

Доклады Российской академии наук. Науки о Земле, 2022, T. 507, № 2, стр. 296-302
Корреляционный и палеогеографический потенциал граптолитовых шкал при сопоставлении ордовикских терригенных глубоководных последовательностей циркумполярных областей
1 Институт нефтегазовой геологии и геофизики
им. А.А. Трофимука Сибирского отделения
Российской академии наук
Новосибирск, Россия
* E-mail: sennikovnv@ipgg.sbras.ru
Поступила в редакцию 05.09.2022
После доработки 07.09.2022
Принята к публикации 08.09.2022
- EDN: CQOABQ
- DOI: 10.31857/S2686739722601867
Аннотация
Построена зональная граптолитовая шкала ордовика о. Беннетта. Приведена циркумполярная корреляция терригенных последовательностей ордовика Арктических районов Евразии и Северной Америки. Проанализированы возможные пути миграции пелагических организмов между ордовикскими бассейнами. Показана палеогеографическая близость в ордовике бассейнов Аляски и островов Де Лонга.
Нижние границы ярусов ордовика в современной Общей (и Международной) стратиграфической шкале маркированы первым появлением видов-индексов граптолитовых зон [1, 2]. Корреляция зональных шкал удаленных друг от друга геологических регионов позволяет однозначно судить (с точностью до граптолитовой зоны – менее 1 млн лет), являются ли сходные седиментологические проявления геологических событий в таких регионах одновозрастными и, вследствие этого, предполагать для них возможность единого триггера, либо доказать их разновозрастность и достоверно обосновать последовательность по времени их проявлений.
В Арктическом секторе России и прилегающих районах к хорошо изученным терригенным ордовикским отложениям, содержащим ортостратиграфическую группу – граптолиты, относятся последовательности на Новой Земле, Таймыре и Колыме [2]. Наименее биостратиграфически изученным объектом в Российской Арктике вследствие своей труднодоступности до недавнего времени оставался Архипелаг Новосибирские острова. Крупнейшим островом архипелага является о. Котельный, а на северо-востоке архипелага расположена группа островов Де Лонга – Беннетта, Генриетты, Жаннетты, Жоховский и Вилькицкого (рис. 1).
Рис. 1.
Схема расположения ключевых регионов Евразии и Северной Америки с ордовикcкими граптолит-содержащими терригенными разрезами. Звездочкой отмечен опорный район исследований – о. Бенннета. Толщина стрелок (большая, средняя и малая) соответствует высокому, среднему и низкому потенциалу межбассейновых миграций пелагических организмов. Прерывистой стрелкой показано маловероятное, сверхдальнее направление миграций.
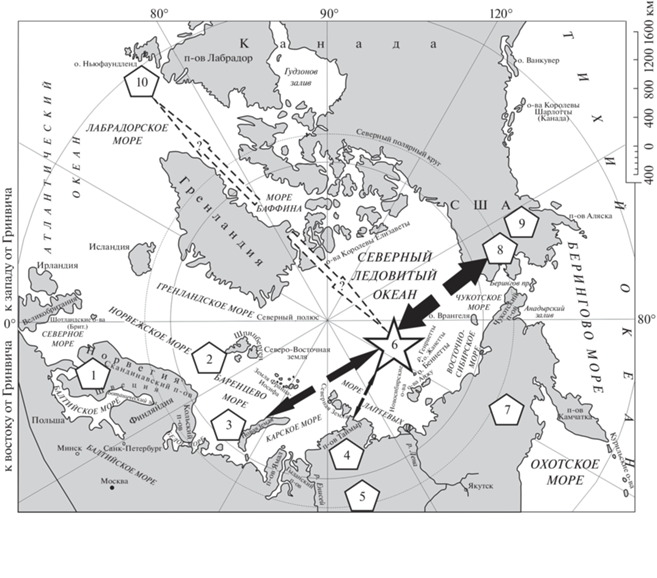
Планомерные геологические исследования этого района Арктики были осуществлены в середине прошлого века, что позволило создать геологическую карту Новосибирских островов масштаба 1 : 1 000 000 [3].
Ордовикские отложения широко распространены на Новосибирских островах (острова Котельный, Беннетта, Жаннетты, Генриетты) (рис. 1). Они сложены двумя типами разрезов [3–5] – карбонатным и терригенным. Карбонатные отложения вскрываются на о. Котельный, где они охватывают весь стратиграфический интервал от флоского до хирнантского века и сложены плитчатыми, реже массивными органогенными известняками. Мощность карбонатного типа ордовика на Новосибирских островах достигает 1350 м. В известняках известны разнообразные кораллы, мшанки, криноидеи, брахиоподы, трилобиты, наутилоидеи, гастроподы, конодонты [3, 4].
Терригенный тип ордовика установлен на островах Беннетта, Жаннетты и Генриетты [3–5] (рис. 1). В его составе присутствуют темно-серые и черные аргиллиты, алевролиты, песчаники, с редкими прослоями известняков. Суммарная мощность терригенного типа разреза более 1100 м. В нем фаунистические остатки (граптолиты и конодонты) известны пока только на о. Беннетта, по которым он датируется ранним и средним ордовиком. На островах Жаннетты и Генриетты предполагается присутствие среднего и верхнего отделов этой системы. Ордовик на о. Беннетта объединяется с верхним кембрием в единую черносланцевую данбарскую свиту [5], делящуюся в ордовикской части на три толщи, мощностью, соответственно, 130–140 м, не менее 250 м и не менее 730 м.
Первое упоминание о находках граптолитов на о. Беннетта в отечественной литературе имеется в работе Б.Н. Аверьянова [6], указавшего, что в коллекции экспедиции Э.В. Толля 1902 г. имеются граптолиты, близкие к колымским граптолитам аренигского (по современной терминологии флоско-дапинского) уровня. Об этой коллекции Э.В. Толля приводит информацию и Р.Ф. Соболевская [7], отмечающая, что эти образцы граптолитов изучал шведский палеонтолог Г.Т. Троедссон (G.T. Troedsson), установивший в них родовые таксоны Didymograptus McCoy, Tetragraptus Salter, Phyllograptus Hall. Позднее, в 1937 г., М.М. Ермолаевым были найдены граптолиты, определенные А.М. Обутом как Didymograptus murchisoni (Beck), что позволило последнему датировать вскрывающиеся там терригенные породы средним ордовиком [8]. В коллекции Д.А. Вольнова и Д.С. Сорокова, собранной в середине прошлого века на о. Беннетта, А.М. Обут определил граптолиты, доказывающие наличие на острове не только среднего, но и нижнего отделов системы: Tetragraptus sp., Phyllograptus sp., Dichograptus sp., Azygograptus sp., Tristichograptus ensiformis (Hall), Cryptograptus antennarius (Hall), Cryptograptus aff. hopkinsoni (Nicholson), Levisograptus cf. dentatus (Brongniart), Didymograptus sp., Didymograptus ex gr. murchisoni (Beck), Glossograptus sp., Leptograptus sp., Diplograptus sp., Gymnograptus sp. (здесь и далее приведена современная родовая и видовая систематика) [3].
Из сборов граптолитов 1972 г. с острова Беннетта Б.А. Клубовым, изучавшая их Р.Ф. Соболевская, пополнила их список – Loganograptus ex gr. logani (Hall), Didymograptus ex gr. bifidus (Hall), Cardiograptus aff. crawfordi Harris, Glossograptus aff. acanthus Elles et Wood, Pseudoclimacograptus sp., Cryptograptus tricornis schaeferi Lapworth, Paraglossograptus aff. etheridgei Harris [7].
В 2011 г. Т.Ю. Толмачева в составе организованной ВСЕГЕИ Международной экспедиции на Новосибирские острова проводила на о. Беннетта биостратиграфические исследования и собрала граптолиты, определенные затем Р.Ф. Соболевской [4]. В нижней части разреза были найдены дапинские граптолиты Pendeograptus pendens (Elles), Isograptus ex gr. сaduceus (Salter), Tylograptus sp., а в верхней части разреза – верхнедапинские и дарривильские таксоны Goniograptus tumidus (Harris et Thomas), Expansograptus sp., Paraglossograptus tentaculatus (Hall), Diplograptus sp., Climacograptus sp., Levisograptus aff. dentatus (Brongniart), Cryptograptus tricornis (Carruthers), Cryptograptus schaeferi Lapworth, Phyllograptus aff. anna Hall, Glossograptus aff. hincksii (Hopkinson), Glossograptus sp., которые, по мнению Р.Ф. Соболевской, указывали на присутствие в разрезе зон hirundo и murchisoni [4].
В 2011 г. М.К. Данукалова и А.Б. Кузьмичев провели детальное полевое геологическое и стратиграфическое изучение ордовика о. Беннетта [5] и собрали коллекцию граптолитов, переданную для изучения автору настоящего сообщения. В работе [5] был представлен первый каркас беннеттской зональной последовательности из шести граптолитовых зон: approximatus, densus, bifidus (= protobifidus), gibberulus, hirundo, austrodentatus. При подготовке к передаче коллекции граптолитов о. Беннетта в Центр “Геохрон” при ИНГГ СО РАН были проведены ревизия определений и сопровождающий ее этап обработки материала, выявивший дополнительные отпечатки граптолитов на различных поверхностях образцов. Это позволило актуализировать биостратиграфические данные и осуществить детальный подход к процедуре обособления зональных комплексов и к выбору их видов-индексов. В результате была дополнена вся граптолитовая последовательность ордовика о. Беннетта. Эта процедура была совершенно необходима, так как в последние годы произошел пересмотр родовой систематики ордовикских граптолитов [9], что повлекло за собой кардинальные изменения как родовых, так и видовых наименований подразделений граптолитовых шкал во всех геологических регионах мира [1, 10]. Сведение в единый реестр всех определений граптолитов из ордовика о. Беннетта позволило добавить в шкалу шесть подразделений: слои с Tetragraptus akzharensis, зону angustifolius elongatus / angustifolius tenuis, слои с Oncograptus, слои с Levisograptus dentatus, слои с Didymograptus artus (=“bifidus”) и слои с Didymograptus murchisoni, а также подзоны deflexus, maximo-divergens и caduceus nanus (рис. 2).
Рис. 2.
Схема корреляции ордовикских граптолитовых подразделений, выделяемых в тонко-терригенных осадочных последовательностях Арктических районов Евразии и Северной Америки.
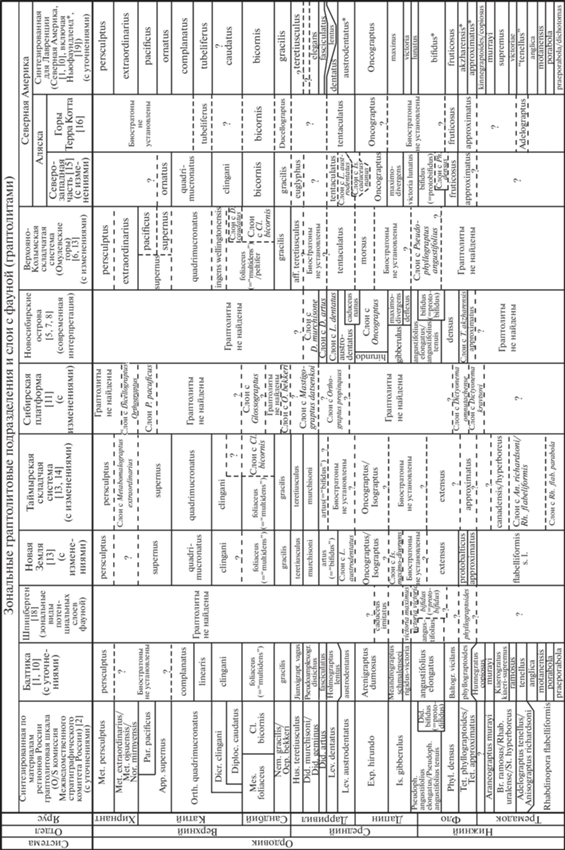
Большинство из рассмотренных граптолитовых зон о. Беннетта относятся к комплексным зонам. По материалам о. Беннетта в ассоциации таксонов верхней подзоны maximo-divergens граптолитовой зоны gibberulus встречается более 10, а в зоне angustifolius elongatus/angustifolius tenuis – более 20 видов. Комплексная зона обладает “смыкаемостью” границ с ниже- и вышележащими зонами, а нижняя ее граница маркируется первым появлением вида-индекса. Слои с фауной относятся к вспомогательным подразделениям и обычно не имеют “смыкаемости”, располагаясь в беннеттской последовательности между зонами или надстраивая их.
Хроностратиграфический интервал распространения родового таксона Leptograptus Lapworth, определенного А.М. Обутом в одной из коллекций с о. Беннетта, ограничивается поздним ордовиком. Это позволяет считать, что окончание седиментации на о. Беннетта произошло не ранее начала-середины сандбийского века позднего ордовика, а не в среднем ордовике, как предполагали ранее [3, 5].
Ордовикские граптолиты обладают высоким космополитизмом – подавляющее большинство таксонов встречается во многих бассейнах Евразии, Северной и Южной Америки, Австралии и Новой Зеландии. Это объясняется планктонным образом жизни граптолитов, наличием длительных течений и открыто-окраинным генезисом большинства древних морских бассейнов. В противном случае, например, в ограниченно изолированном ордовикском бассейне Сибирской платформы [11] существовали комплексы эндемичных сибирских таксонов граптолитов, значительная часть из которых вела не пелагический, а бентосный образ жизни, например, виды-индексы слоев с Dictyonema kravtsovi, слоев с Dictyonema omnutachense, слоев с Mastigograptus datsenkoi (рис. 2). Палеогеографические построения по ордовикским бентосным фаунистическим сообществам осадочных последовательностей Сибирской платформы, Таймыра, о. Котельный, хр. Сетте-Дабан, Селенняхского кряжа, Верхояно-Чукотской складчатой области и Омулевских гор показали их единство в составе Сибирской биогеографической провинции [12] в мелководном морском бассейне на Сибирском палеоконтиненте.
Для сравнения проведем палеозоогеографический анализ зональных комплексов ордовикских пелагических граптолитовых сообществ Арктических районов Евразии и соседствующих с ними районов Северной Америки. Принципиальные отличия таксономического состава между сообществами пелагических организмов различных палеобассейнов объясняются не столько их удаленностью друг от друга, сколько наличием препятствий для миграций в виде континентальных блоков или глубоководных океанических бассейнов. Зональные комплексы ордовика Таймыра и Новой Земли [13, 14] являются опорными для стратиграфии Российской Арктики, позволяющими проводить межрегиональные и дальнемагистральные корреляции с ключевыми разрезами различных стран [1, 10]. Граптолитовая последовательность о. Беннетта восполняет пробел (две граптолитовые зоны и ряд подзон в интервале верхнего фло – нижнего и среднего дапина), существующий в зональных шкалах Таймыра и Новой Земли. Беннеттские граптолитовые биостратоны от фло по дарривил включительно, позволяют не только точно скоррелировать терригенные образования различных районов Арктики России и Северной Америки (рис. 1, 2), но и оценить степень палеобиогеографических связей между такими палеобассейнами.
По своей дробности и присутствию большинства зональных элементов предлагаемая последовательность зон и слоев с граптолитами о. Беннетта во флоско-дапинском хроностратиграфическом интервале может быть хорошо сопоставима с граптолитовой шкалой северо-западной части Аляски [15], где зафиксированы одноименные зоны approximatus, bifidus (= “protobifidus”), maximo-divergens, Oncograptus, а также слои с Phyl. densus, слои с Is. caduceus nanus, слои с Lev. austrodentatus. Это свидетельствует о стабильно существовавших путях взаимных миграций планктонных граптолитов морских бассейнов о. Беннетта и Аляски (рис. 1). На более высоком средне-дарривильском стратиграфическом интервале зональные биостратоны о. Беннетта и северо-западной части Аляски по граптолитам отличаются друг от друга (рис. 2). В других районах Аляски (например, горы Терра Котта [16]) граптолитовые зоны ордовика не имеют подобного высокого сходства с беннетскими биостратонами (рис. 2).
Современные геодинамические реконструкции с масштабным использованием палеомагнитных данных показали непосредственную близость блоков Новосибирских островов и Арктической Канады в кембро-ордовикском интервале (510–450 млн лет) [17]. С этими данными полностью согласуются приведенные выше сведения по высокому сходству зональных биостратонов о. Беннетта и северо-западной Аляски.
На наличие свободных путей миграции планктонных граптолитов между Беннеттским и Таймырским ордовикскими бассейнами указывают выделенные на Таймыре одноименные зоны approximatus, Oncograptus, artus (= “bifidus”), murchisoni. О таких же открытых связях с Ново-Земельским палеобассейном свидетельствуют одноименные зоны approximatus, Oncograptus, artus (= “bifidus”), murchisoni, а также слои с фауной Is. maximo-divergens и слои с Lev. austrodentatus [13, 14] (рис. 1, 2). В то же время следует обратить внимание, что дапин в разрезах Таймыра и Новой Земли имеет слабую граптолитово-зональную расчлененность по сравнению с дапином Беннетта. Кроме того, на Таймыре на уровне нижнего дарривилла нет каких-либо граптолитовых биостратонов (рис. 2).
В граптолитовых комплексах ордовика о. Беннетта не содержатся таксоны, известные на Сибирской платформе, которая, вследствие мелководности своего бассейна, была практически изолирована от путей миграций планктонных сообществ организмов. Также не имеют сходства ордовикские биостратоны о. Беннетта и одновозрастные граптолитовые последовательности Омулевских гор, о. Шпицберген и Балтики (рис. 1, 2). Только на Шпицбергене выделяются одноименные слои с D. bifidus (= “protobifidus”) [18], а в сводной шкале Балтийских регионов обособляются одноименные зоны angustifolius elongatus и austrodentatus [1, 10].
Ордовикские граптолитовые биостратоны перечисленных регионов Российской части Арктики и Сибири с глубоководными терригенными обстановками демонстрируют между собой значительные различия, что не позволяет предполагать какого-то “Сибирского” палеогеографического единства глубоководных акваторий этих палеобассейнов.
В настоящее время большинство специалистов по граптолитам не разделяют существовавшее ранее мнение о необходимости обособления их глобальных палеозоохорий. Рассматривая различия в группах региональных зональных биостратонов, объединенных в сводные граптолитовые шкалы за счет одноименных их названий, в настоящее время используются термины Лавренция (Северная Америка), Балтика, Гондвана, Южный и Северный Китай [1, 10]. И это не названия палеозоохорий, а наименования континентов и их частей, а также стран, в пределах которых располагаются те или иные геологические структуры древних осадочных бассейнов. Встречаются регионы с экотонным составом граптолитовых сообществ. Например, в разрезах ордовика о. Ньюфаундленд выделяются как граптолитовые зоны Северной Америки, так и Балтики [1, 10, 19]. Интересно отметить, что граптолитовые комплексы западного Ньюфаундленда [19] имеют сходство с граптолитовыми биостратонами о. Беннетта. Там выделяются одноименные зоны approximatus, akzharensis, bifidus (= “protobifidus”), austrodentatus. Дапинские ньюфаундлендские граптолитовые зоны отличаются от беннеттских дапинских биостратонов (рис. 1, 2), что позволяет считать маловероятным существование прямого и устойчивого во времени пути миграции пелагических сообществ организмов между Ньюфаундлендским и Беннеттским ордовикскими бассейнами.
В заключение можно сделать следующие обобщения.
1. Выделенные граптолитовые биостратоны позволяют достоверно зафиксировать в ордовикском разрезе о. Беннетта две ярусные границы Общей (и Международной) стратиграфической шкалы – основание фло и дарривила (соответственно, появление Tet. approximatus и Lev. austrodentatus).
2. Граптолитовая зональная последовательность биостратонов о. Беннетта позволяет провести дальнемагистральную (циркумполярную в современных координатах) корреляцию и создать взаимоувязанный каркас хроностратиграфических интервалов ордовикских терригенных образований различных последовательностей Арктических районов Евразии и Северной Америки.
3. Флоско-дапинский бассейн о. Беннетта с терригенной седиментацией имел открытые связи для широкой масштабной миграции планктонных организмов с одновозрастными бассейнами Аляски. Значительное сходство комплексов ордовикских граптолитов о. Беннетта и Аляски позволяет полагать, что это обусловлено не только тесными палеозоогеографическими связями рассматриваемых бассейнов, но и их непосредственной палеогеографической близостью. Подтверждением этого являются данные палеомагнитных реконструкций взаиморасположения Арктических регионов в ордовике [17], показавших палеогеографическое соседство блоков Аляски и архипелага Де-Лонга.
4. Для дарривильских сообществ пелагических организмов бассейна о. Беннетта объем обмена таксонами с Аляскинскими палеобассейнами резко ограничился. На этом фоне повысилась степень коммуникаций беннеттских граптолитовых сообществ с другими пелагическими сообществами – с ново-земельскими до среднемасштабного и с таймырскими до маломасштабного уровней. Эта палеогеографическая трансформация плана доминирующих путей миграций пелагических организмов могла быть связана с предполагаемым событием аккреции в интервале 470–460 млн лет террейна Арктической Аляски с террейном Новосибирских островов [17].
Список литературы
The Geologic Time Scale 2020. Vol. 2. Elsevier. 2020. P. 563–1357.
Сенников Н.В. Зональные граптолитовые подразделения в ордовике России. Региональная стратиграфия позднего докембрия и палеозоя Сибири. Новосибирск. Изд-во СНИИГГиМС. 2013. С. 45–58.
Геологическая карта СССР масштаба 1:100 000. Лист S/T-53, 54, 55, 56 (Новосибирские острова). Объяснительная записка. Москва. Государственное научно-техническое издательство литературы по геологии и охране недр. 1961. 60 с.
Верниковский В.А., Метелкин Д.В., Толмачева Т.Ю., Малышев Н.А., Петров О.В., Соболев Н.Н., Матушкин Н.Ю. К проблеме палеотектонических реконструкций в Арктике и тектонического единства террейна Новосибирских островов: новые палеомагнитные и палеонтологические данные // ДАН. 2013. Т. 451. № 4. С. 423–429.
Danukalova M.K., Kuzmichev A.B., Sennikov N.V., and Tolmacheva T.Yu. Ordovician turbidites and black shales of Bennett Island (De Long Islands, Russian Arctic), and their significance for Arctic correlations and paleogeography // Geological Magazine. 2020. P. 1207–1237.
Аверьянов Б.Н. Нижне-силурийские граптолиты с р. Колымы. Труды совета по изучению производительных сил. Серия Якутская, вып. 11. Академия наук СССР. 1932. С. 29–34.
Соболевская Р.Ф. О граптолитах ордовика и силура на Новосибирских островах. Граптолиты и стратиграфия. Таллин. 1976. С. 202–208.
Обут А.М. Отдел Гемихордовые. Основы палеонтологии. Иглокожие, гемихордовые. Москва. Изд-во Недра. 1964. С. 279–337.
Maletz J. Part V. Second Revision, Chapter 22: Suborder Axonophora, infraorder Diplograptina 1 (Diplograptidae, Lasiograptidae): Introduction, Morphology, and Systematic Descriptions. Treatise online, Number 127. Lawrence, Kansas, USA. 2019. P. 1–26.
Loydell D.K. Graptolite biozone correlation charts // Geological Magazine. 2012. V. 149. P. 124–132.
Сенников Н.В., Толмачева Т.Ю., Обут О.Т., Изох Н.Г., Лыкова Е.В. Зональное расчленение ордовикских отложений Сибири по пелагическим группам фауны // Геология и геофизика. 2015. Т. 56. № 4. С. 761–781.
Каныгин А.В., Гонта Т.В., Тимохин А.В. Происхождение палеозойских террейнов северо-восточной Азии: геологические доказательства раскола перикратонной окраины Сибирского палеоконтинента и миграции ее фрагментов // Геология и геофизика. 2020. Т. 61. № 11. С. 1459–1475.
Корень Т.Н., Толмачева Т.Ю., Соболевская Р.Ф., Раевская Е.Г., Обут О.Т. Ордовикская система. Зональная стратиграфия фанерозоя России. Санкт-Петербург. Изд-во ВСЕГЕИ. 2006. С. 31–47.
Соболевская Р.Ф. Атлас палеозойской фауны Таймыра. Часть II. Граптолиты ордовика и силура. Санкт-Петербург. Изд-во ВНИИОкеангеология. 2011. 282 с.
Lenz A.C., Jackson D.E. Arenig and Llanvirn graptolite biostratigraphy, Canadian Cordillera. Paleoecology and Biostratigraphy of Graptolites // Geological Society. Special Publication. 1986. № 20. P. 27–45.
Churkin M., Carter Cl. Stratigraphy, structure and graptolites of the Ordovician and Silurian sequence in the Terra Cotta Mountains, Alaska Range, Alaska // U.S. Geological Survey. Professional paper. 1996. N 1555. 84 p.
Metelkin D.V., Chernova A.I., Matushkin N.Yu., Vernikovsky V.A. Early paleozoic tectonics and paleogeography of the Eastern Arctic and Siberia: Review of paleomagnetic and geologic data for the De Long Islands. Earth Science Reviews. 2022. https://doi.org/10.1016/j.earscirev.2022.104102
Cooper R.A., Fortey R.A. The Ordovician Graptolites of Spitsbergen // Bull. Brit. Museum Nat. History (Geol.). 1982. V. 36. № 3. P. 157–302.
Williams S.H., Stevens R.K. Summary account of the Lower Ordovician (Arenig) graptolite biostratigraphy of the Cow Head Group, western Newfoundland // Bulletin Geological Society of Denmark. 1987. V. 35. P. 259–270.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле