

Доклады Российской академии наук. Науки о Земле, 2023, T. 509, № 1, стр. 99-104
Трансформация малакофауны Азовского моря в голоцене и ее роль в формировании береговых кос
Академик РАН Г. Г. Матишов 1, *, Е. П. Коваленко 1, В. В. Титов 1
1 Федеральный исследовательский центр
“Южный научный центр РАН”
Ростов-на-Дону, Россия
* E-mail: matishov_ssc-ras@ssc-ras.ru
Поступила в редакцию 21.11.2022
После доработки 22.11.2022
Принята к публикации 23.11.2022
- EDN: TIMFXC
- DOI: 10.31857/S2686739722602666
Аннотация
Изучение и анализ малакофауны из колонок донных отложений и кернов азовских кос показал характер изменения при переходе от древнеазовских к новоазовским слоям, неравномерность распределения сообществ во времени и пространстве. Раковинный материал из прибрежных малакоценозов и мелководных банок служит основным компонентом для формирования кос. На основании серии кернов мощностью до 25 м с косы Долгой показано, что ее формирование началось около 2500 л.н. и карбонатный материал отражает историю развития бентосной фауны моря.
ВВЕДЕНИЕ
Важнейшими этапами формирования современного состояния Азовского моря, согласно классическим взглядам, стали регрессивно-трансгрессивные циклы [1, 2]. Сопоставляя региональную специфику колебания уровня моря в Азово-Черноморском бассейне, гляциоэвстатические изменения уровня и опираясь на анализ кернов бурения и грунтовых колонок, можно выделить до 7–10 чередовавшихся трансгрессий и регрессий в голоцене [1–3]. В частности, в эпоху фанагорийской регрессии (3.1–2.2 тыс. л.н.) уровень моря был ниже современного на 6–7 м. Формирование дельты Дона и азовских кос происходило в период максимума древнеазовской (4–6 тыс. л.н.) и нимфейской (2.4–1.5 тыс. л.н.) трансгрессий.
Важную информацию по палеогеографии региона и характеру изменения уровня моря и особенностей гидрологического режима дает анализ строения береговых кос Таганрогского залива, и собственно, донные отложения Азовского моря. Основной причиной роста аккумулятивных кос азовского типа явилось то, что в береговую зону непрерывно поступал интенсивно продуцируемый органогенный ракушечный материал из акватории Азовского моря. Крупнейший источник наносов для формирования надводно-подводной части косы Долгой, очевидно, существовал в районе банок Железинской, Ахтарской, Елениной [1, 2].
Сведения о составе и среде обитания рецентных бентосных организмов представлены в ряде трудов [4–8]. Имеется и ряд сводок, охватывающих голоценовую историю развития малакофауны этого моря [9]. Учитывая частую встречаемость и высокую информативность раковинного материала, в основу работы положен детальный малакофаунистический анализ образцов, полученных при бурении береговых аккумулятивных форм и отборе на шельфе колонок донных отложений, в комплексе с серией полученных радиоуглеродных датировок.
МАТЕРИАЛЫ И МЕТОДИКА
В ходе бурения, с 2017 по 2022 г., колонковым и ударно-канатным методами на Беглицкой, Павло-Очаковской, Чумбурской, Должанской косах, а также в дельте рек Кагальник и Дон нами получено 30 кернов мощностью от 10 до 26.5 м. На малакофаунистический анализ из них было взято порядка 140 образцов. Отбор колонок грунта осуществлялся с борта НИС “Денеб” прямоточной грунтовой трубкой с утяжелителями и лепестковым клапаном. В работе использованы результаты анализа малакофауны из трех колонок, полученных в Азовском море в 2019 г., мощностью 260, 298 и 300 см (рис. 1).
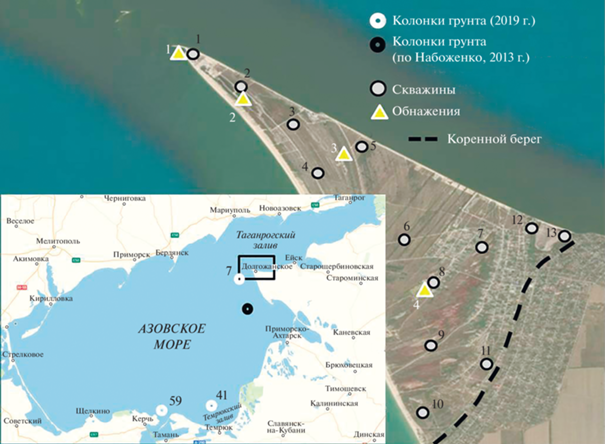
В фокусе литологических и малакофаунистических работ находится самая крупная в Азовском море коса Долгая (рис. 1). Из слоев кернов, содержащих раковинный материал, отбирались пробы на промежутках мощностью 5–10 см. Был также получен ракушечный материал из 8 обнажений мощностью от 95 до 250 см, расположенных в корневой, центральной и дистальной частях косы. Из них материал собирался из слоев, содержащих целые створки раковин, и прослоев, сложенных раковинным детритом, с шагом не более 50 см (чаще 20–25 см). Таксономическое определение малакофауны проводилось с использованием ряда источников [5, 6].
Для определения абсолютного возраста отбирались навески массой 15–40 г преимущественно сомкнутых раковин Cerastoderma Poli, 1795. Получено 55 радиоуглеродных датировок в Лаборатории геоморфологических и палеогеографических исследований СПбГУ. Значения календарного возраста приведены на основании калибровочной программы “OxCal 4.4.1”.
ИССЛЕДОВАНИЕ МАЛАКОФАУНЫ
Аккумулятивное тело косы Долгой было образовано в интервале времени от 1920 ± 110 (ЛУ-9756) до 2500 ± 150 лет назад (ЛУ-9757), в период нимфейской трансгрессии. В целом эта трансгрессия ассоциируется с накоплением отложений новоазовского возраста (от 3.1 тыс. л.н.), которые имеют повсеместное распространение в Азовском море [0, 1, 10].
Основную массу отложений карбонатного материала косы Долгой составляют раковины эвригалинных видов Cerastoderma glaucum, Abra segmentum, Hydrobia sp., способных выносить опреснение до 5‰ (табл. 1). Черноморские виды – маркеры древнеазовской фауны: Gastrana fragilis, Polititapes aureus, Chamelea gallina, Lucinella divaricata, встречаются в пробах эпизодически и приурочены к глубоко залегающим слоям. Солоноватоводный моллюск Theodoxus pallasi, обитающий при оптимуме солености 5–7‰, массово отмечен в отложениях лиманного типа; единичные его занесенные экземпляры встречаются в раковинном чехле косы.
Таблица 1.
Раковины моллюсков, обнаруженные в отложениях косы Долгой
| Азово-черноморские виды (новоазовские, 3.1–0 тыс. л.н.1, и древнеазовские отложения) | Черноморские виды (маркеры древнеазовских отложений, 8.3–4 тыс. л.н.1) |
|---|---|
| Соленость >5‰2 | Соленость >15–17‰ |
| *Cerastoderma glaucum Poiret, 1789 | *Gastrana fragilis (L., 1758) |
| *Abra segmentum (Récluz, 1843) | Solen vagina L., 1758 |
| Mytilaster lineatus (Gmelin, 1790) | *Chamelea gallina (L., 1758) |
| *Lentidium mediterraneum (O.G. Costa, 1830) | *Lucinella divaricata (L., 1758) |
| *Hydrobia acuta (Draparnaud, 1805) | Loripes lucinalis (Lamarck, 1818) |
| Соленость >10–12‰ | *Polititapes aureus (Gmelin, 1791) |
| *Mytilus galloprovincialis Lamarck, 1819 | *Parvicardium exiguum (Gmelin, 1791) |
| *Barnea candida (L., 1758) | Bela nebula (Montagu, 1803) |
| *Bittium reticulatum (da Costa, 1778) | Turbonilla delicata (Monterosato, 1874) |
| *Rissooidea Gray, 1847 | Tragula fenestrata (Jeffreys, 1848) |
| *Tritia reticulata (L., 1758) | |
| Setia valvatoides (Milaschevitsch, 1909) | |
| *Cylichnina variabilis Milaschewitsch, 1912 | |
| *Parthenina interstincta (J. Adams, 1797) | |
| *Ebala pointeli (de Folin, 1868) | |
| *Retusa truncatula (Bruguière, 1792) | |
| Солоноватоводные виды | Виды-вселенцы II половины XX века |
| Dreissena polymorpha (Pallas, 1771) (<5–7‰)2 | *Anadara kagoshimensis (Tokugana, 1906) (10–35‰)2 |
| Theodoxus pallasi Lindholm, 1924 (<10–12‰) | Mya arenaria L., 1758 (6–17‰) |
Ключевую роль в образовании карбонатных осадков в Азовском море играет C. glaucum, характеризующийся высокой продуктивностью и прочностью створок, а также A. segmentum и Mytilaster lineatus. Значительный вклад в этот процесс вносят вселившиеся в прошлом веке моллюски Anadara kagoshimensis и Mya arenaria [11]. Общая годовая продукция зообентоса в Азовском море оценивается в 19–20 млн т [4], из них продукция C. glaucum составляет 13–14 млн т в год, достигая биомассы 2 кг/м2 [11].
Коса Долгая развивалась, приобретала форму прислоненных береговых валов из детрита, в течение более двух тысяч лет. Формирование косы продолжается за счет выброса свежих раковин и живых моллюсков во время сильных ветров (скорость выше 20 м/с) и штормового волнения моря. Накопления карбонатного материала сходны по своему составу и состоят из отложенного и переотложенного материала. Для этой толщи характерно чередование слоев, вмещающих целые створки, и прослоев, содержащих раковинный детрит и песок. В центральной части косы раковинный материал мощностью до 2.5 м представлен комплексом видов одинаковой сохранности C. glaucum – A. segmentum – Barnea candida – Hydrobia sp. с безусловным доминированием по объему раковин и детрита церастодермы, со значительным вкладом таковых от абры. Кроме того, отмечены малочисленные Hydrobia sp., Lentidium mediterraneum и Th. pallasi. При продвижении к подошве изученных разрезов увеличивается количество створок абры и улучшается их сохранность, появляются тонкораковинные удлиненные формы створок церастодермы. В дистальной и прикорневой части косы отложения более плотные; спрессованные прослои детрита и песка мощнее слоев, содержащих целые створки. В центральной части чехол более рыхлый. Основная масса раковин представлена видами, обитающими в настоящее время в акватории косы Долгой. Отмечены малочисленные переотложенные створки раковин черноморских моллюсков Ch. gallina, G. fragilis и Solen vagina.
При бурении были вскрыты более древние горизонты, вмещающие раковинный материал новоазовской (современная азовская) и древнеазовской (обедненный вариант современной черноморской) малакофаун. Раковины голоценовых моллюсков были отмечены до глубины 7.8–8.0 м. В частности, современный малакоценоз Hydrobia–Cerastoderma–Abra–Lentidium–Parthenina, обитающий в районе косы Долгой, отмечен в горизонтах, глубже 420 см (скв. 3) и 320 см (скв. 6). Раковины из этих горизонтов датированы от 910 ± 100 до 2240 ± 130 л.н. Переходный вариант фауны, возрастом 2980±100 л.н., отмечен на глубине 550–570 см (скв. 5). Содержащиеся в горизонте раковины гастропод Bittium reticulatum, Rissoa sp., Hydrobia sp. могут свидетельствовать о наличии водной растительности, A. segmentum – о мелководности и возможном дефиците кислорода в условиях лагуны или залива. В горизонте 430–440 см (скв. 9; 2890 ± 90 л.н.) отмечены растительные остатки и многочисленные раковины эврибионтных Hydrobia, Cerastoderma и Abra, что подтверждает существование залива или лимана с водной растительностью, соленость которого была близка к современной.
Древнеазовский малакофаунистический комплекс в дистальной части косы, отмеченный в слое 750–780 см (скв. 1; 7860 ± 100 л), кроме азовоморской фауны содержит целые раковины ювенильных Ch. gallina и Tritia reticulata. Схожие по таксономическому составу, типу донных осадков и содержанию крупных обломков и детрита раковин сем. Mytilidae слои, вскрытые в интервалах 615–780 см (скв. 3) и 650–670 см (скв. 4), вмещают хорошо сохранившиеся раковины древнеазовских видов L. divaricata, G. fragilis, Parvicardium exiguum и створки P. aureus. Грунт здесь отличается большим содержанием мелких обломков створок митилид, указывающих на динамичность происходивших процессов.
В прикорневой части косы на восточном берегу скважиной 13 были вскрыты слои, содержащие малакофауну с другими экологическими характеристиками. В горизонте 158–165 см (6560 ± 90 л.н.) в глинистом иле темно-серого цвета отмечен таксоценоз лагунного типа. Руководящим видом здесь выступает Hydrobia, многочисленны Abra и Rissoa, малочисленны Cerastoderma и Mytilaster. Значимым компонентом является солоноватоводный Th. pallasi, который указывает на соленость водоема не более 10–12‰. Вероятно, на месте современного озера Лебяжьего на косе Долгой был лиман, имевший слабое сообщение с морем или полностью изолированный от него. В вышележащих слоях этого керна (100–110 см) место Th. pallasi занимает гастропода Setia valvatoides, обитающая в настоящее время в лиманах Таманского залива на водной растительности, что свидетельствует о более интенсивном сообщении лимана с морем и увеличении его солености до морской.
Ранее опубликованные нами данные [10] и анализ колонок донных отложений позволил обозначить источники поступления раковинного материала на косу Долгая. Одним из них, помимо прибрежных малакоценозов, указывались отложения ракуши в районе Железинской банки. Поверхностные слои донных отложений (до глубины 90 см) на северо-западной оконечности банки содержат большое количество раковин моллюсков рода Cerastoderma [10]. При этом до глубины 50 см залегает ракушечный слой, возрастом 2400 ± 180 л, содержание карбонатного материала (преимущественно створок Bivalvia) в котором превышает 40%. Наличие в этих слоях раковин черноморского G. fragilis может свидетельствовать о непрерывных процессах отложения, выноса и переотложения карбонатного материала.
На банке Еленина (ст. 7) в непосредственной близости к косе Долгой была отобрана колонка грунта. Вскрытые ею слои донных осадков, возрастом от 550 ± 80 л у поверхности до 2930 ± 130 л в основании колонки, показали, что здесь на алевритово-илистых песках развивался таксоценоз моллюсков, характерный для Азовского моря, с руководящими видами C. glaucum и Hydrobia acuta и малочисленными A. segmentum, Parthenina interstincta и Cylichnina variabilis. В нижележащих слоях, глубже горизонта 60–70 см (1960 ± 250 л.н.), значимым компонентом сообщества становится B. reticulatum, выдерживающий понижение солености до 10‰ [11], который в отложениях косы Долгой был массово представлен в слоях старше 4000 л. Здесь также отмечены солелюбивые морские виды Retusa truncatula и Ebala pointeli и обитатель прибрежной растительности Rissoa sp.
В отобранных в юго-восточной части Азовского моря грунтовых колонках (ст. 41 и 59) обнаружен более разнообразный таксоценоз моллюсков. Руководящими видами в поверхностных слоях и до глубины 120–130 см (1260 ± 70 л.н.) являются Cerastoderma, Abra, Hydrobia и Lentidium. Содоминантами на ст. 59 выступают Bittium и Parthenina, на обеих станциях многочисленны гастроподы Ebala, Rissoa, Cylichnina и Retusa. В нижележащих слоях, начиная с глубины 130 см, появляются значимые отличия в составе малакофауны. Наблюдается резкое снижение количества раковин L. mediterraneum на обеих станциях и раковин Abra на ст. 59. Для ст. 41 в слоях глубже 200 см (более 2190 ± 110 л.н.) характерно значительное увеличение доли гастропод Bittium и Rissoa, что может указывать на существование мелководий с водной растительностью. На ст. 59 в слое 185–200 см (3140 ± 180 л.н.) комплекс руководящих видов включает Cerastoderma, Hydrobia и Bittium, а начиная с 153–165 см (2280 ± 150 л.н.) в пробах встречаются единичные экземпляры створок Ch. gallina и P. aureus – маркеров древнечерноморской фауны.
ОБСУЖДЕНИЕ
Проанализировав структуру таксоценозов моллюсков, извлеченных из кернов скважин и обнажений на косе Долгой, был выявлен ряд закономерностей. Осадочный чехол на косе сложен раковинами разных возрастов и разных фаун. Здесь встречаются как современные моллюски новоазовской фауны, так и черноморские виды, обитавшие в период древнеазовской трансгрессии. В поверхностных слоях отмечается явное смешение раковин обеих фаун, что может быть связано с волновой активностью и, как следствие, с переотложением раковинного материала; до 90% карбонатного материала здесь представлено створками Cerastoderma.
Аккумулированный пласт карбонатного материала имеет неоднородную слоистую структуру, для которой характерно чередование рыхлых слоев с хорошо сохранившимися створками, и прослоев плотного детрита с илистым и песчаным заполнением. Изучение обнажений в дистальной и центральной части косы показали, что есть отличия в составе и сохранности материала. Наличие в дистальной части целых массивных створок Cerastoderma и Chamelea и призамочных обломков более хрупких экземпляров Barnea и Abra может говорить об интенсивной волновой активности в период формирования этого участка косы. Хорошая сохранность большинства видов в центральной части может свидетельствовать о меньшей динамике вод и о меньшей интенсивности размыва и переотложения материала.
В глубоких слоях кернов вскрываются донные осадки водоемов, существовавших до момента формирования косы. Здесь отмечается раздельное отложение фаун, что позволяет выявлять изменение условий обитания и, как следствие, состава малакоценозов.
ЗАКЛЮЧЕНИЕ
Отбор колонок донных отложений мощностью до 4 м и бурение осадочных толщ с отбором кернов мощностью до 25 м на азовских косах (в частности косе Долгой), а также малакофаунистические исследования в комплексе с абсолютным датированием раковин дали возможность по-новому охарактеризовать зообентос различных этапов голоцена и реконструировать историю колебания уровня моря в Азовском бассейне за последние 5 тыс. лет. Современный уровень морей и океанов возник в эпоху климатического оптимума 6–4 тыс. л.н. [12]. Массовые заморы донной фауны во время развития нимфейской трансгрессии в интервале времени от 1920 ± 110 (ЛУ-1756) до 2500 ± 150 (ЛУ-9757) лет привели к накоплению на стыке Таганрогского залива Азовского моря огромной массы биогенных отложений.
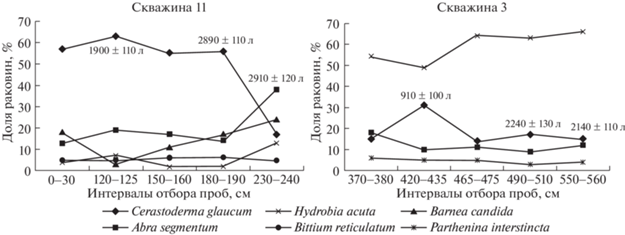
Вскрытая скважинами верхняя часть отложений косы Долгой сложена ракушечным материалом с примесью глины и суглинка [2]. В разных горизонтах ракушечных отложений преобладают раковины двустворчатого моллюска C. glaucum. В поверхностных горизонтах доля раковин этого моллюска колеблется от 96 до 57%. К основанию разрезов увеличивается доля A. segmentum (до 38%), B. candida (до 24%) и H. acuta (рис. 2). Мощность отложений из ракуши увеличивается от корневой к дистальной части косы от 4 до 7 м. Верхнеголоценовые слои ракуши часто подстилаются горизонтами, имеющими признаки субаэральных условий формирования. Лагунные отложения часто имеют черный оттенок с типичным запахом сероводорода. Очевидно, возникли они в эпоху фанагорийской регрессии 3.1–2.5 тыс. л.н. В целом малакофауна, обнаруженная в колонках на шельфе и кернах на косах, практически идентична. Расхождение в количестве таксонов можно объяснить отличиями условий в местах отбора колонок (глубоководные участки) и кернов (прибрежье, мелководье) (табл. 1).
Рис. 2.
Доля створок и раковин руководящих видов моллюсков в разных горизонтах ракушечных отложений косы Долгая.
Крупные источники раковинного материала для формирования тела береговых кос азовского типа, очевидно, существовали на юго-востоке азовского шельфа в районе банок Железинской, Ахтарской и Еленина, в западной прибрежной части в районе Арабатской стрелки, а также в заливах Азовского моря [1, 2, 11]. Можно предположить, что значительное количество раковин моллюсков, составляющих основную часть тела кос, формировалось в благоприятных для развития бентосных организмов условиях в период нимфейской трансгрессии.
Внутривековая цикличность климата обусловливала заметные вариации гидрометеорологических условий в Приазовье. Например, в течение прошлого тысячелетия и в XX–XXI вв. засухи, циклы маловодья повторялись с интервалом порядка 30–40 лет [13]. Перенос и отложение раковинного материала в районе кос осуществлялись за счет штормовых волн, сформировавших серию береговых валов (например, на косе Долгой их насчитывается до пятидесяти), отчетливо выделяющихся на поверхности прибрежных аккумулятивных образований.
Список литературы
Матишов Г.Г., Польшин В.В., Титов В.В., Шевердяев И.В. Голоценовая история азовского шельфа // Наука Юга России. 2019. Т. 15. № 1. С. 42–53. https://doi.org/10.7868/S25000640190105
Матишов Г.Г., Польшин В.В., Коваленко Е.П., Григоренко К.С. Палеоокеанология Азовского моря в голоцене (по данным бурения и изучения малакофауны на косе Долгой) // Океанология. 2020. Т. 61. № 4. С. 609–619. https://doi.org/10.31857/S0030157421030084
Матишов Г.Г., Ковалева Г.В., Арсланов Х.А., Дюжова К.В., Польшин В.В., Золотарева А.Е. Результаты радиоуглеродного датирования голоценовых отложений Азовского моря // ДАН. 2018. Т. 479. № 6. С. 655–658. https://doi.org/10.7868/S0869565218120113
Воробьев В.П. Бентос Азовского моря. Труды АзЧерНИРО. Симферополь: Крымиздат, 1949. 195 с.
Определитель фауны Черного и Азовского морей. Т. 3. Свободноживущие беспозвоночные. Киев: Наукова думка, 1972. 340 с.
Анистратенко В.В., Халиман И.А., Анистратенко О.Ю. Моллюски Азовского моря. Киев: Наукова думка, 2011. 173 с.
Матишов Г.Г., Шохин И.В., Набоженко М.В., Польшин В.В. Многолетние изменения донных сообществ Азовского моря в связи с характером осадконакопления и гидрологическим режимом // Океанология. 2008. Т. 48. № 3. С. 425–435.
Чухчин В.Д. Экология брюхоногих моллюсков Черного моря. Киев: Наукова Думка, 1984. 176 с.
Невесская Л.А. Позднечетвертичные двустворчатые моллюски Черного моря, их систематика и экология. Академия наук СССР. Труды Палеонтологического института. Т. 105. Москва: Наука, 1965. 391 с.
Набоженко М.В. Реконструкция и динамика таксоценоза двустворчатых моллюсков (Mollusca: Bivalvia) Азовского моря в позднем голоцене в связи с изменением солености // Труды зоологического института РАН. Приложение № 3. Пятьдесят лет концепции критической солености. 2013. Т. 13. № 3. С. 182–191.
Ивлиева О.В., Фроленко Л.Н. Биогенное карбонатонакопление донных отложений Азовского моря во второй половине XX века. // Известия ВУЗов. Северо-Кавказский регион. Естественные науки. 2009. № 4. С. 96–100.
Матишов Г.Г., Польшин В.В., Кулыгин В.В., Титов В.В., Коваленко Е.П., Сушко К.С. Новые данные о строении косы Долгой Азовского моря (бурение, исследование обнажений, малакофауна) // Наука Юга России. 2020. Т. 16. № 3. С. 26–39. https://doi.org/10.7868/S25000640200304
Матишов Г.Г., Дашкевич Л.В., Кириллова Е.Э. Цикличность климата в Приазовье: голоцен и современный период (XIX-XXI вв.) // Доклады РАН. Науки о Земле. 2021. Т. 498. С. 96–100. https://doi.org/10.31857/S2686739721050091
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о Земле