

Доклады Российской академии наук. Науки о Земле, 2022, T. 503, № 1, стр. 26-31
Таксономическая принадлежность ювенильного гадрозавроидного динозавра из верхнемеловой баинширэинской свиты Монголии
А. О. Аверьянов 1, *, академик РАН А. В. Лопатин 2, Х. Цогтбаатар 3
1 Зоологический институт Российской академии наук
Санкт-Петербург, Россия
2 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
3 Институт палеонтологии Монгольской Академии наук
Улан-Батор, Монголия
* E-mail: dzharakuduk@mail.ru
Поступила в редакцию 01.11.2021
После доработки 10.11.2021
Принята к публикации 11.11.2021
- EDN: DJDUTD
- DOI: 10.31857/S2686739722030033
Аннотация
Почти полный сочлененный скелет динозавра из верхнемеловой (сеноман–сантон) баинширэинской свиты местонахождения Байшин-Цав в пустыне Гоби (Монголия) принадлежит ювенильному представителю гадрозавроидов. Этот образец группируется с Gobihadros mongoliensis из той же свиты того же местонахождения на 68% из 304 наиболее парсимоничных филогенетических деревьев, полученных с помощью программы филогенетического анализа PAUP. Тем не менее он четко отличается от Gobihadros более продвинутым озублением, демонстрирующим редукцию краевых зубчиков и отсутствие дополнительных гребней на коронках зубов. Ювенильная особь также отличается от Gobihadros треугольным ростральным отростком теменных костей, вклинивающимся между лобными костями, и намного более коротким заглазничным отростком скуловой кости. Этот экземпляр может представлять второй, ранее неизвестный таксон Hadrosauroidea в баинширэинской свите.
В 1971 г. Совместной советско-монгольской палеонтологической экспедицией были обнаружены и собраны многочисленные остатки гадрозавроидных динозавров, включая почти полный скелет ювенильной особи, в сеноман-сантонской баинширэинской свите местонахождения Байшин-Цав в пустыне Гоби, Монголия [1]. Указанный скелет (экз. ПИН, № 3458/5; рис. 1) был помещен в экспозицию Палеонтологического музея им. Ю.А. Орлова Палеонтологического института им. А.А. Борисяка РАН (ПИН) в Москве и в 1973–1974 гг. участвовал во временной выставке в Японии, где получил неофициальное название “Gadolosaurus” [2–4]. В экспозиции Палеонтологического музея в Москве этот скелет обозначен как Arstanosaurus sp. [5]. Единственный вид рода Arstanosaurus, A. akkurganensis Shilin et Suslov, 1982, основан на фрагменте верхней челюсти из сантон-кампанской бостобинской свиты местонахождения Аккурган в Казахстане и в настоящее время считается nomen dubium [6, 7]. Череп экз. ПИН, № 3458/5 идентифицировался как Lambeosaurinae indet. [6: рис. 7B] или Hadrosauridae indet. [8: рис. 21.11]. В.Р. Алифановым [9] этот экземпляр был определен как Hadrosaurinae indet. В настоящей статье мы даем предварительное описание экз. ПИН, № 3458/5 и обсуждаем его возможную таксономическую принадлежность.
Рис. 1.
Hadrosauroidea indet., экз. ПИН, № 3458/5, череп: а – с дорсальной стороны; б – с левой латеральной стороны; местонахождение Байшин-Цав, пустыня Гоби, Монголия; баинширэинская свита, верхний мел (сеноман – сантон).

ОПИСАНИЕ
Экземпляр представляет собой ювенильную особь, и признаки, указывающие на его молодой возраст, отмечены в скобках знаком астериска (*). Длина черепа – 143 мм, высота квадратной кости – 68 мм. Предглазничная часть черепа составляет 42% от общей длины черепа (*). Глазница большая, 29% от общей длины черепа (*). Глазница овальная, дорсовентральный диаметр немного больше переднезаднего диаметра. Нижневисочное окно небольшое (*), каплевидное, заостренное дорсально. Череп наиболее расширен поперечно позади глазниц, поперечная ширина черепа на уровне головок квадратных костей меньше (*). Задний край черепа прямой, медиальные крылья чешуйчатых костей направлены медиально (*). Крыша черепа куполообразная, с дорсально выпуклыми лобными костями (*). Между парными носовыми и лобными костями имеется фонтанель (*). Наружная ноздря небольшая, ее дорсальный край образован носовой костью. Околоноздревой впадины нет. Большое затылочное отверстие ромбовидное, обращено постеровентрально.
Вдоль ротового края предчелюстных костей расположены четыре зубца, уменьшающиеся в размерах от наиболее медиального к наиболее латеральному. Имеется второй, внутренний ряд предчелюстных зубцов, отделенный от внешнего ряда бороздой, содержащей сосудистые отверстия (“двуслойное” строение предчелюстной кости). Дорсальный отросток предчелюстной кости относительно длинный, простирается назад далеко за пределы наружной ноздри (*). Вентролатеральный отросток предчелюстной кости контактирует со слезной костью. Контакт между вентролатеральным отростком предчелюстной кости и предлобной костью отсутствует, по-видимому, из-за неполной сохранности или незавершенного окостенения этих костей.
Самая широкая часть носовых костей находится у заднего конца вентролатерального отростка предчелюстной кости. Задние части парных носовых костей разделены фонтанелью (*) и не перекрывают лобные кости (*).
На верхнечелюстной кости имеется крупный ростролатеральный отросток, отделенный от вентролатерального отростка предчелюстной кости глубоким желобком. Контакт между верхнечелюстной и слезной костями в виде короткой линии между вентролатеральным отростком предчелюстной кости и ростральным отростком скуловой кости. Постеролатеральный край верхнечелюстной кости вырезан глубокой фасеткой для рострального отростка скуловой кости. Вентральная часть верхнечелюстной кости углублена медиально относительно дорсальной части. Между этими двумя частями верхнечелюстной кости протягивается ряд крупных сосудистых отверстий, лучше сохранившийся с левой стороны черепа. Точное число локусов верхнечелюстных зубов нельзя установить, поскольку задняя часть верхнечелюстной кости закрыта зубной костью; присутствует не менее 13 локусов зубов на верхнечелюстной кости (*).
Скуловая кость трехлучевая, с ростральным, заглазничным и каудальным отростками. Ростральный отросток, контактирующий с верхнечелюстной костью, спереди от глазницы немного расширен дорсовентрально. Заглазничный отросток скуловой кости направлен постеродорсально, относительно короткий (*); область контакта этого отростка со скуловым отростком заглазничной кости составляет менее трети его длины (*). Каудальный отросток скуловой кости целиком формирует вентральное обрамление нижневисочного окна. Вентральный край каудального отростка треугольной формы, заострен вентрально. Задняя часть каудального отростка сочленяется с квадратно-скуловой костью.
Слезная кость формирует часть переднего края глазницы и контактирует с заглазничной, носовой, предчелюстной, верхнечелюстной и скуловой костями на латеральных сторонах черепа. Вдоль глазничного края слезной кости протягивается желобок.
Предлобная кость образует антеродорсальную часть глазницы и контактирует с лобной, носовой и слезной костями. Она состоит из двух частей: вертикальной передней и слегка выпуклой горизонтальной задней. Вдоль глазничного края предлобной кости имеется борозда для крепления свободно сочленяющейся надглазничной кости. С левой стороны сохранилась стержневидная надглазничная кость.
Длина лобной кости приблизительно вдвое больше ее ширины (*), дорсальная сторона кости выпуклая (*). Передние концы парных лобных костей разделены фонтанелью. Лобная кость контактирует с носовой и предлобной костями спереди и с заглазничной, боковой клиновидной и теменной костями сзади и имеет относительно длинный свободный глазничный край. Углубленная ямка для предлобной кости небольшая (*). Задние части парных лобных костей разделены треугольным ростральным отростком теменной кости.
Теменная кость образует передний и медиальный края верхневисочного окна, контактирует с заглазничной, лобной и чешуйчатой костями дорсально и с мозговой коробкой вентрально. Теменная кость имеет заметный ростральный отросток, вклинивающийся между задними частями парных лобных костей. Медиальные края полностью сросшихся парных теменных костей образуют отчетливый сагиттальный гребень; он контактирует с чешуйчатыми костями сзади, а спереди раздваивается и протягивается вдоль области контакта теменных и лобных костей к заглазничным костям.
Заглазничная кость трехлучевая, состоит из медиального, вентрального и каудального отростков. Она образует постеродорсальный край глазницы, передний край верхневисочного окна и антеродорсальный край нижневисочного окна. Медиальный отросток массивный, сочленяется с лобной, боковой клиновидной и теменной костями. Треугольный вентральный отросток заострен вентрально и контактирует с заглазничным отростком скуловой кости. Каудальный отросток латерально перекрывает краниальный отросток чешуйчатой кости, формируя верхневисочную перемычку.
Чешуйчатая кость образует задний и латеральный края верхневисочного окна. Парные чешуйчатые кости медиально не контактируют. Краниальный отросток, контактирующий с заглазничной костью, представляет собой тонкую пластинку с острым дорсальным краем. На вентральной стороне чешуйчатой кости имеется глубокая впадина для головки квадратной кости, ограниченная спереди и сзади выступающими преквадратным и постквадратным отростками соответственно. Постквадратный отросток перекрывает переднюю сторону парокципитального отростка боковой затылочной кости.
Квадратная кость дугообразно изогнута вперед. С задней стороны квадратная кость почти вертикальная, расширена вентрально. Вентральный конец квадратной кости двухмыщелковый, с сильнее выступающим дистально латеральным надугловым мыщелком и меньшим медиальным мыщелком, сочленяющимся с суставной впадиной сочленовной кости. Птеригоидное крыло квадратной кости контактирует с крыловидной костью медиально.
Верхнезатылочная кость исключена из дорсального края большого затылочного отверстия вследствие контакта боковых затылочных костей, которые образуют выступающий горизонтальный гребень над большим затылочным отверстием. Верхнезатылочная кость наклонена вперед и несет срединный гребень. Боковые затылычные кости образуют затылочные кондилиды, но медиально не контактируют вентральнее большого затылочного отверстия (*). На дне большого затылочного отверстия имеется узкая полоса основной затылочной кости. Затылочный мыщелок направлен постеровентрально.
Предзубная кость U-образной формы, с прямым передним краем. Ротовой край зубчатый, но зубцы менее отчетливые по сравнению с предчелюстной костью. Дорсальный и вентральный края зубной ветви почти параллельны. Вентральный край спереди вогнутый, а сзади выпуклый. На зубной кости насчитывается не менее 16 зубных локусов (*). Венечный отросток почти вертикальный и лишь немного расширен в дорсальной части. Его задний край образован надугловой костью. Нижний зубной ряд сзади достигает середины венечного отростка. Небольшая часть угловой кости экспонирована латерально на правой стороне черепа.
Как и на верхнечелюстных костях, на зубных костях в зубных рядах имеется не более двух замещающих зубов в каждом зубном локусе (*). Самые крупные зубы находятся ближе к заднему концу зубного ряда. Краевые зубчики редуцированы до состояния мелких бугорков. На коронках зубов нет дополнительных гребней. Главный гребень отчетливый, расположен в центре коронки.
Имеется семь крестцовых позвонков. Лопасть лопатки сильно расширена дистально. Передние конечности более чем в два раза короче задних. Плечевая кость короче лопатки. Когтевые фаланги II и III пальцев кисти уплощенные, копытообразные, дистально заостренные. В V пальце кисти две фаланги. Имеется четкий надвертлужный отросток подвздошной кости, протягивающийся вниз по меньшей мере на половину высоты подвздошной кости. Ствол седалищной кости выпуклый вентрально. Предлобковый отросток лобковой кости расширен дорсовентрально, тонкий в поперечном направлении. Межмыщелковая разгибательная борозда бедренной кости спереди почти закрыта дистальными мыщелками.
ОБСУЖДЕНИЕ
Х. Цогтбаатар и соавт. [10] описали нового гадрозавроидного динозавра Gobihadros mongoliensis Tsogtbaatar et al., 2019 на основе почти полного сочлененного скелета из местонахождения Байшин-Цав и дополнительных образцов из этого и других местонахождений баинширэинской свиты. Новый таксон отличался от всех других гадрозавроидов, не относящихся к гадрозавридам, по предчелюстной кости с “двуслойным” ротовым краем и наличием до трех функциональных зубов в зубном ряду зубной кости. Эти признаки, свойственные Hadrosauridae, считались результатом параллельной эволюции Gobihadros. Другими диагностическими признаками Gobihadros являются шиповидный I палец кисти, сигмовидный дорсальный контур подвздошной кости и сильное латеральное выступание надвертлужного гребня [10]. Три из этих диагностических признаков также имеются у экз. ПИН, № 3458/5: “двуслойный” ротовой край предчелюстной кости, сигмовидный дорсальный контур подвздошной кости и сильное латеральное выступание надвертлужного гребня. I палец кисти не сохранился на экз. ПИН, № 3458/5. В зубном ряду зубной кости у последнего не более двух функциональных зубов, что, вероятно, связано с его ювенильным возрастом.
Цогтбаатар и соавт. [10] провели филогенетический анализ, основанный на таксон-признаковой матрице, сфокусированной на базальных гадрозавроидах и содержащей 108 признаков 36 таксонов. Экз. ПИН, № 3458/5 по перекрывающимся признакам из этой матрицы идентичен Gobihadros во всех случаях, кроме четырех. Одно из этих различий, очевидно, связано с молодым онтогенетическим возрастом экз. ПИН, № 3458/5: предглазничная длина черепа составляет приблизительно 50% от общей длины черепа (1(0)). Три других отличительных признака характеризуют зубную морфологию, которая у экз. ПИН, № 3458/5 имеет более продвинутый облик по сравнению с Gobihadros: краевые зубчики на зубах отсутствуют или редуцированы до небольших бугорков (62(2)); коронки зубов зубной кости симметричные, вершина и главный гребень расположены на средней линии (68(1)); дополнительный гребень мезиальнее главного гребня коронок зубов зубной кости отсутствует или встречается редко (69(2)). Последний признак Цогтбаатаром и соавт. [10] считался синапоморфией Hadrosauridae. При филогенетическом анализе мы следовали аналитическому протоколу Цогтбаатара и соавт. [10]: матрица была проанализирована с использованием алгоритма Branch-and-Bound в PAUP v4.0127, с 14 признаками, обозначенными как ордированные, и Hypsilophodon foxii Huxley, 1869 в качестве внешней группы. В результате анализа было получено 304 наиболее парсимоничных дерева, каждое из которых имеет длину 232 шага, индекс консистентности составляет 0.57, а остаточный индекс – 0.84. На строго консенсусном дереве экз. ПИН, № 3458/5 и Gobihadros попадают в политомию с некоторыми другими базальными таксонами гадрозавроидов. На 68% наиболее парсимоничных деревьев экз. ПИН, № 3458/5 группируется с Gobihadros (рис. 2).
Рис. 2.
Фрагмент консенсусного 50% филогенетического древа, полученного по принципу большинства из 304 наиболее парсимоничных деревьев, восстановленных с использованием алгоритма PAUP Branch-and-Bound, демонстрирующий положение экз. ПИН, № 3458/5. Числа в узлах показывают частоту клад в наиболее парсимоничных деревьях.
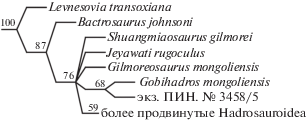
У экз. ПИН, № 3458/5 высота квадратной кости составляет 68 мм, что заметно меньше, чем у полувзрослого голотипа Gobihadros mongoliensis (115 мм) и отнесенного к этому виду взрослого экземпляра (154 мм) [10]. Однако имеется по меньшей мере один черепной признак, который не может быть объяснен онтогенетической изменчивостью: треугольный ростральный отросток теменных костей вклинивается между лобными костями у экз. ПИН, № 3458/5 (рис. 1), тогда как у Gobihadros лобно-теменный шов прямой. Сходный треугольный выступ теменных костей обнаружен у Levnesovia и Batyrosaurus [11, 12]. Другим предполагаемым морфологическим отличием является заглазничный отросток скуловой кости – очень длинный, формирующий длинную область контакта с заглазничной костью у полувзрослого голотипа Gobihadros mongoliensis, но намного более короткий у экз. ПИН, № 3458/5. Это различие, по-видимому, выходит за пределы онтогенетической изменчивости. У экз. ПИН, № 3458/5 подвздошная кость сильно выпуклая дорсально над лобковой ножкой, как у Gilmoreosaurus [13, 14], тогда как у Gobihadros она в этой области почти прямая.
Как отметили Цогтбаатар и соавт. [10], в маастрихте пустыни Гоби Монголии представлены только гадрозавриды Saurolophus и Barsboldia. В формации Ирэн-Дабасу близлежащей области пустыни Гоби во Внутренней Монголии, Китай, присутствуют два таксона негадрозавридных гадрозавроидов – Bactrosaurus и немного более продвинутый Gilmoreosaurus, сходный по эволюционному уровню с Gobihadros [13–15]. Если соотносить формацию Ирэн-Дабасу в Китае с маастрихтской немэгэтинской свитой в Монголии, картина эволюции орнитопод выглядела бы очень странно: продвинутые гадрозавриды в одной части пустыни Гоби и реликтовые негадрозавридные гадрозавроиды в соседней части пустыни Гоби, жившие в одинаковых экологических условиях. Более вероятно, что формация Ирэн-Дабасу древнее и коррелируется с баинширэинской свитой Монголии [16]. Как и в формации Ирэн-Дабасу, в баинширэинской свите могут присутствовать два таксона гадрозавроидов – Gobihadros и несколько более продвинутый таксон, представленный экз. ПИН, № 3458/5.
Список литературы
Цыбин Ю.И., Курзанов С.М. // Труды Совместной советско-монгольской палеонтологической экспедиции. 1979. Т. 8. С. 108–112.
Glut D.F. The New Dinosaur Dictionary. Secaucus, New Jersey: Citadel Press, 1982.
Lambert D. A Field Guide to Dinosaurs. New York: Avon Books, 1983.
Saito T. Wonder of the World’s Dinosaurs. Tokyo: Kodansha Publishers, 1979.
Tsogtbaatar K., Chinzorig T. // Korean Journal of Quaternary Research. 2007. V. 21. № 2. P. 49–53.
Norman D.B., Kurzanov S.M. // Proc. Geologists’ Association. 1997. V. 108. P. 191–199.
Шилин П.В., Суслов Ю.В. // Палеонтологический журнал. 1982. № 1. С. 131–135.
Norman D.B., Sues H.-D. // The Age of Dinosaurs in Russia and Mongolia. Benton M.J., Shishkin M.A., Unwin D.M., Kurochkin E.N., eds. Cambridge: Cambridge University Press, 2000. P. 462–479.
Алифанов В.Р. // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 2. Курочкин Е.Н., Лопатин А.В., ред. М.: ГЕОС, 2012. С. 153–309.
Tsogtbaatar K., Weishampel D.B., Evans D.C., Watabe M. // PLoS One. 2019. V. 14. № 4: e0208480.
Sues H.-D., Averianov A.O. // Proc. Royal Society B: Biological Sciences. 2009. V. 276. № 1667. P. 2549–2555.
Godefroit P., Escuillié F., Bolotsky Y.L., Lauters P. // Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Godefroit P., ed. Bloomington, Indianopolis: Indiana University Press, 2012. P. 335–358.
Gilmore C.W. // Bulletin of the American Museum of Natural History. 1933. V. 67. P. 23–78.
Prieto-Márquez A., Norell M.A. // American Museum Novitates. 2010. № 3694. P. 1–49.
Godefroit P., Dong Z.-M., Bultynck P., et al. // Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre. 1998. V. 68. Suppl. P. 3–70.
Averianov A.O., Sues H.-D. // Journal of Stratigraphy. 2012. V. 36. № 2. P. 462–485.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле