

Доклады Российской академии наук. Науки о Земле, 2022, T. 506, № 2, стр. 210-213
Первая находка хищных динозавров семейства Avimimidae в позднем мелу Центральной Азии
А. О. Аверьянов 1, *, академик РАН А. В. Лопатин 2
1 Зоологический институт Российской академии наук
Санкт-Петербург, Россия
2 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: dzharakuduk@mail.ru
Поступила в редакцию 16.05.2022
После доработки 25.05.2022
Принята к публикации 26.05.2022
- EDN: MMYRGX
- DOI: 10.31857/S2686739722600692
Аннотация
Дистальный фрагмент тибиотарзуса из позднемелового местонахождения Кансай (яловачская свита, сантон) в Таджикистане отнесен к овирапторозавровым тероподам семейства Avimimidae по срастанию большой берцовой кости и проксимальных тарзальных костей в тибиотарзус, редукции малой берцовой кости и срастанию ее дистального конца с латеральным мыщелком тибиотарзуса, а также по наличию хорошо развитого постфибулярного фланга. Описанный экземпляр практически идентичен тибиотарзусу Avimimus portentosus Kurzanov, 1981 из позднего мела Монголии и может быть определен как Avimimus sp. Он является одной из древнейших находок семейства Avimimidae в ископаемой летописи, примерно одновозрастной с авимимидом из формации Ирэн-Дабасу в Северном Китае.
Авимимиды – специализированные птицеподобные хищные динозавры из клады Oviraptorosauria [1]. Авимимиды отличаются от других овирапторозавров адаптацией к быстрому бегу, выраженной в удлинении голени относительно бедра, образовании тибиотарзуса путем полного срастания большой берцовой кости и проксимальных тарзальных костей, редукции малой берцовой кости, арктометатарзальном строении стопы и срастании боковых плюсневых костей в проксимальной части [2, 3]. До сих пор авимимиды были известны только из верхнемеловых отложений пустыни Гоби, в Монголии и провинции Внутренняя Монголия, Китай [1, 2, 4–7]. В данной работе приводится описание первой находки авимимида за пределами пустыни Гоби – дистального фрагмента тибиотарзуса из верхнемеловых отложений Таджикистана (Центральная, или Средняя, Азия). Находка происходит из яловачской свиты (сантон [8]) местонахождения Кансай, откуда известен разнообразный комплекс динозавров, включающий завропод, хищных динозавров Ornithomimidae, Therizinosauridae, Troodontidae, дромеозаврида Kansaignathus sogdianus Averianov et Lopatin, 2021 и гадрозавроидов [8–13]. Описываемый в статье фрагмент тибиотарзуса авимимида был найден в ходе раскопок на местонахождении Кансай (40°29′ с.ш., 69°42′ в.д., около 22 км на север от г. Худжанд, Согдийская область) под руководством А.К. Рождественского [14] в 1963–1964 гг. и хранится в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН, коллекция № 3041), г. Москва.
На экз. ПИН, № 3041/49 проксимальные тарзалии полностью срастаются между собой и с большой берцовой костью, образуя тибиотарзус. Отчетливая борозда вдоль медиального края восходящего отростка таранной кости представляет собой почти полностью закрывшийся шов между таранной и большой берцовой костями (рис. 1 б, в). Диафиз тибиотарзуса уплощен в переднезаднем направлении, его ширина в 1.6 раза превышает переднезадний диаметр. Сечение диафиза близ сохранившегося проксимального конца овальной формы (рис. 1 а). Передняя и задняя поверхности диафиза плоские. Диафиз резко расширяется в дистальном направлении. Наибольшая ширина дистального мыщелка в 1.8 раза превышает ширину диафиза близ сохранившегося проксимального конца. Дистальный мыщелок загнут вперед. Медиальный и латеральный мыщелки примерно равны по размерам при виде спереди. При виде с дистального конца медиальный мыщелок больше выдается вперед и медиально (рис. 1 г). Вдоль проксимального края мыщелков на передней стороне восходящего отростка таранной кости имеется веретеновидное углубление (рис. 1 в). Постфибулярный фланг латерального мыщелка в виде мощного гребня, поднимающегося вдоль заднелатерального края диафиза (рис. 1 д, е). Небольшой дистальный фрагмент малой берцовой кости сохранился в сочленении с латеральным мыщелком, образованным пяточной костью (рис. 1 в, д, е). Он сращен с тибиотарзусом, но швы между двумя костями хорошо видны спереди и с латеральной стороны (рис. 1 в, д). Проксимальнее сохранившегося дистального фрагмента имеется узкая продольная бороздка, в которой помещалась несросшаяся часть малой берцовой кости (рис. 1 д). На латеральном мыщелке дистальнее конца малой берцовой кости имеется короткая, но глубокая продольная борозда (рис. 1 д). На медиальном мыщелке имеется короткий поперечный гребень (рис. 1 б).
Рис. 1.
Avimimus sp., экз. ПИН, № 3041/49, дистальный фрагмент левого тибиотарзуса: (а) с проксимального конца; (б) с медиальной стороны; (в) спереди; (г) с дистального конца; (д) с латеральной стороны; (е) сзади. Фотографии (а–е) и скриншоты цифровой модели (а'–е'). Таджикистан, Согдийская область, местонахождение Кансай; верхний мел (сантон), яловачская свита. Обозначения: d – веретеновидная депрессия на передней стороне тибиотарзуса; fgr – борозда для малой берцовой кости; fi – малая берцовая кость (обозначена голубым цветом); gr – борозда на латеральном мыщелке; lc – латеральный мыщелок; mc – медиальный мыщелок; pff – постфибулярный фланг; r – гребень на медиальном мыщелке; s – шов между восходящим отростком таранной кости и большой берцовой костью.
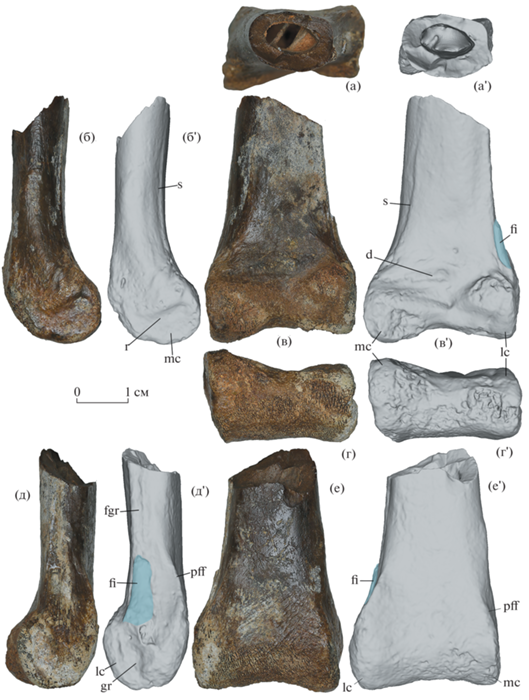
Ширина дистального эпифиза – 30.5 мм; переднезадний диаметр дистального эпифиза – 18.6 мм.
Экз. ПИН, № 3041/49 отнесен к Avimimus Kurzanov, 1981, типовому роду монотипичного семейства Avimimidae Kurzanov, 1981, на основании следующих признаков: дистальные тарзалии срастаются между собой и с большой берцовой костью, образуя тибиотарзус; малая берцовая кость редуцирована и ее дистальный конец срастается с латеральным мыщелком тибиотарзуса видимыми швами; более проксимальная часть малой берцовой кости помещается в узкой борозде вдоль переднелатерального края тибиотарзуса; на тибиотарзусе имеется хорошо развитый постфибулярный фланг [1, 2, 4, 7]. Непосредственное сравнение экз. ПИН, № 3041/49 с тибиотарзусом голотипа Avimimus portentosus Kurzanov, 1981 (экз. ПИН, № 3907/1) из местонахождения Удан-Сайр в Монголии показало их практически полную идентичность по строению и размерам. Кансайский экземпляр отличается присутствием веретеновидной депрессии на передней стороне тибиотарзуса проксимальнее дистальных мыщелков и более развитыми постфибулярным флангом и поперечным гребнем на медиальном мыщелке. Эти отличия могут быть связаны с большим онтогенетическим возрастом кансайского экземпляра. Такое явное сходство позволяет определить экз. ПИН, № 3041/49 как Avimimus sp.
Находка Avimimus sp. в сантоне Таджикистана является одной из древнейших для семейства Avimimidae. Из близкой по возрасту формации Ирэн-Дабасу (турон–сантон) в провинции Внутренняя Монголия, Китай (о возрасте формации см. [15]), известны многочисленные остатки авимимид, определенные как Avimimidae indet. [4]. Геологический возраст Avimimus portentosus проблематичен. При первоописании типовым местонахождением вида было указано местонахождение Удан-Сайр, которое традиционно относится к кампанской джадохтской свите [2, 16]. По другим данным, голотип может происходить из близлежащего местонахождения Ягаан-Ховил, которое относится к маастрихтской нэмэгэтинской свите [17]. Однако из отложений нэмэгэтинской свиты (местонахождение Нэмэгт) недавно был описан особый вид авимима – A. nemegtensis Funston et al., 2018 [7].
Представители семейства Avimimidae появляются в геологической летописи уже в полностью сформированном виде, с характерными адаптациями к быстрому бегу, и на протяжении примерно 15 миллионов лет последующей эволюции не претерпевают существенных изменений. В настоящее время неизвестны какие-либо овирапторозавры, которые могли бы рассматриваться как предки авимимид, поэтому происхождение семейства Avimimidae остается загадкой.
Список литературы
Osmólska H., Currie P.J., Barsbold R. Oviraptorosauria // The Dinosauria. 2nd ed. Berkeley, Los Angeles, London: Univ. California Press, 2004. P. 165–183.
Курзанов С.М. Авимимиды и проблема происхождения птиц // Труды Совм. Сов.-Монгол. палеонтол. эксп. 1987. Т. 31. С. 1–93.
Holtz T.R., Jr. The arctometatarsalian pes, an unusual structure of the metatarsus of Cretaceous Theropoda (Dinosauria: Saurischia) // J. Vertebr. Paleontol. 1994. V. 14. № 4. P. 480–519.
Funston G.F., Currie P.J., Ryan M.J., Dong Z.-M. Birdlike growth and mixed-age flocks in avimimids (Theropoda, Oviraptorosauria) // Sci. Rep. 2019. V. 9. P. 18816.
Tsuihiji T., Witmer L.M., Watabe M., et al. New information on the cranial morphology of Avimimus (Theropoda: Oviraptorosauria) // J. Vertebr. Paleontol. 2017. V. 37. № 4. P. e1347177.
Funston G.F., Currie P.J., Eberth D.A., et al. The first oviraptorosaur (Dinosauria: Theropoda) bonebed: evidence of gregarious behavior in a maniraptoran theropod // Sci. Rep. 2016. V. 6. P. 35782.
Funston G.F., Mendonca S.E., Currie P.J., Barsbold R. Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2018. V. 494. P. 101–120.
Averianov A.O., Sues H.-D. Review of Cretaceous sauropod dinosaurs from Central Asia // Cret. Res. 2017. V. 69. P. 184–197.
Алифанов В.Р., Аверьянов А.О. О находках орнитомимидных динозавров (Saurischia, Ornithomimosauria) в отложениях верхнего мела Таджикистана // Палеонтол. журн. 2006. № 1. С. 98–102.
Аверьянов А.О., Лопатин А.В. Теризинозавр (Theropoda, Therizinosauridae) из позднего мела Таджикистана // Доклады РАН. Науки о Земле. 2022. Т. 504. № 1. С. 65–68.
Averianov A.O., Sues H.-D. A new troodontid (Dinosauria: Theropoda) from the Cenomanian of Uzbekistan, with a review of troodontid records from the territories of the former Soviet Union // J. Vertebr. Paleontol. 2007. V. 27. № 1. P. 87–98.
Аверьянов А.О., Лопатин А.В. Новый хищный динозавр (Theropoda, Dromaeosauridae) из позднего мела Таджикистана // Доклады РАН. Науки о Земле. 2021. Т. 499. № 1. С. 49–53.
Аверьянов А.О., Алифанов В.Р. Новое в изучении остатков утконосых динозавров (Ornithischia, Hadrosauridae) из верхнемеловых отложений Таджикистана // Палеонтол. журн. 2012. № 5. С. 67–73.
Рождественский А.К. Кансайское местонахождение меловых позвоночных в Фергане // Ежегодн. Всес. палеонтол. об-ва. 1977. Т. 30. С. 235–247.
Averianov A.O., Sues H.-D. Correlation of Late Cretaceous continental vertebrate assemblages in Middle and Central Asia // J. Stratigr. 2012. V. 36. № 2. P. 462–485.
Czepiński Ł. New protoceratopsid specimens improve the age correlation of the Upper Cretaceous Gobi Desert strata // Acta Palaeontol. Polon. 2020. V. 65. № 3. P. 481–497.
Watabe M., Tsogtbaatar K., Suzuki S., Saneyoshi M. Geology of dinosaur-fossil-bearing localities (Jurassic and Cretaceous: Mesozoic) in the Gobi Desert: results of the HMNS-MPC Joint Paleontological Expedition // Hayashibara Mus. Nat. Sci. Res. Bull. 2010. V. 3. P. 41–118.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле