

Доклады Российской академии наук. Науки о Земле, 2022, T. 507, № 1, стр. 75-79
Новый вид тюленей рода Pachyphoca (Cystophorinae, Phocidae) из позднего миоцена Северного Кавказа
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: tarasenkokk@gmail.com
Поступила в редакцию 19.07.2022
После доработки 22.07.2022
Принята к публикации 24.07.2022
- EDN: XYYUBP
- DOI: 10.31857/S2686739722601417
Аннотация
По остаткам тюленя из верхнемиоценовых отложений местонахождения Фортепьянка в Республике Адыгея описан новый вид – Pachyphoca volkodavi. Это первые находки Pachyphoca на Северо-Западном Кавказе, расширяющие представления о палеогеографическом и стратиграфическом распространении этого рода.
Подсемейство Cystophorinae в настоящее время представлено тюленем хохлачом Cystophora cristata Erxleben, 1777. В позднем миоцене (среднем сармате) Северного Причерноморья были широко распространены представители рода Pachyphoca: P. ukrainica Koretsky et Rahmat, 2013 и P. chapskii Koretsky et Rahmat, 2013 [1, 2].
В 2019 г. собраны ископаемые остатки тюленей в местонахождении Фортепьянка 2, которое было открыто в 2003 г. в долине р. Фортепьянки (притоке р. Белой) на Северном Кавказе (рис. 1 а) [3]. Эти остатки (плечевая кость, ребра и грудные позвонки) найдены в небольшой песчаной линзе нижней части разреза (слой 2, см. рис. 1 б), которая относится к первой половине тортона Восточного Паратетиса и соответствует раннему валлезию (зона MN 9 Европы) [4]. Другая часть остатков этой же формы найдена в залегающем выше песчаном слое 9. Из верхнесарматской части этого разреза (слои 23, 26) известны тюлени Monochopsis pontica (Eichwald, 1850) [3].
Рис. 1.
Местонахождение Фортепьянка 2 (верхний миоцен, средний сармат): а – географическое положение; б – сводный геологический разрез на р. Фортепьянке с указанием находок тюленей. 1 – галечники и гравийники; 2 – глинистые пески; 3 – пески; 4 – песчаники; 5 – глинистые пески; 6 – мергели; 7 – алевриты; 8 – известняки; 9 – биогенные известняки; раковины моллюсков; 11 – границы между подразделениями; 12 – находки позвоночных.

Изучение новых материалов позволило отнести их к новому виду рода Pachyphoca. Изученные образцы хранятся в коллекции Палеонтологического института им. А.А. Борисяка (ПИН), г. Москва.
ПОДСЕМЕЙСТВО CYSTOPHORINAE GRAY, 1886
Pachyphoca volkodavi Tarasenko, sp. nov.
Название вида в честь геолога И.Г. Волкодава.
Голотип – ПИН № 3462/46; правая плечевая кость; Россия, Республика Адыгея, Майкопский р-н, Фортепьянка 2; верхний миоцен, MN 9; средний сармат, блиновская свита.
Описание (рис. 2, 3). Тюлень средних размеров с выраженным пахиостеосклерозом костей. Плечевая кость массивная. Межбугорковая борозда широкая и неглубокая. Дельтопекторальный гребень короткий, расширен в средней части. Дельтовидная бугристость широкая, вытянутой субтреугольной формы (рис. 2 а, 2 в). Поверхность прикрепления дельтовидной мышцы широкая, направлена кзади. Головка плечевой кости выпуклая и полусферическая, слегка сжата краниокаудально (рис. 2 б); расположена немного ниже уровня большого и малого бугорков. Малый бугорок плечевой кости, крупный, округлый и расположен дистальнее по отношению к головке плечевой кости и большому бугорку (рис. 2 г). Ямка для медиальной головки трехглавой мышцы плеча хорошо выражена. Ямка для грудной мышцы не выражена.
Рис. 2.
Pachyphoca volkodavi Tarasenko, sp. nov., голотип ПИН, № 3462/46, правая плечевая кость: а – с краниальной стороны, б – с каудальной стороны, в – с латеральной стороны, г – с медиальной стороны; р. Фортепьянка, Северный Кавказ; верхний миоцен, средний сармат.
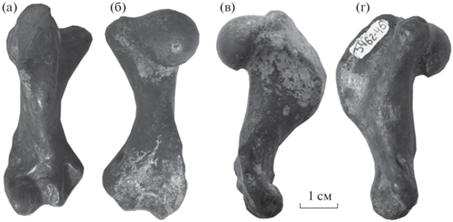
Рис. 3.
Pachyphoca volkodavi Tarasenko, sp. nov.: а – экз. ПИН, № 3462/82, грудной позвонок, с каудальной стороны, б–г – экз. ПИН, № 3462/83, поясничный позвонок: б – с краниальной стороны, в – с каудальной стороны, г – с правой латеральной стороны; Фортепьянка, Северный Кавказ; верхний миоцен, средний сармат.

Тело грудного позвонка укороченное (рис. 3 а). Тело поясничного позвонка (рис. 3 б–3 г) уплощено в дорсовентральном направлении и имеет в поперечном сечении сердцеобразную форму, позвоночное отверстие овальной формы. Остистые отростки очень короткие и широкие. Поперечные отростки короткие, утолщенные и направлены краниовентрально. Каудальный суставной отросток очень короткий и округлый. Сосцевидный отросток толстый, широкий и короткий. Невральная арка очень массивная, утолщенная.
Ребро с ярко выраженным пахиостеосклерозом, массивное и утолщенное. Структура губчатой оболочки сохранилась, хотя межтрабекулярные пространства заполнены эндостальными отложениями (рис. 4).
Рис. 4.
Поперечный срез через ребро с выраженным пахиостеосклерозом, экз. ПИН, № 3462/85; Фортепьянка, Северный Кавказ; верхний миоцен, средний сармат.

Размеры в мм. Голотип ПИН, № 3462/46, плечевая кость: общая длина кости – 87.3; длина дельтопекторального гребня – 46.7; ширина дельтопекторального гребня – 18; высота головки – 21.9; ширина головки – 25.2; ширина проксимального эпифиза – 37.1; толщина проксимального эпифиза – 47.6; поперечная ширина диафиза – 16.6; диаметр диафиза – 27.8; ширина межбугорковой борозды (максимальная) – 13.3; высота блока – 17.0; ширина блока (дистальная) – 19.7; ширина дистального эпифиза – 34.5; ширина медиального мыщелка – 16.9; ширина блока (спереди) – 14.6.
Экз. ПИН, №3462/83, фрагмент плечевой кости: длина дельтопекторального гребня – ~45.1; высота головки – 20; ширина головки – 23.5; ширина проксимального эпифиза – 35.2; поперечная ширина диафиза – 15.7.
Экз. ПИН, № 3462/82, грудной позвонок: высота тела (спереди) – 17.2; ширина тела (спереди) – 19.5; длина тела – 20.6; высота тела (сзади) – 19.3; ширина тела (сзади) – 24.3.
Экз. ПИН, № 3462/83, поясничный позвонок: высота тела (спереди) – 18.9; ширина тела (спереди) – 20.1; длина тела – 19.2; высота тела (сзади) – 19.5; ширина тела (сзади) – 22.
Сравнение. От P. ukrainica и P. chapskii новый вид отличается размерами и формой дельтопекторального гребня (гребень укорочен, дельтовидная бугристость более широкая); положением и формой малого бугорка плечевой кости (более массивный, расположен под прямым углом к поперечной оси головки плечевой кости, за счет чего межбугорковая борозда более широкая); формой блока плечевой кости (менее широкий). Кроме того, P. volkodavi отличается от P. chapskii более мелкими размерами и строением латерального надмыщелка плечевой кости (он короче и немного выше медиального, схожее состояние у P. ukrainica), а от P. ukrainica – положением венечной ямки (находится примерно на одном уровне с надмыщелковым отверстием).
Замечания. P. volkodavi отличается от P. ukrainica и P. chapskii более коротким дельтопекторальным гребнем. Исследования онтогенетической и половой изменчивости на основе посткраниальных элементов современных и ископаемых тюленей [5–8] показывают, что половой диморфизм в строении плечевой кости выражен в массивности и удлиненности этой кости у самцов. Плечевая кость у самцов Cystophora длиннее и чуть толще, чем у самок, головка плечевой кости сжата в дорсовентральном направлении, а у самок больше и более сферической формы [1]. При более удлиненной плечевой кости у самцов длина дельтопекторального гребня такая же, как у самок, но дистальная часть дельтопекторального гребня у самок узкая, V-образной формы, у самцов она шире, U-образной формы (в строении плечевой кости современных Phocinae эта разница не наблюдается) [1, 8]. Таким образом, более мелкие размеры дельтопекторального гребня P. volkodavi не связаны с индивидуальной изменчивостью и половым диморфизмом.
Следует отметить, что укороченный дельтопекторальный гребень, массивный и расположенный под прямым углом к поперечной оси головки плечевой кости малый бугорок у P. volkodavi свидетельствуют о специализации плечевого сустава. У многих миоценовых Phocidae Восточного Паратетиса малый бугорок не так сильно отведен от большого и формирует более узкий межбугорковый канал, как, например, у Monochopsis pontica (Eichwald, 1850). У этих тюленей подлопаточная мышца крепится к малому бугорку, который развит сильнее большого бугорка плечевой кости [9, 10]. Сдвиг малого бугорка вперед и его разрастание у P. volkodavi свидетельствуют о мощной подлопаточной мышце, которая как часть ротаторной манжеты плеча играет важную роль в стабилизации плечевого сустава и участвует в пронации и сгибании в суставе (одновременно с участием большой круглой мышцы) во время пропульсивного гребка.
У P. volkodavi, как и у прочих представителей рода Pachyphoca кости сильно утолщены и несут выраженные признаки пахиостеосклероза. В настоящее время отмечено, что развитие пахиостеосклероза костей у морских млекопитающих в Центральном Паратетисе совпадает с условиями гиперсолености (баденский кризис солености) около 13.8–13.4 млн л.н.; впоследствии морские млекопитающие с пахиостеосклерозом распространились в Восточном Паратетисе [11]. Считается, что тюлени с выраженным пахиостеосклерозом костей плавают с небольшой скоростью и ныряют на небольшие глубины, и это предполагает специализацию, связанную с охотой на медленную придонную добычу [1]. Для P. volkodavi основным объектом охоты, возможно, являлись придонные мелководные Sparidae, известные из данного местонахождения [4] и имевшие широкое распространение в Паратетисе в позднем миоцене [12].
Распространение. Верхний миоцен (средний сармат), Северный Кавказ.
Материал. Помимо голотипа, из типового мастонахождения экз. ПИН № 3462/82, грудной позвонок (слой 2); экз. ПИН, № 3462/84, фрагмент плечевой кости (слой 9); экз. ПИН, № 3462/83, фрагмент поясничного позвонка (слой 9); экз. ПИН, № 3462/85, фрагмент ребра (слой 9).
Список литературы
Koretsky I.A., Rahmat S.J. First record of fossil Cystophorinae (Carnivora, Phocidae): Middle Miocene seals from the northern Paratethys // Riv. Ital. Paleontol. Stratigr. 2013. V. 119. № 3. P. 325–350.
Тарасенко К.К., Титов В.В. Первая находка тюленей рода Pachyphoca (Carnivora, Phocidae) в породах миоценового возраста на территории Ростовской области // Доклады РАН. Науки о Земле. 2022. Т. 505. № 2. С. 176–180.
Тарасенко К.К., Лопатин А.В., Зеленков Н.В. и др. Новые местонахождения наземных и морских млекопитающих позднего миоцена в Адыгее (Россия, Северный Кавказ) // ДАН. 2014. Т. 459. № 6. С. 726–728.
Вислобокова И.А., Тарасенко К.К. Находки жвачных (Artiodactyla, Ruminantia) и новые данные по стратиграфии позднемиоценового местонахождения Фортепьянка 2 Северный Кавказ, Республика Адыгея) // Палеонтол. журн. 2019. № 6. С. 105–110.
Гаджиев Д.В. Половой диморфизм в структуре крестцовой кости каспийского тюленя // Фауна экол. назем. водн. живот. Кура-Араксинской низменности малого Кавказа. № 6. 1982. С. 90–95.
Коретская И.А. Половой диморфизм в структуре плечевой и бедренной кости Monachopsis pontica (Pinnipedia:Phocinae) // Вестн. Зоол. № 4. 1987. С. 77–82.
Van Bree P., Erdbrink D. Fossil Phocidae in some Dutch collections (Mammalia, Carnivora) // Beaufortia, Inst. Taxon. Zool. (Zool. Mus.), Univ. Amsterdam, 37. 1987. P. 43–66.
Koretsky I.A. Morphology and Systematics of Miocene Phocinae (Mammalia: Carnivora) from Paratethys and the North Atlantic Region // Geol. Hung. Ser. Paleontol. № 54. 2001. P. 1–109.
Howell A.B. Contribution to the comparative anatomy of the eared and earless seals (genera Zalophus and Phoca). Proc. US National Mus. 1929. V. 73. 600 p.
Howell A.B. Aquatic mammals. Charles C. Thomas, Baltimore, Maryland. 1930. 338 p.
Dewaele L., Gol’din P., Marx F.G., et al. Hypersalinity drives convergent bone mass increases in Miocene marine mammals from the Paratethys // Current biology: CB 32 (1). 2021. https://doi.org/10.1016/j.cub.2021.10.065
Rüeckert-Űlkümen N. Carangidae, Pricanthidae, Scopaenidae und Sparidae (Pisces) aus den sarmatischen Schicten von Pinarhisar (Thrakien, Turkei). Mitteilungen der Bayerischen Staatssammlungfuer Palaeontologie und Historische Geologie 35. 1995. P. 65–86.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле