

Доклады Российской академии наук. Науки о Земле, 2021, T. 499, № 2, стр. 185-192
Диагностика начала аграрного освоения северо-запада Восточно-Европейской равнины по споро-пыльцевым спектрам
Член-корреспондент РАН А. А. Тишков 1, *, Н. Г. Царевская 1, Е. Ю. Новенко 1, Е. А. Белоновская 1
1 Институт географии, Российская академия наук
Москва, Россия
* E-mail: tishkov@igras.ru
Поступила в редакцию 04.05.2021
После доработки 12.05.2021
Принята к публикации 13.05.2021
Аннотация
Рассматриваются вопросы диагностики по споро-пыльцевым спектрам начала аграрного освоения хвойно-широколиственных лесов северо-запада Европейской России и его специфики относительно агрогенной трансформации зонального лесного покрова и формирования на его месте лесо-поле-лугового ландшафта. Показано, что сельскохозяйственное освоение лесов рассматриваемого региона, согласно материалам споро-пыльцевого анализа по ключевым разрезам, начинает проявляться с 2300–2600 кал. л.н., а после 1500 кал. л.н., судя по непрерывному присутствию в диаграммах пыльцы культивируемых растений, оно становится повсеместным и необратимым. Приведены соответствующие показатели и виды растений, наличие пыльцы которых в диаграммах позволяет индицировать время и характер агрогенных изменений растительности.
ВВЕДЕНИЕ
Климатогенные и антропогенные изменения растительности в голоцене, которые можно надежно индицировать и реконструировать по изменениям состава и соотношений основных компонентов споро-пыльцевых спектров, как известно, имеют разное “характерное время” и “характерное пространство” проявления в ландшафте [1]. Первые, если это касается критических изменений среднегодовых температур и последующих зональных перестроек растительного покрова, проявляются как “географические смены” и сдвиги зональных и подзональных границ растительности. Их характерное время – сотни и первые тысячи лет. Они позволяют в умеренном климате выявлять последовательные смены лесных пород, а по периферии лесной зоны – отмечать расширение или сокращение переходной полосы от леса к степи или к тундре. А вторые, на фоне возможных изменений климата, будут демонстрировать дигрессивно-демутационные процессы с характерным временем в десятки и первые сотни лет. Это относится только к вторичным восстановительным сукцессиям лесов, так или иначе связанным с деятельностью человека – вырубкой и выжиганием, распашкой, перелогой, поддержанием послелесной луговой стадии за счет сенокошения и выпаса и забрасыванием пашни. Первичные сукцессии растительности по параметрам “характерного времени” сопоставимы с вековыми и тысячелетними климатическими циклами и протекают на их фоне, который, в сущности, определяет их конечные стадии – зональный климакс, как это было на северо-западе Восточно-Европейской равнины по мере становления здесь зональной растительности и эволюции озерно-болотных комплексов после голоценового оптимума.
В сообщении рассматриваются вопросы диагностики по пыльцевым спектрам начала аграрного освоения хвойно-широколиственных лесов северо-запада Европейской России и его специфики относительно изменения пространственной структуры зонального лесного покрова и его антропогенных модификаций, свойственных лесо-поле-луговому ландшафту так называемых “поозерий” – Валдайского, Псковского, Смоленского, Белорусского и др. Можно полагать, что эта полоса вдоль “конечно-моренной дуги”, окаймляющей Балтийское море, еще в начале раннего железного века (конец I тыс. до н.э. – первая половина I тыс. н. э.) стала ареной фронтального аграрного освоения лесов, достигшего максимума в период около 2000 кал. л.н., в так называемый “Римский оптимум” [2]. Уточнению начала необратимых агрогенных изменений лесов здесь по имеющимся споро-пыльцевым диаграммам и посвящена данная работа.
В качестве одной из гипотез в основе статьи рассматривается положение, что раннее сельскохозяйственное освоение данных территорий имело “очаговый” характер, свойственный, например, финно-угорским народам, и не находило явного “индикационного” отражения в споро-пыльцевых спектрах. Только “фронтальное” волнообразное освоение лесов с оборотом их выжигания, расчистки и забрасывания, меньшим, чем период восстановительной сукцессии зональных лесов, привел к коренной перестройке структуры растительного покрова. В рассматриваемом регионе она происходила в период, предшествовавший “великому переселению народов”, и отразилась в большинстве выявленных споро-пыльцевых спектров опосредованно через необратимое изменение соотношения компонентов, трендов доли их участия и появления пыльцы и спор новых групп растений, связанных с примитивным аграрным производством [2–7].
Ранее нами было показано [8], что сохранившиеся разновозрастные леса и другие элементы лесо-поле-лугового ландшафта Валдайской возвышенности имеют близкую наследованную антропогенную фрагментированность, составляющую около 2 га – соответствующую контуру первичной подсечно-огневой расчистки под пашню. Ее выявление посредством анализа спектральных снимков Landsat позволило определить, что основные факторы антропогенной фрагментации и мелкоконтурности действовали в период доминирования именно подсечно-огневой системы землепользования, задолго до появления в рассматриваемом регионе постоянных поселений и аграрных угодий вокруг них, когда аграрная нагрузка стала более концентрированной в пространстве (поля и луга), а в отношении лесного покрова ослабла [9].
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
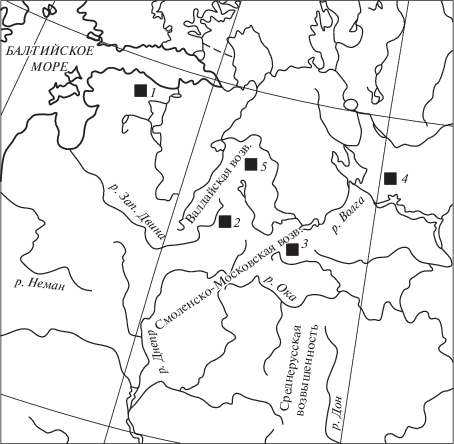
В качестве объектов анализа, синтеза и интерпретации результатов индикации агрогенных изменений растительности рассматриваемого региона, которые прослеживаются в споро-пыльцевых диаграммах из многочисленных разрезов, выбраны такие хорошо датированные объекты (рис. 1): 1 – оз. Райгаствере в Эстонии [10]; 2 – болото Старосельский мох в Центрально-лесном заповеднике на юге Валдайской возвышенности в Тверской области [11]; 3 – оз. Галичское в Костромской области [12]; 4 – оз. Долгое в Московской области [13]; 5 – болото Ольгино в национальном парке Валдайский в Новгородской области [14].
Рис. 1.
Географическое положение ключевых разрезов, рассмотренных в тексте: 1 – оз. Райгаствере, Эстония; 2 – болото Старосельский мох, Тверская область; 3 – оз. Долгое, Московская область; 4 – оз. Галичское, Костромская область; 5 – болото Ольгино, Новгородская область.
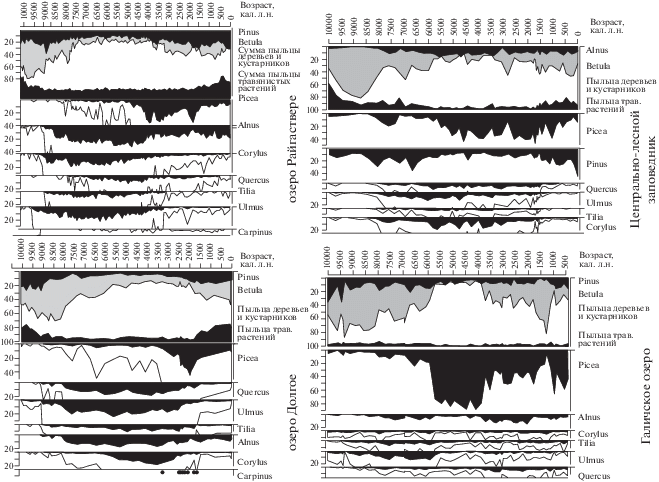
Имеющиеся радиоуглеродные датировки позволили построить достоверные модели скорости накопления отложений в разрезах, представленных в анализируемых публикациях, и споро-пыльцевые диаграммы, в которых по оси ординат представлен возраст отложений (рис. 2_1–2_4). Калибровка радиоуглеродных дат и построение модели “возраст–глубина разреза” проведены при помощи программы Bchron. Обработка данных анализа разрезов и построение споро-пыльцевых диаграмм проводились с помощью программы TILIA TILIA-Graph.
Рис. 2.
Спорово-пыльцевые диаграммы разрезов: 1 – оз. Райгаствере, 2 – болото Старосельский мох, 3 – оз. Долгое, 4 – оз. Галичское. В пыльце древесных растений дополнительный контур показывает увеличение базового таксона в 10 раз.
Более детально скорость накопления отложений в разрезе прослежена и на примере нашего разреза “Болото Ольгино” в Новгородской области, заложенного еще при участии Л.Р. Серебрянного и В.А. Климанова (нижняя датировка для глубины 648 см – 12 100 ± 270 л.н., № GIN-9018). По нашим данным [14], в позднеледниковье скорость осадконакопления здесь была минимальной и составляла 0.075 мм/год. В пребореале – 0.15 мм/год, а в бореале достигла 1.15 мм/год. В атлантический период скорость накопления замедлилась – 0.71 мм/год, в суббореальный период – 0.75 мм/год, в начале субатлантического периода – около 0.72 мм/год, в его середине – 0.56 мм/год и за последние 200 лет (исходя из самой “свежей” датировки 190 ± 60 л.н.) скорость торфонакопления была максимальной – около 4.25 мм/год.
К облигатным диагностическим признакам агрогенных изменений лесной зоны Восточно-европейской равнины в споро-пыльцевых спектрах позднего голоцена могут быть отнесены:
– увеличение в целом доли пыльцы травянистых растений (как отражение расширения площадей, занятых послелесной луговой растительностью на водоразделе);
– рост доли спор зеленых мхов-пирофитов, единственной группы споровых растений, обильно продуцирующих споры; лесные виды мохообразных, например pp. Pleurozium, Hylocomium, Ptilium, Dicranum спороносят редко, зато обильно спороносящие первые десятилетия после выжигания леса Leptobryum, Polytrichum, Funaria и др.;
– сокращение доли пыльцы ели в регионе и широколиственных пород (дуба, липы, клена), конкретно в районе расположения объекта, как индикатор снижения площадей формируемых ими лесов на благоприятных для создания пашни территориях;
– рост участия березы, серой ольхи и сосны как пионерных пород на месте коренных хвойных и хвойно-широколиственных лесов после их сведения под пашню;
– появление в составе споро-пыльцевого спектра пыльцы культивируемых растений, сорняков и других растений-индикаторов нарушенных земель, пашни и пр.;
– наличие пыльцы видов растений эвтрофных местообитаний (p. Urtica), малосъедобных и ядовитых растений для развития опушечного и лесного (подкронового) выпаса коров и свиней;
– рост доли пыльцы растений-индикаторов пастбищного использования лугов – индикаторов уплотнения почвы (Polygonum, Plantago), а также ядовитых и малосъедобных растений (Ranunculus, Rumex, Calamagrostis, Deschampsia);
– в случае анализа озерных отложений с участием макрофитов за счет их появления и доминирования, можно фиксировать начальные стадии агрогенного евтрофирования в местах усиления стоков с полей.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Палинологические исследования разрезов голоцена лесной зоны Восточно-Европейской равнины (рис. 2) позволяют выделить этапы естественных изменений растительности, обусловленных климатическими изменениями [3]. В раннем голоцене, в бореальном периоде (10 300–8800 кал. л.н.) рассматриваемая территория была покрыта березовыми лесами. Потепление климата в течение атлантического периода (8800–5700 кал. л.н.) способствовало расселению здесь дуба, вяза, липы, ольхи и лещины и формированию зональных широколиственных, хвойно-широколиственных и неморальных еловых лесов. Несмотря на похолодание климата в суббореальное время, в течение периода 5700–4000 кал. л.н., роль широколиственных пород в лесных формациях оставалась достаточно высокой. В то же время увеличение содержания пыльцы ели в спектрах из разрезов в Прибалтике, на Валдайской возвышенности (разрез Старосельский мох, Ольгино болото) и в бассейне Верхней Волги отражает активную экспансию ели – “верхний максимум ели” в голоцене по Н.А. Хотинскому [3]. В Верхнем Поволжье, как показывают данные по стратиотипическому разрезу голоцена – Половецко-Купанскому болоту [15] и Галичскому озеру [12], климатогенная деградация широколиственных сообществ была даже более глубокой. На этой территории господствующими растительными формациями стали южно-таежные ельники. На Смоленско-Московской возвышенности (разрез оз. Долгое, [13]) и в бассейне Верхней Оки [16] сохранялись широколиственные леса из дуба, вяза и липы с густым подлеском из лещины.
Существенные изменения растительности выявлены в интервале 4000–3000 кал. л.н. (средний суббореал) и связаны с сокращением доли пыльцы ели и увеличением содержания широколиственных пород (липы, дуба и вяза) и березы. В период после 3000 кал. л.н. еловые леса восстановили свои позиции на Валдае и в Верхнем Поволжье и расширялись к югу. Хорошо выраженный максимум пыльцы ели (до 50%) и сокращение участия пыльцы дуба, вяза и орешника на диаграмме оз. Долгое на Смоленско-Московской возвышенности относятся к интервалу 2600–1800 кал. л.н.
Существенные изменения спорово-пыльцевых спектров происходят почти синхронно в рассмотренных разрезах и относятся к рубежу 1500 кал. л.н., и это, очевидно, может быть связано с влиянием хозяйственной деятельности человека. Выше этой хроностратиграфической границы в спектрах зафиксированы резкое сокращение участия пыльцы ели и снижение почти до нуля содержания пыльцы широколиственных пород, увеличение процентных соотношений пыльцы березы и сосны (пионерных пород деревьев). Особенно ярко уничтожение широколиственных лесов проявилось по пыльцевым данным разрезов, расположенных на возвышенных моренных равнинах – Валдайской и Среднерусской возвышенностях. В настоящее время хвойно-широколиственные леса и ельники неморальной группы занимают наиболее благоприятные для земледелия участки, и на заре становления сельского хозяйства в регионе они уничтожались в первую очередь [7, 17].
После 1500 кал. л.н. кривая пыльцы культурных злаков в рассмотренных разрезах становится непрерывной, существенно увеличивается содержание пыльцы травянистых растений, в том числе подъем кривых Poaceae, Chenopodiaceae и Artemisia. Появляются такие индикаторы нарушенных грунтов и поселений человека, как виды рр. Plantago, Rumex, Polygonum и Urtica. Следует отметить, что единичные пыльцевые зерна культурных злаков Cerealia и сорных (например, Asteracea) видов отмечены на спорово-пыльцевых диаграммах начиная с 2300–2600 кал. л.н. Это позволяет предположить, что земли обрабатывались под посевы раньше – в течение всего субатлантического периода.
Однако, согласно полученным при синтезе споро-пыльцевых диаграмм данным, массовое сведение лесов началось только в последние две тысячи лет. В северо-восточных районах Восточно-Европейской равнины, особенно там, где хозяйственное освоение было не столь интенсивным, как в центральных районах, хвойные леса частично восстановились. Об этом свидетельствует увеличение участия пыльцы ели в спектрах отложений Галичского озера. В центре Европейской части России и в Прибалтике верхние горизонты разрезов характеризуются высоким содержанием пыльцы березы, сосны и трав. Участие пыльцы ели и широколиственных пород остается низким, а в отложениях, соответствующих последним 400–500 лет, доля этих таксонов еще уменьшается. Для этого же временного интервала выявлена максимальная концентрация пыльцы культивируемых растений, луговых трав и сорняков. Подобные изменения спорово-пыльцевых спектров свидетельствуют о том, что в течение последних столетий естественные растительные сообщества постепенно были уничтожены на больших территориях и их место заняли вторичные леса и сельскохозяйственные угодья.
В итоге анализ споро-пыльцевых диаграмм на северо-западе Восточно-Европейской равнины позволил уточнить “характерное время” и некоторые индикаторы агрогенной динамики растительности (табл. 1) для периода последних 2000 лет, когда климатогенная динамика перестала доминировать.
Таблица 1.
“Характерное время” и индикаторы агрогенной динамики растительности в споро-пыльцевых диаграммах голоцена северо-запада Восточно-Европейской равнины
| Форма антропогенной трансформации | Дигрессивно-демутационные процессы | Индикаторы | “Характерное время” |
|---|---|---|---|
| Сплошная рубка, выжигание леса, расчистка | Вторичная сукцессия, начало формирования антропогенных модификаций растительных сообществ – агрогенных, пирогенных, пасквальных | Рост доли пыльцы Pinus sylvestris, Betula pendula, Alnus incana; сокращение доли пыльцы Quercus robur, других широколиственных пород и Picea abies | Продолжительность: (1) замещения коренных пород (дуб, ель) на вырубках вторичными породами (сосна, береза, осина, ольха) – 30–50 лет; (2) полного цикла вторичной сукцессии – 80–120 лет; (3) начало плодоношения молодняка на залежи (выделения пыльцы): сосна – 20, ель – 20–30, береза – 15, дуб – 30, липа и клен – 20 |
| Режим пашни и перелоги | Цикл перелоги с восстановлением растительности до длительно-производной (луговой) стадии | Пыльца культивируемых растений – злаков рр. Secale, Triticum, Hordeum, Panicum, Avena, овощей – рр. Brassica (редко культивировались до образования пыльцы), сорняков и видов открытых грунтов (из Brassicaceae, Asteraceae и др.) | За счет фронтального освоения территории под пашню (режим 10–15 лет) и параллельного существования перелоги (до 20–30 лет) фиксируется сначала как “событие”, а потом как постоянно действующий фактор и элемент структуры растительного покрова |
| Режим послелесного лугового сенокоса и выпаса (коров, лошадей) | “Управляемое” с помощью сенокошения, выпаса и палов существование послелесных лугов – кормовых угодий (обязательного элемента аграрного хозяйства | Доминирование пыльцы луговых злаков и разнотравья – Festuca, Poa, Polygonum, Taraxacum officinale Plantago, Trifolium, Rumex; на сырых местах – малосъедобных и ядовитых растений рр. Filipendula, Carex, Ranunculus, Deschampsia | Сначала как элемент перелоги, а в дальнейшем как постоянно действующий фактор. Цикл демутации вторичного леса “по лугу” – 30–50 лет, коренного леса – 80–120 лет |
| Режим подкронового выпаса в лиственных лесах (коров, свиней) и заготовка веточно-листового корма для скота | Эвтрофирование леса, уничтожение подроста и подлеска, дигрессия первичных (дубрав) и вторичных (травяные березняки) лесов | Urtica sp.sp., рост доли пыльцы непоедаемых и ядовитых растений – Ranunculus, Rumex, Deschampsia, Calamagrostis, Pteridium, исчезновение пыльцы Corylus и других видов подлеска, образующих пыльцу | Постоянно действующий фактор “волн” плодоношения не только по климатическим причинам (засухи), но и за счет эвтрофирования (больше плюсовых деревьев). Переложное использование выпасов совпадало и с циклами плодоношения дуба (15–20–30 лет) |
| Сельскохозяйственные стоки с полей и пастбищ | Эвтрофирование прибрежной полосы озер и влажных западин, развитие полосы макрофитов, образование сплавины на мелководьях озер (феномен “второго” после раннего голоцена всплеска образования болот) | Potamogeton, Lemna, Phragmites, Scirpus, Hydrocháris, Nuphar, Equisetum, Typha, Comarum, Carex, а из мохообразных – виды рр. Calliergon, Sphagnum (например, Sph. obtusum), Drepanocladus и др. | Цикл в случае нового заболачивания охватывает около 2000 лет с короткой – несколько сот лет – “эвтрофной” фазой. Эвтрофирование обводненной каймы мелкоконтурного верхового болота охватывает также несколько сот лет и находит отражение в пыльцевой диаграмме |
| Выжигание леса, лесные и луговые палы, палы для лесной пастьбы | Постпирогенные сукцессии, формирование пирогенных субклимаксов, например сосняков – брусничных и беломошников | Виды Bryales и Hepaticae– pp. Funaria, Polytrichum, Marchantia. Для лугов с частыми палами – виды рр. Calamagrostis, Rumex, Equisetum | Восстановление вторичного леса на гари 30–50 лет, коренного леса – 100–120 лет. “Оборот огня”, не позволяющий восстанавливаться под пологом сосняка дубравы и ельника – 30–50 лет, для лугов – раз в несколько лет |
ЗАКЛЮЧЕНИЕ И ВЫВОДЫ
Для конечно-моренных ландшафтов северо-запада Восточно-Европейской равнины все признаки выделения антропогенных изменений растительности по споро-пыльцевым диаграммам, за исключением явного присутствия пыльцы культурных растений, справедливы только при анализе постоптимальных фаз голоцена, когда природная зональность приобрела современную структуру.
Агрогенная трансформации лесов рассматриваемого региона, согласно материалам споро-пыльцевого анализа (рис. 2), начинает проявляться с 2300–2600 кал. л.н., а после 1500 кал. л.н., судя по непрерывному присутствию в диаграммах пыльцы культивируемых растений, она становится повсеместной и необратимой.
Выявляемые индикаторы агрогенной трансформации растительности в спорово-пыльцевых спектрах (рис. 2; табл. 1) могут быть сгруппированы по следующим критериям: (1) соотношению сумм древесной пыльцы и пыльцы трав, отражающему пропорцию площадей каждого из элементов лесо-поле-лугового ландшафта, прежде всего, устойчивому присутствию в растительном покрове послелесных водораздельных лугов; (2) изменениям доли пыльцы внутри группы древесных пород, индицирующих уничтожение зональных лесов – дубрав и ельников – и их замещение вторичными лесами – березняками, сосняками, сероольшанниками, а также отражающих интенсивность массовых заготовок веточных кормов для скота; (3) устойчивое (в некоторых случаях “волнообразное”) появление в спектрах спор и пыльцы пирофитных растений – зеленых мхов, папоротника – Pteridium aquilinum, иван-чая (Onagraceae) и др.; (4) присутствие пыльцы, а с начала субатлантического периода появление непрерывной кривой концентрации культурных злаков Cerealia и других индикаторов пашенного земледелия, а также сорняков из Asteraceae (например, Cirsium, Centaurea) и Chenopodiaceae (Chenopodium album); (5) присутствие пыльцы луговых злаков и разнотравья из семейств Poaceae, Polygonaceae, Ranunculaceae, Fabaceae, Brassicaceae и Asteraceae, характерных для послелесных водораздельных лугов; (6) наличие пыльцы растений, индицирующих пасквальное уплотнение грунтов (Polygonum aviculare, Plantago major, Р. media) и пастбищный режим использования растительности, в нашем случае – рост в травостое доли малосъедобных и ядовитых растений – рр. Deschampsia, Calamagrostis, Carex, Ranunculus, Plantago lanceolata, Rumex acetosella, видов рр. Rubiaceae, Rosaceae.
Важным результатом исследований споро-пыльцевых диаграмм в нашем случае можно считать вывод о том, что рассмотрение по отдельности каждого из приведенных параметров, видов и групп растений в качестве индикаторов агрогенных изменений зональной растительности возможно лишь в совокупности с подробным анализом ее динамики и “характерного времени” проявления ее форм (табл. 1). Единичные находки пыльцы агрогенных индикаторов, включая пыльцу Secale, Triticum, Hordeum, Panicum, Avena, нельзя рассматривать признаком развития аграрного производства, а интерпретация становится возможной только с учетом совокупности индикационных признаков.
Список литературы
Тишков А.А. “Характерное пространство” и “характерное время” как ключевые категории биогеографии // Изв. РАН. Сер. геогр., 2016. № 4. С. 20–33.
Новенко Е.Ю. Изменения растительности и климата Центральной и Восточной Европы в позднем плейстоцене и голоцене в межледниковые и переходные этапы климатических макроциклов. М.: ГЕОС, 2016. 227 с.
Хотинский Н.А. Голоцен Северной Евразии. М.: Наука, 1977. 200 с.
Гуман M.A. Антропогенные изменения растительности юга Псковской области в голоцене (по палинологическим данным) // Бот. журн. 1978. Т. 63. № 10. С. 1415–1429.
Гуман М.А., Хотинский Н.А. Антропогенные изменения растительности центра Русской равнины в голоцене (по палинологическим данным) // Антропогенные факторы в истории развития современных экосистем. М., 1981. С. 7–19.
Антропогенные факторы в истории развития современных экосистем. М.: Наука. АН СССР, Науч. совет по пробл. Биосферы. Отв. ред. Л.Г. Динесман. 1981. 246 с.
Носова М.Б., Новенко Е.Ю., Зерницкая В.П., Дюжова К.В. Палинологическая индикация антропогенных изменений растительности восточно-европейских хвойно-широколиственных лесов в позднем голоцене // Изв. РАН. Сер. геогр. 2014. № 4. С. 35–43.
Белоновская Е.А., Кренке-мл. А.Н., Тишков А.А., Царевская Н.Г. Природная и антропогенная фрагментация растительного покрова Валдайского поозерья // Изв. РАН. Сер. геогр. 2014. № 5. С. 67–82.
Тишков А.А. Ландшафтная основа происхождения названия “Валдай” // Изв. РАН. Сер. геогр. № 1. 2014. С. 109–119.
Seppä H., Poska A. Holocene Annual Mean Temperature Changes in Estonia and Their Relationship to Solar Insolation and Atmospheric Circulation Patterns // Quaternary Research. 2004. V. 61. P. 22–31.
Novenko E.Yu., Volkova E.M., Nosova M.B., Zuganova I.S. Late Glacial and Holocene Landscape Dynamics in the Southern Taiga Zone of East European Plain According to Pollen and Macrofossil Records from the Central Forest State Reserve (Valdai Hills, Russia) // Quaternary International. 2009. P. 93–103.
Величко А.А., Кременецкий К.В., Негенданк Й., Минграм Й., Борисова О.К., Зеликсон Э.М., Новенко Е.Ю., Писарева В.В. Позднечетвертичная история растительности Костромского Заволжья по данным палинологического изучения донных осадков Галичского озера // Бюлл. Ком. по изуч. четв. пер. 2001. № 64. С. 5–20.
Kremenetski K.V., Borisova O.K., Zelikson E.M. The Late Glacial and Holocene History of Vegetation in the Moscow Region // Paleontological Journal. 2000. V. 34 (1). P. 67–74.
Климанов В.А., Кожаринов А.В., Тишков А.А. Палеогеоэкологические реконструкции динамики растительности и климата Валдайского поозерья в позднеледниковье и в голоцене // Тр. нац. парка “Валдайский”. Вып. 1. СПб., 2010. С. 254–261.
Khotinski N.A., Klimanov V.A. Alleröd, Younger Dryas and Early Holocene Palaeo- Environmental Stratigraphy // Quaternary International. 1997. V. 41/42. P. 67–70.
Носова М.Б., Новенко Е.Ю., Зерницкая В.П., Дюжова К.В. Палинологическая индикация антропогенных изменений растительности восточно-европейских хвойно-широколиственных лесов в позднем голоцене // Изв. РАН. сер. геогр. 2014. № 4. С. 35–43.
Носова М.Б. Центрально-лесной заповедник: естественно- и антропогенно-обусловленная динамика растительности в голоцене // Бюлл. МОИП. Отдел биол. 2008. Т. 113. № 2. С. 38–45.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле