

Доклады Российской академии наук. Науки о жизни, 2020, T. 490, № 1, стр. 90-93
ОБТЮРАКУЛЫ ВЕСТИМЕНТИФЕР (ANNELIDA, SIBOGLINIDAE) ГОМОЛОГИЧНЫ ДОРСАЛЬНЫМ ГУБАМ ПОЛИХЕТ СЕМЕЙСТВА SABELLIDAE
Н. П. Карасева 1, Н. Н. Римская-Корсакова 1, М. М. Ганцевич 1, *, академик РАН В. В. Малахов 1
1 Московский государственный университет
им. М.В. Ломоносова
Москва, Россия
* E-mail: mgantsevich@gmail.com
Поступила в редакцию 16.10.2019
После доработки 16.10.2019
Принята к публикации 16.10.2019
Аннотация
Проведен сравнительный анализ строения дорсальных губ полихеты Eudistylia polymorpha из семейства Sabellidae и обтюракул вестиментиферы Oasisia alvinae. Сделан вывод о том, что обтюракулы вестиментифер являются гомологами дорсальных губ полихет семейства Sabellidae. Высказано предположение о том, что головная лопасть сибоглинид подсемейства Frenulata гомологична слившимся обтюракулам Vestimentifera.
Вестиментиферы – бескишечные морские черви, обитающие в очагах подводного вулканизма или в местах холодных углеводородных просачиваний. Жизнедеятельность вестиментифер обеспечивается хемоавтотрофными бактериями. Согласно современным представлениям, вестиментиферы – это кольчатые черви, которые составляют отдельное подсемейство в пределах семейства Siboglinidae [1]. В то же время положение Siboglinidae в системе Annelida остается неопределённым. Данные молекулярной филогенетики не могут внести ясность в этот вопрос. В зависимости от выбранных методов анализа одни исследователи сближают Siboglinidae с Oweniida [2], другие – с Cirratulidae [3], третьи – с Clitellata, Echiurida и Sipunculida [4]. На фоне такого разнообразия мнений стоит вспомнить, что ещё один из первых исследователей сибоглинид, российский зоолог П.В. Ушаков [5] рассматривал этих червей как представителей семейства Sabellidae. Мнение о близости сибоглинид к полихетам семейства Sabellidae высказывалось и в ряде современных работ [6, 7]. Всё это повышает ценность сравнительно-морфологических исследований, тем более, что сибоглиниды имеют много особенностей строения, которые отличают их от типичных аннелид.
Подобно многим седентарным аннелидам, сибоглиниды имеют щупальца на переднем конце тела. Наиболее развитым щупальцевым аппаратом обладают представители Vestimentifera, у которых щупальца организованы в многочисленные щупальцевые ламеллы [8–10]. Щупальцевый аппарат вестиментифер поддерживается обтюракулами – двумя симметричными лопастями, представляющими собой выросты переднего конца тела [10, 8, 9 ]. А.В. Иванов [11] предполагал, что обтюракулы – это преобразованные личиночные щупальца первой пары. По данным других авторов [12], личиночные щупальца редуцируются и не имеют гомологов у взрослых форм. Проблема гомологии обтюракул остается не решенной вплоть до настоящего времени. Цель настоящей работы – попытаться обосновать гомологию обтюракул Vestimentifera и дорсальных губ седентарных аннелид семейства Sabellidae.
В качестве объектов исследования были выбраны представитель семейства Sabellidae – Eudistylia polymorpha (Johnson, 1901) и представитель вестиментифер Oasisia alvinae Jones, 1985. Полихеты E. polymorpha собраны в прибрежной зоне на биологической станции “Восток” Национального научного центра морской биологии им. А.В. Жирмунского в заливе Восток Японского моря на глубине 1 м. Особи вестиментиферы O. alvinae были добыты в 2003 г. в 49-м рейсе НИС “Академик Мстислав Келдыш” с использованием глубоководных обитаемых аппаратов “Мир-1”и “Мир-2” в гидротермальном оазисе Восточно-Тихоокеанского поднятия в точке с координатами 09°50.53' с.ш., 104°17.52' в.д. на глубине 2524 м. Материал зафиксирован жидкостью Буэна, обезвожен в спиртах возрастающей крепости и пропитан парапластом по стандартной методике. Срезы толщиной 5 мкм были окрашены гематоксилином.
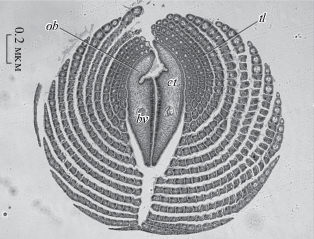
Обтюракулы вестиментифер представляют собой парные выросты переднего конца тела, сдвинутые на спинную сторону и окруженные многочисленными концентрическими щупальцевыми ламеллами. На поперечных срезах обтюракулы выглядят как два симметричных полумесяца, тесно прижатые друг к другу плоскими сторонами (рис. 1). На вентральной стороне обтюракулы образуют острый гребень. На дорсальной стороне обтюракулы расходятся, формируя жёлоб. Поверхность обтюракул одета кутикулой, подостланной однослойным эпидермисом. Толща обтюракул заполнена рыхлой соединительной тканью, состоящей из основного вещества, в толще которого залегают поперечно-ориентированные соединительнотканные клетки. В центре обтюракулярной лопасти проходит целомический канал, по оси которого проходит слепой кровеносный сосуд (рис. 1).
Рис. 1.
Поперечный срез переднего конца тела Oasisia alvinae. Обозначения: bv – кровеносные сосуды, ct – соединительная ткань, ob – обтюракулы, tl – щупальцевые ламеллы.
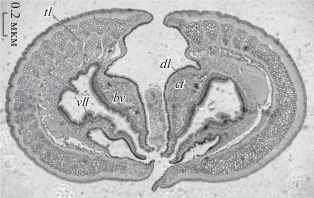
Дорсальные губы Sabellidae представляют собой выросты переднего конца тела, расположенные на дорсальной стороне надо ртом и окруженные парными щупальцевыми ламеллами (рис. 2). В отличие от вестиментифер у сабеллид имеется только одна пара щупальцевых ламелл. На срезах через основание дорсальные губы имеют форму треугольников, направленных острыми концами к вентральной стороне. Снаружи дорсальные губы одеты однослойным ресничным эпидермисом. Ресничный эпидермис более толстый и несёт более густую кайму ресничек на вентро-латеральных сторонах дорсальных губ. Толща дорсальных губ заполнена соединительной тканью, состоящей из плотного основного вещества с соединительнотканными клетками. Вдоль вентро-латеральных сторон дорсальных губ в толще соединительной ткани проходит 11–13 целомических каналов, по оси которых проходят слепые кровеносные сосуды (рис. 2).
Рис. 2.
Поперечный срез переднего конца тела Eudistylia polymorpha. Обозначения: bv – кровеносные сосуды, ct – соединительная ткань, dl – дорсальные губы, tl – щупальцевые ламеллы, vll – вентро-латеральные губы.
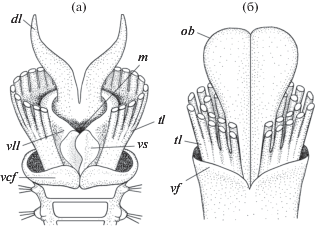
Дорсальные губы сабеллид и обтюракулы вестиментифер могут рассматриваться как гомологи в соответствии с двумя главными критериями гомологии [13]. Первый из них – это критерий сходства положения сравниваемых структурных систем (Kriterium der Lagergleichheit in vergleichbaren Gefugesystem [13]). Дорсальные губы и обтюракулы одинаково расположены по отношению к другим органам. Это парные дорсальные выросты переднего конца тела, окруженные щупальцевыми ламеллами (рис. 3). У полихет семейства Sabellidae имеется только одна пара щупальцевых ламелл (рис. 1). У Vestimentifera щупальцевые ламеллы полимеризованы, они располагаются концентрическими полукругами вокруг обтюракул.
Рис. 3.
Схемы строения переднего конца тела у полихет семейства Sabellidae (а) и сибоглинид подсемейства Vestimentifera, для простоты изображены только две пары щупальцевых ламелл (б). Обозначения: dl – дорсальные губы, m – рот, tl – щупальцевые ламеллы, vcf – вентральная складка воротничка, vf – вестиментальная складка, vll – вентро-латеральные губы, vs –вентральный мешок.
Второй критерий – это критерий специального качества структур (Kriterium der speziellen Qualitat der Strukturen [13]). Дорсальные губы представляют собой вырост, заполненный клеточной соединительной тканью. В толще соединительной ткани по оси дорсальной губы проходят продольные целомические каналы, содержащие кровеносные сосуды. Обтюракулы также представляют собой выросты, заполненные клеточной соединительной тканью. По оси каждой обтюракулярной лопасти имеется продольный целомический канал, внутри которого проходит кровеносный сосуд. Эпидермис дорсальных губ представлен ресничным эпителием, тогда как эпидермис обтюракул одет толстым слоем кутикулы. В толще дорсальной губы проходит много целомических каналов с кровеносными сосудами, тогда как в толще обтюракулярной лопасти проходит единственный центральный целомический канал с единственным кровеносным сосудом.
Различия в строении дорсальных губ и обтюракул могут быть объяснены разными функциями дорсальных губ и обтюракул. Как известно, полихеты-сабеллиды являются фильтраторами, при этом дорсальные губы играют важную роль в сортировке частиц, отфильтрованных щупальцами [14]. Частицы мелкого размера попадают в рот, частицы среднего размера переносятся в вентральный мешок и используются для построения трубки. Крупные частицы, непригодные для питания и построения трубки, транспортируются ресничным эпидермисом вдоль дорсальных губ к их вершинам и отбрасываются [14]. У вестиментифер нет рта и щупальца используются только для газообмена. У холодноводных вестиментифер через щупальца происходит поглощение кислорода и углекислоты (как источника углерода для хемосинтеза), тогда как у гидротермальных вестиментифер через щупальца происходит поглощение кислорода, углекислоты и сероводорода. В связи с этим количество щупальцевых ламелл и общее число щупалец сильно возрастает, достигая у гигантской вестиментиферы Riftia pachyptila 200 тысяч [1]. Обтюракулы выполняют иные функции, чем дорсальные губы сабеллид. У вестиментифер обтюракулы – это опорные структуры для щупальцевого аппарата, выполняющие, кроме того, защитные функции. У многих вестиментифер передний конец обтюракул несёт хитиновые крышечки, которыми закрывается вход в трубку при втягивании обтюракального отдела. В связи с этим обтюракулы вестментифер лишены ресничного покрова, зато они одеты толстой кутикулой, а соединительная ткань имеет плотную консистенцию.
Можно предположить, что при переходе предков вестиментифер к симбиотрофному питанию дорсальные губы не редуцировались, но сменили свою функцию. Если у сабеллид дорсальные губы служили как орган, участвующий в сегрегации отфильтрованных частиц, то у вестментифер обтюракулы стали опорной и защитной структурой. Признание гомологии обтюракул и дорсальных губ служит дополнительным аргументом в пользу сближения сибоглинид с сабеллидами. У других представителей сибоглинид щупальцевый аппарат развит значительно слабее, чем у вестиментифер. Тем не менее у представителей подсемейства Frenulata сохраняется орган, который, вероятно, является гомологом слившихся обтюракул. Это так называемая головная лопасть. В головной лопасти находится целомическая полость и в неё проникают слепые кровеносные сосуды [15]. Относительно слабое развитие щупальцевого аппарата у представителей Monilifera и Osedax обусловили полную редукцию структур, гомологичных обтюракулам.
Список литературы
Карасева Н.П., Римская-Корсакова Н.Н., Галкин С.В., Малахов В.В. Таксономия, географическое и батиметрическое распространение вестиментифер (Annelida, Siboglinidae) // Зоологич. журн. 2016. Т. 95. № 6. С. 624–659. https://doi.org/10.7868/S004451341606012X
Rousset V., Rouse G.W., Siddall M.E., Tillier A., Pleijel F. 2004. The Phylogenetic Position of Siboglinidae (Annelida) Inferred from 18S rRNA, 28S rRNA and Morphological Data // Cladistics. 2004. V. 20. P. 518–533.
Struck T.H., Paul C., Hill N., Hartmann S., Hösel C., Kube M., Lieb B, Meyer A, Tiedemann R., Purschke G., Bleidorn C. Phylogenomic Analyses Unravel Annelid Evolution. Nature. 2011. V. 471. P. 95–98.
Andrade S.C.S., Novo M., Kawauchi G.Y., Worsaae K., Pleijel F., Giribet G., Rouse G.W. Articulating “Archiannelids”: Phylogenomics and Annelid Relationships, with Emphasis on Meiofaunal Taxa // Mol. Biol. Evol. 2015. V. 32. № 11. P. 2860–2875. https://doi.org/10.1093/molbev/msv157
Uschakow P. Eine neue Form aus der Familie Sabellidae (Polychaeta) // Zoologischer Anzeiger. 1933. 104: 205–208.
Rouse G.W., Pleijel F. Polychaetes. Oxford Univ. Press, Oxford: 2001. 1354 p.
Schulze A. Phylogeny of Vestimentifera (Siboglinidae, Annelida) inferred from morphology // Zoologica Scripta. 2003. 32, 321–342.
Gardiner S.L., Jones M.L. Vestimentifera. In: Microscopic anatomy of Invertebrates. Vol. 12. Onychophora, Chilopoda, and lesser Protostomata. Wiley. 1993. P. 371–460.
Малахов В.В., Галкин С.В. Вестиментиферы – бескишечные беспозвоночные морских глубин. М.: КМК, 1998. 204 с.
Webb M. Lamellibrachia barhami, gen. nov., sp. nov., (Pogonophora), from the northeast Pacific//Bull. Mar. Sci. 1969. V. 19. P. 18–47.
Иванов А.В. О морфологической природе обтюракулов у Pogonophora // ДАН. 1989. Т. 308. № 3. С. 758–759.
Southward E.S. Development of the Gut and Segmentation of Newly Settled Stages of Ridgeia (Vestimentifera): Implications for Relationship Between Vestimentifera and Pogonophora // J. Mar. Biol. Assoc. U.K. 1988. V. 68. P. 465–487.
Remane A. Die Grundlagen des natürlichen Systems, der vergleichenden Anatomie und der Phylogenetik. 2 Auflage. Leipzig: Geest & Portig K.-G. 1956. 364 S.
Nicol B.A.T. The feeding mechanism, formation of the tube and physiology of digestions in Sabella pavonina. Trans. R. Soc. Edinburgh 1931. T. 56. P. 537–698.
Иванов А.В. Погонофоры // Фауна СССР. Новая сер. М.; Л: Изд-во АН СССР, 1960. № 75. 271 с.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни