

Доклады Российской академии наук. Химия, науки о материалах , 2020, T. 495, № 1, стр. 83-87
Компьютерное моделирование химико-микробиологических процессов в ризомикробиофитосферной системе
Академик РАН В. П. Мешалкин 1, 2, В. А. Четырбоцкий 1, *, А. Н. Четырбоцкий 3, А. Ф. Пэлий 4
1 Российский химико-технологический университет
им. Д.И. Менделеева
Москва, Россия
2 Институт общей и неорганической химии
им. Н.С. Курнакова Российской академии наук
Москва, Россия
3 Дальневосточный геологический институт Дальневосточного отделения Российской академии наук
Владивосток, Россия
4 Российский университет дружбы народов
Москва, Россия
* E-mail: vchetyrbotskii@gmail.com
Поступила в редакцию 04.07.2020
После доработки 10.11.2020
Принята к публикации 12.11.2020
Аннотация
Разработана компьютерная модель химико-микробиологических продукционных процессов биомассы растений, почвенных микроорганизмов и элементов питания в ризосфере – сложной гетерогенной полидисперсной многофакторной природной системы, – что позволяет определять рост биомассы сельскохозяйственных культур и продуктивность ризосферы. Проведены параметрическая идентификация и проверка адекватности разработанной математической модели на основании имеющейся выборки экспериментальных данных роста яровой пшеницы сорта “Красноуфимская-100”. Проведены вычислительные эксперименты по определению безопасного срока эксплуатации земель сельскохозяйственного назначения при внесении удобрений с различной степенью содержания тяжелых металлов.
Для обеспечения высокого плодородия почвы, оптимизации цикла выращивания урожая культур сельскохозяйственного назначения, а также прогноза их состояния актуальна задача построения модели совместной динамики органических и неорганических составляющих почвы, учитывающей сложные химико-микробиологические связи между элементами ризосферы. Ее многофакторность обусловливает рассмотрение изменений достаточно большого числа параметров, а функциональные отношения обусловлены нелинейными взаимодействиями, трудно поддающимися традиционному формализованному представлению.
Исследование биологических продукционных процессов в ассоциациях живых организмов представлено в [1]. Динамика отдельной биотической фазы, в которой приводятся выраженные в количественных формах различные аспекты жизненного цикла растений: рост, дыхание, фотосинтез и распределение продуктов жизнедеятельности, – детально рассмотрена в [2]. Однако за рамками рассматриваемых моделей оказываются проблемы согласования совместной динамики биомассы растений и распределенных в ризосфере (прилегающей к корням растений узкой зоны почвы) элементов минерального питания, динамики популяции почвенных микроорганизмов, которые являются катализаторами круговорота питательных веществ в ризосфере. Открытыми остаются вопросы выбора и проведения процедур параметрической идентификации и оценки адекватности сложных многопараметрических моделей ризосферы [3, 4]. Имеющаяся экспериментальная база таких моделей представлена достаточно большим массивом данных, анализ которой в рамках исследуемых систем представляет собой сложную задачу [5].
В связи с этим нами разработана компьютерная модель химико-микробиологических процессов в ризомикробиофитосферной системе как совокупности взаимосвязанных внутрипочвенными потоками внешних минеральных питательных веществ, множества корней сельскохозяйственных растений и колоний почвенных микроорганизмов, отличающаяся использованием трех типовых уравнений Ферхюльста, отображающих во времени: временное накопление питательных элементов в корнях растений, химико-микробиологический процесс роста биомассы корней растений в зависимости от численности популяции микроорганизмов с учетом убыли массы корней как жертвы под воздействием поедающих микроорганизмов-хищников, зависимость химико-микробиологического процесса роста популяции микроорганизмов от количества питательных элементов с учетом роста численности популяции микроорганизмов и убывающего количества нутриентов, – что позволяет определять рост биомассы сельскохозяйственных растений и продуктивность ризосферы.
Блок-схема модели химико-микробиологических процессов в ризомикробиофитосферной системе представлена на рис. 1.
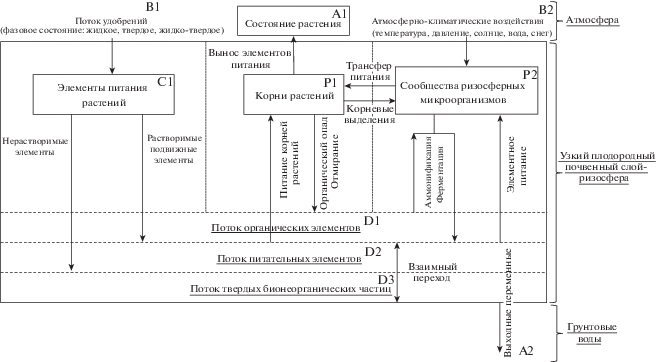
Функциональность представленной схемы состоит в следующем. Элементами системы являются ризосфера (узкий плодородный почвенный слой), корни растений (P1) и сообщества ризосферных микроорганизмов (P2). Эти элементы взаимодействуют посредством непрерывных потоков вещества внутри системы. Так, например, корни растений выделяют в почву потоки белков, аминокислот, глюкозу, которые поглощаются микроорганизмами и растениями, тем самым увеличивая численность своей популяции и биомассу соответственно. В свою очередь, микроорганизмы повышают плодородие почвы, сохраняют баланс питательных веществ в среде, взаимодействуют с корнями растений, восполняя дефицит необходимых для растений элементов, а также контролируют обмен веществ в среде.
Входными переменными системы являются вносимые удобрения (B1), принимающие различные состояния (жидкое, твердое, жидко-твердое), атмосферно-климатические воздействия (B2), которые в основном характеризуются температурой, давлением, воздействием со стороны солнца, воды, снега и т.д.
Поступающие в ризосферу удобрения (C1) расщепляются на питательные элементы (D2) и твердые бионеорганические частицы (D3). При этом отдельные элементы системы сами продуцируют органические вещества с помощью почвенной микробиоты (отмершие ризосферные микроорганизмы, органический опад листьев растений, корни и вещества жизнедеятельности всей почвенной органики). Связь между ними (D1, D2, D3) выполняется за счет почвенного водного раствора ризосферы, физического состояния самой почвы, а также функций жизнедеятельности растений и микроорганизмов.
Выходными переменными выступают состояние растения (A1) и суммарный поток питательных элементов из ризосферы (P1), доставляемый к корням этого растения.
Приведенная на рис. 1 блок-схема позволяет составить балансовые уравнения модели основных химико-микробиологических процессов в ризосфере.
Динамика потребляемых растением элементов питания ризосферы следует принятым в модели “ресурс–потребитель” положениям [6–8], где ресурсом питания растений выступают поступившие в качестве удобрений химические элементы, а потребителями – растения. Допускаются только парные взаимодействия между элементами системы, которые следуют уравнению баланса вещества [9]. Динамика роста растений следует фактору самолимитирования, который подробно описан в модели Ферхюльста [6]. Тогда согласно принятым допущениям динамика элементов питания в растении определяется записью:
(1)
$\frac{{d{{C}_{i}}}}{{dt}} = \left( {{{p}_{{i,0}}}{{C}_{{i,0}}} - \mathop \sum \limits_{j = 1}^3 {{p}_{{i,j}}}{{C}_{j}}} \right)\left( {qY + pB} \right),$Для ассоциации микроорганизмов ризосферы присутствующие в ней продукты органического происхождения (отмершие органы и ткани растений и животных), сами растения и минеральные элементы ризосферы представляют основной единый ресурс их питания, что обусловливает конкурентные отношения за общий для микроорганизмов ресурс их питания:
(2)
$\frac{{dY}}{{dt}} = \left[ {{{d}_{0}}\, + \,\mathop \sum \limits_{k = 1}^3 \left( {{{d}_{{k,0}}}{{C}_{{k,0}}} - \mathop \sum \limits_{j = 1}^3 {\kern 1pt} {{d}_{{k,j}}}{{C}_{k}}} \right)\, + \,{{d}_{B}}B\, - \,{{d}_{Y}}Y} \right]Y,$Удельный рост биомассы растений определяется их естественным приростом, внутри- и межвидовыми отношениями растений, содержанием в ризосфере элементов питания подвижной формы и отношениями “микроорганизмы–растение”:
(3)
$\frac{{dB}}{{dt}} = \left[ {{{b}_{0}} + \mathop \sum \limits_{k\, = \,1}^3 \left( {{{b}_{{k,0}}}{{C}_{{k,0}}}\, - \,\mathop \sum \limits_{j\, = \,1}^3 {\kern 1pt} {{b}_{{k,j}}}{{C}_{k}}} \right)\, - \,{{b}_{Y}}Y - {{b}_{B}}B} \right]B,$Система дифференциальных уравнений в частных производных уравнений (1)–(3) замыкается следующими начальными и граничными условиями:
(4)
$\begin{gathered} {{C}_{i}}\left( {{{t}_{0}}} \right) = 0,\quad Y\left( {{{t}_{0}}} \right) = {{Y}_{0}},\quad B\left( {{{t}_{0}}} \right) = {{B}_{0}}, \\ {{\left. {\frac{{\partial {{Y}_{k}}}}{{\partial{ \vec {n}}}}} \right|}_{{\Gamma }}} = 0,\quad {{\left. {\frac{{\partial {{B}_{m}}}}{{\partial{ \vec {n}}}}} \right|}_{{\Gamma }}} = 0, \\ \end{gathered} $Численное решение задачи Коши системы уравнений (1)–(4) при начальном и граничном условиях уравнения (4) следует двухшаговому методу 2-го порядка точности “с перешагиванием” решения обыкновенных дифференциальных уравнений [10]:
Цифровым материалом для исследований выступают результаты выполненных экспериментов по оценке влияния азотных, фосфорных и калийных удобрений на рост яровой пшеницы сорта “Красноуфимская-100” на торфяной низинной почве [11, 12].
Оценка коэффициентов модели следует результату поиска минимума нормы невязки выборочных и модельных распределений, в котором условием прекращения процедуры является превышение наперед заданного значения так называемой функции толерантности. Для оценивания параметров была выполнена 231 итерация поиска минимума заданной нормы. На первой итерации значение нормы оказалось равным 273, а на последней – 163 (уменьшение в 1.7 раза).
Результаты оценки коэффициентов указывают на следующее: присутствие фосфора в почвенном питательном растворе ведет к ингибированию сорбции калия и азота в корневые поры растений, тогда как наличие азота способствует сорбции калия. Оценка численных значений коэффициентов также позволяет определить степень их влияния на динамику роста биомассы рассматриваемой культуры, где азот имеет повышенную значимость над другими, что свидетельствует о его ведущей роли в процессе формирования биомассы. Далее следует фосфор, а затем калий.
Оценка адекватности модели выполнялась на основании коэффициента корреляции, который составил 0.9. Такое высокое значение указывает на хорошее соответствие модели изучаемому процессу. На рис. 2 изображены выборочные и модельные распределения выноса Cd надземной частью биомассы растений.
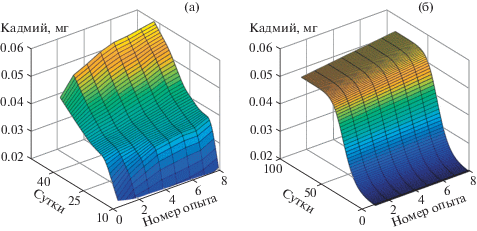
На основании модели (1)–(4) была разработана методика оценивания загрязнения почв и растений тяжелыми металлами (ТМ) при внесении в почву минеральных удобрений (МУ), поскольку фосфатные и калийные руды являются основными составляющими МУ, которые аккумулируют ТМ в процессе своего формирования. В частности, кадмия, одного из высокотоксичных ТМ [13–15]. Суть методики состоит в следующем: на основании уравнений (1)–(4) был имитирован многолетний жизненный цикл растения (вегетационный период), где каждый год наряду с вносимыми в ризосферу удобрениями, содержащими кадмий, оценивалось его остаточное содержание в растениях и почве. Условия проведения эксперимента учитывают ту долю ТМ, которая поглотилась надземной частью растения и которая в конце каждого вегетационного периода удаляется (например, снятие урожая). Далее на основании вычисленных распределений оценивался период времени, в течение которого содержание Cd превысит предельно допустимую концентрацию (ПДК) согласно законодательным актам Российской Федерации.
Проблема очистки почв с помощью сельскохозяйственных растений является основной задачей “зеленой” биотехнологии по разработке, созданию и внедрению таких культур в агросистемы, принятой Международным Конгрессом по биотехнологии [16]. Результаты экспериментов показывают, что сельскохозяйственные почвы нельзя будет эксплуатировать при использовании слабозагрязненных (20 мг Cd/кг P2O5) удобрений на протяжении 55 лет внесения, среднезагрязненных (40 мг Cd/кг P2O5) – 89 лет, сильнозагрязненных (60 мг Cd/кг P2O5) – 271 год.
В результате проведенных исследований разработана компьютерная модель химико-микробиологических процессов в ризомикробиофитосферной системе, сущность которой состоит в стремлении динамических переменных модели к единому механизму потребления элементов питания. В ней также присутствует ограниченный рост составляющих элементов системы, который определяется такими важными факторами биологической среды, как внутри- и межвидовая конкуренция, ограниченная жизнедеятельность биологических видов, предельный запас питания составляющих почвенной среды. Разработан механизм и проведена оценка на территориях Российской Федерации загрязнения почв ТМ при внесении удобрений. Данный механизм составляет сущность предмета “зеленой” биотехнологии.
Список литературы
Ризниченко Г.Ю., Рубин А.Б. Математические модели биологических продукционных процессов / М.: Изд-во МГУ, 1983. 302 с.
Торнли Дж.Г.М. Математические модели в физиологии растений (пер. с англ.) / Киев.: Наук. думка, 1982. 312 с.
Мешалкин В.П., Ивашкин Ю.А., Никитина М.А. Компьютерная мультиагентная модель химико-физиологических процессов в желудочно-кишечном тракте человека как в живой биохимической системе // ДАН. 2019. Т. 484. № 3. С. 303–306. https://doi.org/10.31857/S0869-56524843303-306
Кафаров В.В., Мешалкин В.П. Анализ и синтез химико-технологических систем: Учебник для вузов / М.: Химия, 1991. 432 с.
Мешалкин В.П., Чионов А.М., Казак А.С., Аристов В.М. Компьютерная модель нестационарного газового потока в протяженном многослойно изолированном подводном газопроводе высокого давления // ДАН. 2016. Т. 469. № 6. С. 694–697. https://doi.org/10.7868/s0869565216240129
Свирежев Ю.М. Нелинейные волны, диссипативные структуры и катастрофы в экологии / М.: Наука, 1987. 368 с.
Murray J.D. Mathematical Biology / B.: Springer, 2007. 574 p. https://doi.org/10.1007/978-3-662-08542-4
Schmidt B. The art of modeling and simulation / SCS-Europe BVBA. Chent: Belgium, 2001. 480 p.
Кафаров В.В., Мешалкин В.П. Анализ и синтез химико-технологических систем / М.: Химия, 1991. 432 с.
Поттер Д.М. Вычислительные методы в физике (пер. с англ.) / М: Мир, 1975. 392 с.
Ефремова М.А., Дричко В.Ф. Влияние калия на физико-химические свойства торфяной низинной почвы //Агрохимия. 2010. Т. 43. № 4. С. 3–10.
Ефремова М.А., Сладкова Н.А., Вяльшина А.С. Динамика накопления кадмия и калия растениями пшеницы на дерново-подзолистой и торфяной низинной почвах //Агрохимия. 2013. № 11. С. 86–96.
Violante A., Cozzolino V., Peremolov L., Caporale A.G., Pigna M. Mobility and bioavailability of heavy metals and metalloids in soil environments // J. Soil. Sci. Plant Nutr. 2010. V. 10. № 3. P. 268–292. https://doi.org/10.4067/S0718-95162010000100005
Водяницкий Ю.Н. Загрязнение почв тяжелыми металлами и металлоидами / М.: МГУ им. М.В. Ломоносова, 2017. 192 с.
Казнина Н.М., Титов А.Ф., Лайдинен Г.Ф., Батова Ю.В. Влияние кадмия на некоторые физиологические показатели растений ячменя в зависимости от их возраста // Труды Карельского научного центра РАН. 2010. № 2. С. 27–31.
Barcelos M., Lupki F.B., Campolina G.A., Nelson D.L. The colours of Biotechnology: general overview and developments of White, Green and Blue areas // FEMS Microbiol. Lett. 2018. V. 365. № 21. P. 1–12. https://doi.org/10.1093/femsle/fny239
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Химия, науки о материалах