. № 6. P. 700.
Gur R.C., Alsop D., Glahn D. et al. An fMRI study of sex differences in regional activation to a verbal and a spatial task // Brain Lang. 2000. V. 74. № 2. P. 157.
Lefebvre C.D., Marchand Y., Eskes G.A. et al. Assessment of working memory abilities using an event-related brain potential (ERP)-compatible digit span backward task // Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2005. V. 116. № 7. P. 1665.
Wang A.L., Mouraux A., Liang M. et al. The enhancement of the N1 wave elicited by sensory stimuli presented at very short inter-stimulus intervals is a general feature across sensory systems // PLoS One. 2008. V. 3. № 12. P. e3929.
Lenartowicz A., Escobedo-Quiroz R., Cohen J.D. Updating of context in working memory: an event-related potential study // Cogn. Affect. Behav. Neurosci. 2010. V. 10. № 2. P. 298.
Phillips S., Takeda Y. An EEG/ERP study of efficient versus inefficient visual search / Proceedings of the Annual Meeting of the Cognitive Science Society. 2009. P. 383.
Coenen A. Modelling of auditory evoked potentials of human sleep-wake states // Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2012. V. 85. № 1. P. 37.
Zhang X., Yang S., Jiang M. Rapid implicit extraction of abstract orthographic patterns of Chinese characters during reading // PLoS One. 2020. V. 15. № 2. P. e0229590.
Freunberger R., Klimesch W., Doppelmayr M. et al. Visual P2 component is related to theta phase-locking // Neurosci. Lett. 2007. V. 426. № 3. P. 181.
Cepeda-Freyre H.A., Garcia-Aguilar G., Eguibar J.R. et al. Brain processing of complex geometric forms in a visual memory task increases P2 Amplitude // Brain Sci. 2020. V. 10. № 2. P. 114.
Föcker J., Mortazavi M., Khoe W. et al. Neural correlates of enhanced visual attentional control in action video game players: An event-related potential study // J. Cogn. Neurosci. 2019. V. 31. № 3. P. 377.
Linnert S., Reid V., Westermann G. ERP correlates of two separate top-down mechanisms in visual categorization // Int. J. Psychophysiol. 2016. V. 108. P. 83.
Vogel E.K., Machizawa M.G. Neural activity predicts individual differences in visual working memory capacity // Nature. 2004. V. 428. № 6984. P. 748.
Bianco V., Berchicci M., Quinzi F. et al. Females are more proactive, males are more reactive: neural basis of the gender-related speed/accuracy trade-off in visuo-motor tasks // Brain Struct. Funct. 2020. V. 225. № 1. P. 187.
Picton T.W. The P300 wave of the human event-related potential // J. Clin. Neurophysiol. Off. Publ. Am. Electroencephalogr. Soc. 1992. V. 9. № 4. P. 456.
Bledowski C., Cohen Kadosh K., Wibral M. et al. Mental chronometry of working memory retrieval: a combined functional magnetic resonance imaging and event-related potentials approach // J. Neurosci. Off. J. Soc. Neurosci. 2006. V. 26. № 3. P. 821.
Rawdon C., Murphy J., Blanchard M.M. et al. Reduced P300 amplitude during retrieval on a spatial working memory task in a community sample of adolescents who report psychotic symptoms // BMC Psychiatry. 2013. V. 13. № 1. P. 125.
Maciejewska K., Drzazga Z. Differences in spatio‑temporal distribution of the visual P3b event-related potential between young men and women // Acta Neurobiol. Exp. (Wars). 2019. V. 79. P. 25.
Vaquero E., Cardoso M.J., Vázquez M. et al. Gender differences in event-related potentials during visual-spatial attention // Int. J. Neurosci. 2004. V. 114. № 4. P. 541.
Deldin P.J., Duncan C.C., Miller G.A. Season, gender, and P300 // Biol. Psychol. 1994. V. 39. № 1. P. 15.
Steffensen S.C., Ohran A.J., Shipp D.N. et al. Gender-selective effects of the P300 and N400 components of the visual evoked potential // Vision Res. 2008. V. 48. № 7. P. 917.
Morrison R.G., Reber P.J., Bharani KL. et al. Dissociation of category-learning systems via brain potentials // Front. Hum. Neurosci. 2015. V. 9. P. 389.
Rabi R., Joanisse M.F., Zhu T. et al. Cognitive changes in conjunctive rule-based category learning: An ERP approach // Cogn. Affect. Behav. Neurosci. 2018. V. 18. № 5. P. 1034.
Gevins A., Smith M.E. Neurophysiological measures of working memory and individual differences in cognitive ability and cognitive style // Cereb. Cortex. 2000. V. 10. № 9. P. 829.
Nittono H., Nageishi Y., Nakajima Y. et al. Event-related potential correlates of individual differences in working memory capacity // Psychophysiology. 1999. V. 36. № 6. P. 745.
Hill A.C., Laird A.R., Robinson J.L. Gender differences in working memory networks: a BrainMap meta-analysis // Biol. Psychol. 2014. V. 102. P. 18.
Alarcón G., Cservenka A., Fair D.A. et al. Sex differences in the neural substrates of spatial working memory during adolescence are not mediated by endogenous testosterone // Brain Res. 2014. V. 1593. P. 40.
Zilles D., Lewandowski M., Vieker H. et al. Gender differences in verbal and visuospatial working memory performance and networks // Neuropsychobiology. 2016. V. 73. № 1. P. 52.
Yin J., Gao Z., Jin X. et al. Tracking the mismatch information in visual short term memory: an event-related potential study // Neurosci. Lett. 2011. V. 491. № 1. P. 26.
Михайлова Е.С., Герасименко Н.Ю., Славуцкая А.В. и др. Временны́е и топографические характеристики вызванных потенциалов в ситуации конфликта двух последовательных зрительных стимулов в задаче на зрительную память // Физиология человека. 2017. Т. 43. № 3. С. 13. Mikhailova E.S., Gerasimenko N.Yu., Slavutskaya A.V. et al. Temporal and topographic characteristics of evoked potentials in the conflict of two consecutive visual stimuli in a working memory task // Human Physiology. 2017. V. 43. № 3. P. 248.
Coluccia E., Louse G. Gender differences in spatial orientation: A review // J. Environ. Psychol. 2004. V. 24. № 3. P. 329.
Lawton C.A. Strategies for indoor way-finding:the role of orientation // J. Environ. Psychol. 1996. V. 16. P. 137.
Saucier D.M., Green S.M., Leason J. et al. Are sex differences in navigation caused by sexually dimorphic strategies or by differences in the ability to use the strategies? // Behav. Neurosci. 2002. V. 116. № 3. P. 403.
Garden S., Cornoldi C., Logie R.H. Visuo-spatial working memory in navigation // Appl. Cogn. Psychol. 2002. V. 16. № 1. P. 35.
Pazzaglia F., Cornoldi C. The role of distinct components of visual-spatial working memory in the processing of texts // Memory. 1999. V. 7. № 1. P. 19.
Bosco A., Longoni A.M., Vecchi T. Gender effects in spatial orientation: Cognitive profiles and mental strategies // Appl. Cogn. Psychol. 2004. V. 18. № 5. P. 519.
Дополнительные материалы отсутствуют.
Инструменты
Физиология человека