

Доклады Российской академии наук. Науки о Земле, 2021, T. 499, № 2, стр. 138-143
Палеопасцихниды из чернокаменской свиты верхнего венда Среднего Урала (Пермский край)
В. Д. Десяткин 1, *, А. В. Колесников 1, **, А. А. Римский 1, А. О. Сысоева 1, В. А. Терехова 1, Н. Б. Кузнецов 1, 2, А. В. Шацилло 2, 3, И. В. Латышева 1, Т. В. Романюк 3, академик РАН М. А. Федонкин 1
1 Геологический институт Российской академии наук
Москва, Россия
2 Институт земной коры Сибирского отделения Российской академии наук
Иркутск, Россия
3 Институт физики Земли им. О.Ю. Шмидта Российской академии наук
Москва, Россия
* E-mail: desiatkin@ginras.ru
** E-mail: kolesnikov@ginras.ru
Поступила в редакцию 27.04.2021
После доработки 28.04.2021
Принята к публикации 28.04.2021
Аннотация
В типовом для вендской биоты (проблематичные дисковидные отпечатки и арумбериеморфные текстуры) Среднего Урала местонахождении Красная Горка (район Широковского водохранилища на р. Косьва, восток Пермского края), открытом Ю.Р. Беккером еще в 1972 г., в 2020–2021 гг. впервые обнаружены остатки ископаемых вендских организмов группы палеопасцихнид. В стратиграфическом смысле местонахождение приурочено к чернокаменской свите сылвицкой серии верхнего венда. Результаты морфометрического анализа находок палеопасцихнид из местонахождения Красная Горка позволили выделить три морфологических типа остатков: Palaeopascichnus delicatus, Yelovichnus gracilis и Palaeopascichnus linearis. Обнаружение большого количества остатков палеопасцихнид не только расширило представление об их палеобиологии и тафономии, но также показало, что палеонтологический потенциал типового местонахождения и одних из самых представительных разрезов чернокаменской свиты до настоящего времени был раскрыт не полностью.
Палеопасцихниды – одна из наиболее распространенных групп ископаемых макроостатков вендского или эдиакарского возраста. Они широко известны из разрезов венда юго-восточного Беломорья, Вычегодского бассейна, Мезенской и Московской синеклиз, Приладожья, Подольского Приднестровья, Среднего и Южного Урала, Оленекского поднятия Сибири, Учуро-Майского бассейна Дальнего Востока и эдиакария о. Ньюфаундленд, Уэльса Англии, п-ова Дигермулен севера Норвегии, южной части Китая и Австралии [1–12]. Палеопасцихниды характерны для крупнейших местонахождений ископаемых организмов эдиакарского типа в указанных регионах. Именно поэтому их присутствие в ассоциации окаменелостей низкого таксономического разнообразия в местонахождении Красная Горка дает основание прогнозировать открытие в изучаемом районе вендской биоты высокого разнообразия. В разное время их интерпретировали как ископаемые следы или продукты жизнедеятельности, остатки водорослей или животных неясного систематического положения [1–3, 13]. Результаты недавних исследований показали, что палеопасцихниды имели камерное строение и внешний скелет, который строился путем агглютинации зерен осадка из окружающего пространства [8]. Однако остается еще много вопросов, связанных с интерпретацией и систематикой этих палеонтологических остатков, которые демонстрируют настолько широкий диапазон типов сохранности, что до сих пор не существует унифицированного диагноза группы. Несмотря на то что пока не все ископаемые остатки монографически описаны, так же, как и сама группа палеопасцихнид, к ним предлагают относить таксоны Palaeopascichnus linearis, Palaeopascichnus delicatus, Yelovichnus gracilis и Orbisiana simplex [12].
Вендская ископаемая биота на Среднем Урале впервые была открыта Ю.Р. Беккером в 1972 г. в породах сылвицкой серии на правом берегу Широковского водохранилища (рис. 1), приблизительно в 3.5 км на юго-восток от устья р. Нюр (Няр) и непосредственной близости к урочищу Красная Горка [13]. Для обнаруженных Ю.Р. Беккером вендских палеонтологических остатков характерно низкое таксономическое разнообразие. Они представлены главным образом дисковидными образованиями с концентрическими желобками и складками, описанными как отпечатки медузоидных организмов рода Tirasiana. В последние годы такие формы все чаще считают отпечатками микробиальных колоний [14]. Вместе с тем дисковидные окаменелости, нередко объединяемые под общим названием Aspidella, подчас имеют весьма сложное строение [16] и являются частью (прикрепительным органом) крупных организмов, например, Charnia, Charniodiscus и др. Помимо этого, из типового местонахождения Красная Горка ранее были описаны отпечатки бесскелетных организмов Arumberia banksi, происхождение которых долгое время оставалось дискуссионным [15], пока не были обнаружены современные аналоги таких форм, являющиеся разновидностью микробиально-индуцированных осадочных текстур [17]. В 2020–2021 гг. мы провели рекогносцировочные исследования по берегам Широковского водохранилища, повторили находки Ю.Р. Беккера и в типовом местонахождении (рис. 1б, 1в) вендской биоты Красная Горка впервые обнаружили многочисленные и разнообразные остатки ископаемых организмов группы палеопасцихнид, приуроченных к средней части разреза чернокаменской свиты (рис. 1 а).
Рис. 1.
а – сводный разрез чернокаменской свиты верхнего венда Среднего Урала. б, в – географические карты расположения местонахождений остатков палеопасцихнид на восточном обрамлении Восточно-Европейской платформы: 1 – р. Сылвица (Свердловская обл.); 2 – Башкирийский мегаантиклинорий (Респ. Башкортостан и Челябинская обл.); 3 – р. Косьва, Широковское вдхр. (Пермский край). Представители ископаемых остатков палеопасцихнид из местонахождения Красная Горка: г–е – Palaeopascichnus delicatus; ж–и – Yelovichnus gracilis; й–л – Palaeopascichnus linearis. Условные обозначения: 1 – тонкослоистые алевролиты; 2 – чередующиеся аргиллиты и алевролиты; 3 – чередующиеся аргиллиты, алевролиты и песчаники; 4 – переслаивающиеся песчаники и аргиллиты; 5 – переслаивающиеся песчаники; 6 – косослоистые песчаники; 7 – чередующиеся аргиллиты, алевролиты и песчаники; 8 – чередующиеся аргиллиты и алевролиты; 9 – косослоистые песчаники и алевролиты; 10 – ровно- и волнистослоистые песчаники и алевролиты; 11 – стратиграфическое положение вулканических пеплов и их U–Pb-возраст.
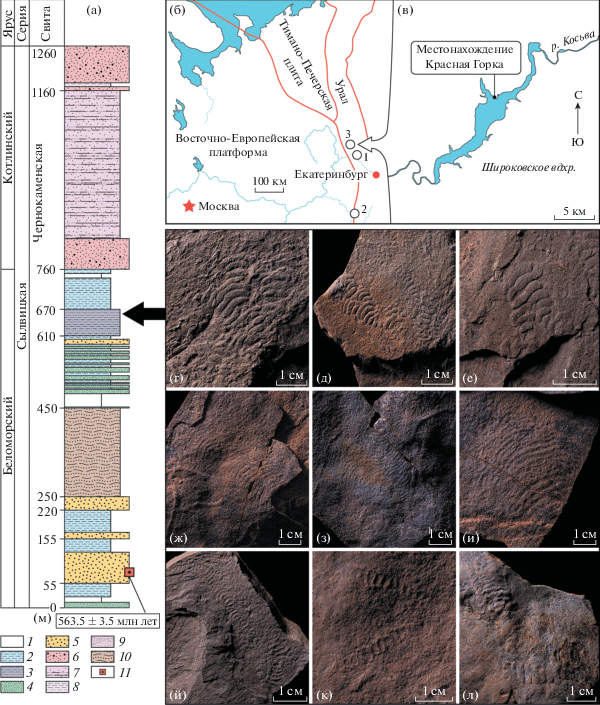
Собранная нами коллекция состоит из 96 экземпляров палеопасцихнид, остатки которых сохранились в виде однорядных раковин, состоящих из глобулярных, вытянутых, фасолевидных или аллантоидных (выгнутой формы) камер, расположенных цепочкообразно друг за другом (рис. 1г–1л). Палеонтологические остатки демонстрируют разную степенью сохранности на поверхностях напластования мелкозернистых песчаников и алевролитов: раковины представлены цельными камерами (рис. 1г–1и), частично разрушенными или сколлапсированными (рис. 1й–1л). У некоторых экземпляров отчетливо видно разделение раковин на два или три ответвления. Ветвящиеся формы характерны для экземпляров любых размеров от самых мелких до самых крупных. Ширина камер изменяется от 2 до 20 мм и может оставаться в раковине относительно постоянной или последовательно увеличиваться. Количество камер в раковине может достигать 15. Длина раковин варьирует от 5 мм до 10 см. Изучение фрагментов камер с помощью сканирующего электронного микроскопа показало наличие агглютинированной стенки, сложенной более крупнозернистым материалом, чем вмещающая порода, с повышенной концентрацией кристаллов пирита и кварца (рис. 2). Этот факт подтверждает наличие внешнего агглютинированного скелета у организмов группы палеопасцихнид, который ранее был установлен при изучении идентичных остатков из хатыспысткой свиты Оленекского поднятия и верховской свиты Онежского полуострова [8].
Рис. 2.
Строение и состав стенок камер палеопасцихнид под сканирующим электронным микроскопом Phenom XL (ГИН РАН, Москва). а – экземпляр Palaeopascichnus linearis и область проведения анализа; б – изображение стенки камер (показаны стрелками) и вмещающей породы, полученное в обратно-рассеянных электронах. Элементное картирование области с стенкой камеры и вмещающей породой: в – кислород; г – алюминий; д – кремний; е – калий; ж – железо; з – сера.

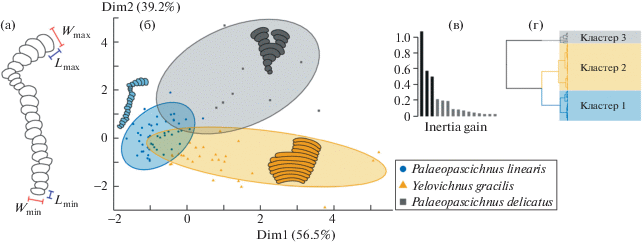
Представительная коллекция палеопасцихнид из местонахождения Красная Горка позволила провести замеры их основных морфометрических характеристик (рис. 3 а): длина и ширина начальной (Lmin, Wmin) и конечной камер (Lmax, Wmax), а также общее количество камер в раковине (N). На основе прямых измерений были выбраны параметры, отражающие морфологическую изменчивость ископаемых организмов: отношение ширины между начальной и конечной камерами (Wratio) и коэффициент удлинения камер (отношение длины и ширины у начальных и конечных камер соответственно – Kmin, Kmax). Морфометрический анализ палеопасцихнид проведен по методу главных компонент и иерархической кластеризации данных на главные компоненты (HCPC, hierarchical clustering on principal components) с помощью программы, написанной в языковой среде R версии 4.0.3 (R Core Team 2020), и ПО RStudio версии 1.4.1103 (RStudio, PBC 2021). Этот метод позволил проанализировать массив морфометрических характеристик путем уменьшения размерности многомерного набора данных и сглаживания случайных флуктуаций в формах и размерах камер палеопасцихнид для последующего построения и визуализации кластерных структур, отражающих морфологическое разнообразие ископаемых организмов. На рис. 3б представлен результат PCA-анализа по трем параметрам Wratio (39.2% от общей дисперсии значений), Kmax и Kmin (56.5% соответственно). Установлено, что пространство PCA делится на три кластера (рис. 3 в–г): (кластер 1) – палеопасцихниды с относительно постоянным размером и формой камер от глобулярной до слабо вытянутой (Kmax не превышает значение 1.75, Wratio колеблется от 1.0 до 1.5); (кластер 2) – палеопасцихниды с относительно постоянным размером, но сильно вытянутой формой камер от фасолевидной до аллантоидной (Kmin и Kmax достигают значений 5.5, Wratio не превышает 2.0); (кластер 3) – палеопасцихниды с постоянно увеличивающейся шириной камер от почти глобулярной в начале до сильно вытянутой и дугообразной в конце (Kmax достигает 3.75, Wratio доходит до 4.5–4.75). Сравнение с ранее опубликованными данными показывает, что для первого кластера характерен таксон Palaeopascichnus linearis [8], для второго – Yelovichnus gracilis [2], и для третьего – Palaeopascichnus delicatus [1]. Похожий результат был получен при изучении морфологического разнообразия палеопасцихнид методами PCA и HCPC из формации Фермюс эдиакария о. Ньюфаундленд, Канада [10]. Стоит отметить, что таксон Yelovichnus gracilis пока не относится к роду Palaeopascichnus и ожидает ревизии, однако, многими исследователями ранее высказывалось мнение о его принадлежности к группе палеопасцихнид [10–12].
Рис. 3.
Морфологический анализ палеопасцихнид из чернокаменской свиты методами PCA и HCPC: а – схематичное изображение формы палеопасцихниды и замеры их основных морфометрических характеристик; б – визуализация результатов кластерного анализа форм палеопасцихнид; в – распределение разброса коэффициентов инерции (inertia gain) в пространстве главных компонент (PCA) подтверждает разделение форм палеопасцихнид на три кластера; г – результат иерархической кластеризации данных на главные компоненты (HCPC) по трем кластерам (морфотипам).
Вместе с палеопасцихнидами в разрезе чернокаменской свиты были обнаружены многочисленные слепки органов прикрепления и отпечатки концентрических микробиальных колоний, белтанелломорфных, рангееморфных и дикинсониеморфных организмов. Совокупность литологических типов осадочных пород и их текстурный облик позволяют интерпретировать условия осадконакопления фоссилиеносного уровня чернокаменской свиты в изученном местонахождении как крайне мелководные приливно-отливные. Литологически и палеонтологически сходный уровень ранее был обнаружен в верхней части коноваловской подсвиты чернокаменской свиты, вскрытый в нижнем течении р. Сылвицы [18, 19] примерно в 110 км к юго-юго–востоку от местонахождения Красная горка. Это допускает предположить, что в местонахождении Красная Горка тоже обнажена коноваловская подсвита. Кроме того, полученный недавно U–Pb-возраст 563.5 ± 3.5 млн лет цирконов из вулканических туфов, обнаруженных в одном из фрагментов разреза сылвицкой серии, позволяет отнести находки палеопасцихнид к беломорскому региоярусу венда на Среднем Урале [20]. До настоящего времени остатки палеопасцихнид в восточном обрамлении Восточно-Европейской платформы были известны из сылвицкой серии Среднего Урала (обнажения перевалокской и чернокаменской свит по р. Сылвицы) и ашинской серии Южного Урала (обнажения басинской и зиганской свит) [4, 6]. Таким образом, разрезы чернокаменской свиты на Широковском водохранилище представляют новое местонахождение палеопасцихнид на Среднем Урале. Результаты морфометрического анализа и подтверждение наличия агглютинированного скелета в строении относимых к ним фоссилий дают новое понимание природы и таксономического разнообразия проблематичных остатков и раскрывают возможный потенциал применения группы палеопасцихнид для биостратиграфии вендских отложений.
Список литературы
Палий В.М. Остатки бесскелетной фауны и следы жизнедеятельности из отложений верхнего докембрия и нижнего кембрия Подолии // Палеонтология и стратиграфия верхнего докембрия и нижнего палеозоя юго-запада Восточно-Европейской платформы. 1976. Киев: Наукова думка. С. 63–77.
Федонкин М.А. Систематическое описание вендских Metazoa. Вендская система. Т. 1. Палеонтология. М.: Наука, 1985. С. 70–106.
Antcliffe J.B., Gooday A.J., Brasier M.D. Testing the Protozoan Hypothesis for Ediacaran Fossils: A Developmental Analysis of Palaeopascichnus // Palaeontology. 2011. V. 23 (5). P. 1157–1175.
Гражданкин Д.В., Маслов А.В. и др. Осадочные системы сылвицкой серии (верхний венд Среднего Урала). Екатеринбург: УрО РАН, 2010. 280 с.
Wan B., Xiao S., et al. Orbisiana linearis from the Early Ediacaran Lantian Formation of South China and its Taphonomic and Ecological Implications // Precambrian Research. 2014. V. 255. P. 266–275.
Kolesnikov A.V., Marusin V.V., et al. Ediacaran Biota in the Aftermath of the Kotlinian Crisis: Asha Group of the South Urals // Precambrian Research. 2015. V. 263. P. 59–78.
Nagovitsin K.E., Rogov V.I., et al. Revised Neoproterozoic and Terreneuvian Stratigraphy of the Lena-Anabar Basin and North-western Slope of the Olenek Uplift, Siberian Platform // Precambrian Research. 2015. V. 270. P. 226–245.
Kolesnikov A.V., Rogov V.I., et al. The Oldest Skeletal Macroscopic Organism Palaeopascichnus linearis // Precambrian Research. 2018. V. 316. P. 24–37.
Kolesnikov A.V., Liu A.G., et al. A Reassessment of the Problematic Ediacaran Genus Orbisiana Sokolov 1976 // Precambrian Research. 2018. V. 316. P. 197–205.
Hawco J.B., Kenchington C.H., McIlroy D. A Quantitative and Statistical Discrimination of Morphotaxa within the Ediacaran Genus Palaeopascichnus // Papers in Palaeontology. 2019. https://doi.org/10.1002/spp2.1290
Jensen S., Hoegstroem A.E.S., et al. New Occurrences of Palaeopascichnus from the Stahpogieddi Formation, Arctic Norway, and their Bearing on the Age of the Varanger Ice Age // Canadian J. Earth Sci. 2018. V. 55. P. 1–10.
Kolesnikov A.V. Stratigraphic Correlation Potential of the Ediacaran Palaeopascichnids // Estudios Geologicos. 2019. V. 75 (2), e102. P. 1–5.
Беккер Ю.Р. Первые палеонтологические находки в рифее Урала // Изв. АН СССР. Сер. геол. 1977. № 3. С. 90–100.
Grazhdankin D., Gerdes G. Ediacaran Microbial Colonies // Lethaia. 2007. V. 40. P. 201–210.
Колесников А.В., Маслов А.В., Гражданкин Д.В. Арумбериеморфные текстуры в верхнем венде Урала // ДАН. 2012. Т. 447. № 1. С. 66–72.
Luzhnaya E., Ivantsov A. Skeletal Nets of the Ediacarian Fronds // Paleontological Journal. 2019. V. 53. P. 667–675.
Kolesnikov A.V., Danelian T., et al. Arumberiamorph Structure in Modern Microbial Mats: Implications for Ediacaran Palaeobiology // Bull. Soc. géol. Fr. 2017. V. 188 (1–2). P. 1–10.
Sozonov N.G., Bobkov N.I., et al. The Ecology of Dickinsonia on Tidal Flats // Estudios Geologicos. 2019. V. 75 (2), e116. P. 1–5.
Bobkov N.I., Kolesnikov A.V., et al. The Occurrence of Dickinsonia in Non-marine Facies // Estudios Geologicos. 2019. V. 75 (2), e096. P. 1–5.
Кузнецов Н.Б., Белоусова Е.А. и др. Результаты геохронологического и изотопно-геохимического изучения циркона из туфов сылвицкой серии (западный склон Среднего Урала): к происхождению пепловых прослоев в вендских толщах Восточно-Европейской платформы // ДАН. 2017. Т. 473. № 3. С. 341–345.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле