

Доклады Российской академии наук. Науки о жизни, 2020, T. 490, № 1, стр. 85-89
БЫСТРАЯ МОРФОЛОГИЧЕСКАЯ ДИВЕРСИФИКАЦИЯ КАРПОВОЙ РЫБЫ Oreoleuciscus potanini (CYPRINIDAE) ПРИ ОБРАЗОВАНИИ ВОДОХРАНИЛИЩА НА РЕКЕ СЕМИАРИДНОЙ ЗОНЫ
Академик РАН Ю. Ю. Дгебуадзе 1, 2, *, А. Н. Мироновский 1, 3, Б. Мэндсайхан 4, Ю. В. Слынько 5
1 Институт проблем экологии и эволюции
им. А.Н. Северцова Российской Академии наук
Москва, Россия
2 Московский государственный университет
им. М.В. Ломоносова
Москва, Россия
3 Институт биологии внутренних вод им. И.Д. Папанина Российской Академии наук
Ярославской обл., п. Борок, Россия
4 Institute of Geography and Geoecology of Mongolian Academy of Sciences
15170 Ulanbaatar, Mongolia
5 Институт биологии южных морей
им. А.О. Ковалевского Российской Академии наук
Севастополь, Россия
* E-mail: yudgeb@gmail.com
Поступила в редакцию 08.12.2019
После доработки 08.12.2019
Принята к публикации 08.12.2019
Аннотация
Анализ фенетического разнообразия алтайского османа Потанина Oreoleuciscus potanini Тайширского водохранилища, построенного в 2007 г. на р. Завхан (Котловина Больших Озер, Монголия), выявил четыре морфологически различаемые формы. Ранее на речном участке р. Завхан описано две формы O. potanini, в озерах Котловины Больших Озер – три формы. Полученные данные демонстрируют сходство с цикличным формообразованием у близкого вида Oreoleuciscus humilis в водоемах Долины озер, также относящимся к Центрально-Азиатскому бессточному бассейну.
Котловина Больших Озер находится в семиаридной зоне, в западной части Центрально-Азиатского бессточного бассейна, где реки Кобдо и Завхан образуют систему озер. В озерах один из немногих обитающих там видов рыб, алтайский осман Потанина Oreoleuciscus potanini (Kessler, 1879), образует три морфо-экологические формы: растительноядную, рыбоядную и острорылую [1]. В среднем течении р. Завхан обнаружены две морфологические формы O. potanini, одна из которых близка растительноядной форме системы озер Хар-Ус – Дургун [2].
Цель настоящей работы – исследование фенетического разнообразия популяции O. potanini Тайширского водохранилища, в 2007 г. построенном в среднем течении р. Завхан.
Исследование выполнено на материале, собранном гидробиологическим отрядом Совместной Российско-Монгольской комплексной биологической экспедиции РАН и Академии наук Монголии. Первые пробы рыб (21 экз. SL = 129–201 мм) брали из сетных уловов (ячея: 20, 30, 40, 50 и 60 мм) в приплотинном участке Тайширского водохранилища 20 августа 2010 г. Теми же орудиями лова в том же месте взяты пробы 10 августа 2012 г. (25 экз. SL = 142–298 мм); 11 августа 2013 г. (16 экз. SL = 154–360 мм) и 24–26 августа 2015 г. (43 экз. SL = 128–386 мм). В качестве реперов использованы выборки форм “а” (19 экз. SL = 133–188 мм) и “б” (18 экз. SL = 135–167 мм) османов речного участка р. Завхан, взятые в 2008 г. [2].
Головы рыб вместе с костью плечевого пояса cleithrum фиксировали поваренной солью, и после препарирования измеряли 15 остеологических параметров: базальную длину черепа (BL); расстояние между внешними краями frontalia, pterotica и sphenotica; ширину черепа на уровне соединения frontale и pteroticum; высоту черепа на уровне изгиба parasphenoideum и заднего края parasphenoideum; высоту hyomandibulare; расстояние между крайними точками ветвей глоточного зуба; длину praemaxillare; длину praeoperculum, interoperculum, dentale и maxillare; длину по диагонали cleithrum [4, 5].
В расчётах средствами пакета биостатистики NTSYS 1.7 [6] использовали индексы, вычисляемые как отношения абсолютных значений промеров к длине черепа BL. В анализе главных компонент собственные векторы считали по корреляционной матрице; длину вектора принимали равной 1. Разобщенность объектов в многомерном пространстве индексов оценивали, используя таксономическую дистанцию [6]. Дендрограммы строили методом upgma на уровне особей и методом single linkage на уровне выборок.
Результаты анализа фенетического разнообразия множества из 105 особей (объединенная выборка за годы наблюдений) на рис. 1 показывают, что в пространстве рассматриваемых признаков анализируемая совокупность особей образует четыре в разной степени разобщенные морфологические формы (рис. 1а). С составом кластеров дендрограммы совпадает состав полигонов, разобщённых на плоскости главных компонент (рис. 1б).
Рис. 1.
Морфологическое разнообразие O. potanini в объединённой выборке Тайширского водохранилища (2010, 2012, 2013 и 2015 гг.). а – дендрограмма различий; б – распределение на плоскости ГК1 и ГК2. 1 – особи формы 1, 2 – особи формы 2, 3 – особи формы 3, 4 – особи формы 4.
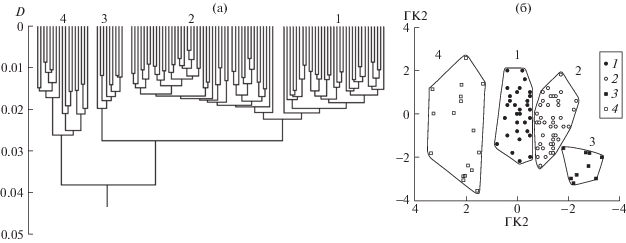
В исследуемый период предельная длина рыб год от года увеличивалась [3] и дифференциация на рис. 1 может отражать различия между крупными и мелкими особями мономорфной популяции в результате постепенного изменения пропорций по мере роста. Однако, как видно из рис. 2, где по осям отложена длина основания черепов особей BL и ГК1 индексов промеров, контуры образовавшегося скаттера можно рассматривать как онтогенетический канал, т.е. проекцию области многомерного пространства признаков, где располагаются индивидуальные онтогенетические траектории [7]. Анализ взаиморасположения каналов позволяет оценить соотношение различий, обусловленных разными размерами особей, с различиями, которые разницей размеров объяснить нельзя [7, 8].
Рис. 2.
Онтогенетические каналы O. potanini Тайширского водохранилища. а – каналы форм 1 (1) 1 и 2 (2); б – форм 1 (3) и 3 (4). в – форм 1 (1) и 4 (4); г – форм 2 (2) и 3 (3); д – форм 2 (2) и 4 (4); е – форм 3 (3) и 4 (4).
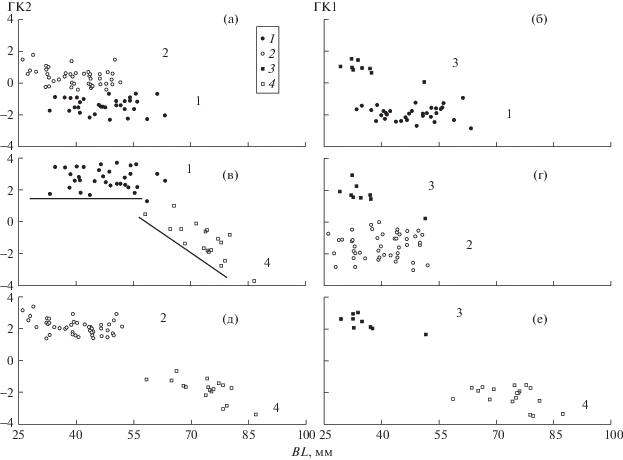
Как видим, разницей размеров обусловлены могут быть отличия лишь формы 4, онтогенетический канал которой во всех сочетаниях начинается после завершения онтогенетического канала сопоставляемых с нею форм (рис. 2в, 2д и 2е). Плавно, без хиатусов по осям смена каналов происходит лишь на рис. 2в, что даёт основания предположить, что для форм 1 и 4 канал общий и форма 4 – это крупные особи формы 1. Если это так, речь не о постепенном, а скачкообразном изменении пропорций по достижению BL ~ 56 мм, о чём свидетельствует резкое изменение наклона общего для форм канала (если он общий) по отношению к осям координат. В обсуждаемом контексте такая трансформация – явление не менее (если не более) интересное, чем симпатрия форм, развивающихся в разных онтогенетических каналах. Сопоставление каналов форм 1, 2 и 3 (рис. 2а, 2б и 2г) не даёт оснований полагать, что различия между ними обусловлены размерными изменениями.
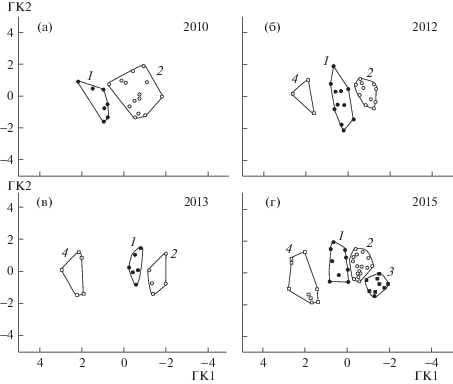
Распределение особей, взятых в водохранилище в разные годы, иллюстрирует рис. 3. В 2010 г. проба представлена лишь формами 1 и 2 (рис. 3а). В пробе 2012 г. появляется форма 4 (рис. 3б); в пробе 2013 г. относительная доля формы 4 увеличивается (рис. 3в). В 2015 г. появляется форма 3 (рис. 3г).
Рис. 3.
Распределение форм O. potanini на плоскости ГК в пробах разных лет. а – проба 2010 г; б – проба 2012 г.; в – проба 2013 г.; г – проба 2015 г. Обозначение форм, как на рис. 1 и 2.
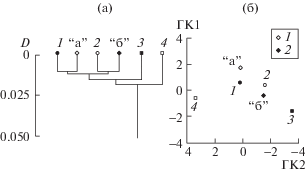
Анализ рис. 4 позволяет предположить, что формы 1 и 2 Тайширского водохранилища – это дериваты форм “а” и “б” р. Завхан, развивающиеся при смене лотических условий лентическими: и на дендрограмме (рис. 4а), и на плоскости главных компонент (рис. 4б) форма “а” из речного участка близка форме 1, а форма “б” близка форме 2. В пользу этого предположения свидетельствует и то, что в 2010 г. в пробах из водохранилища присутствовали особи лишь форм 1 и 2. Предположения о происхождении форм 3 и 4 на данном этапе исследований представляются преждевременными.
Рис. 4.
Фенетические отношения форм O. potanini на уровне средневыборочных оценок. а – дендрограмма сходства; б – распределение на плоскости ГК1 и ГК2. 1 – речная форма “а”; 2 – речная форма “б”; прочие обозначения как на рис. 1–3.
Проведённое исследование показывает, что с течением лет фенетическая изменчивость популяции Тайширского водохранилища увеличивается за счёт появления новых морфологических форм. Анализ онтогенетических каналов показывает, что различия форм не могут быть результатом постепенного изменения остеологических пропорций по мере роста рыб и являются либо результатом независимого развития в разных онтогенетических каналах, либо тем, что формы развиваются в одном канале и при достижении некоторых размеров одна форма переходит в другую путём быстрой трансформации.
Следует отметить, что формирование популяции O. potanini Тайширского водохранилища происходит в условиях исключительно бедной ихтиофауны водоема семиаридной зоны. В водохранилище отмечены еще лишь два вида рыб: монгольский хариус Thymallus brevirostris и сибирский усатый голец Orthrias barbatulus toni. Полученные данные демонстрируют сходство с установленным ранее цикличным формообразованием у близкого вида Oreoleuciscus humilis в периодически высыхающих водоемах Долины озер, также относящихся к Центрально-Азиатскому бессточному бассейну и тоже характеризующихся исключительно бедной ихтиофауной [9, 10].
В дальнейших исследованиях O. potanini Тайширского водохранилища необходимо попытаться выявить наследственную и экологическую составляющие обнаруженной морфологической диверсификации.
Список литературы
Баасанжав Г., Дгебуадзе Ю.Ю., Демин А.Н. и др. Рыбы МНР. 1983. М.: Наука. С. 102–224.
Дгебуадзе Ю.Ю., Мироновский А.Н., Мендсайхан Б. и др. ДАН. Общ. биология. 2017. Т. 473. № 2. С. 250–253. https://doi.org/10.7868/S0869565217080266
Мендсайхан Б., Дулмаа А., Крылов А.В. и др. Аридные экосистемы. 2016. Т. 22. № 3. С. 84–91. https://doi.org/10.1134/S2079096116030082
Мироновский А.Н. Вопр. ихтиологии. 2006. Т. 46. № 2. С. 240–251. https://doi.org/10.1134/S0032945206020056
Дгебуадзе Ю.Ю., Мина М.В., Мироновский А.Н. // Вопросы ихтиологии. 2008. Т. 48. № 3. С. 315–323. https://doi.org/10.1134/S0032945208040024
Rohlf F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System. Vers. 1.7. N.Y.: Exeter software, 1992. 356 p.
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. // J. Fish Biology. 1996. V. 48. P. 383–404.
Мироновский А.Н., Касьянов А.Н., Слынько Ю.В. и др. // Вопросы ихтиологии. 2014. Т. 54. № 1. С. 25–31. https://doi.org/10.7868/S0042875214010081
Dgebuadze Yu.Yu. // Hydrobiologia. 1995. V. 303. P. 235–245.
Dgebuadze Yu.Yu. In: Ecosystems of Central Asia under Current Conditions of Socio-economic Development. 2015, Ulaanbaatar, Mongolia. V 2. P. 29–33.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни