

Доклады Российской академии наук. Науки о жизни, 2020, T. 490, № 1, стр. 67-71
ФИЛОГЕНЕТИЧЕСКИЕ СВЯЗИ НАЛИМА (Lota lota L., 1758) ВОЛЖСКО-КАМСКОГО РЕЧНОГО БАССЕЙНА ПО ДАННЫМ АНАЛИЗА МИТОХОНДРИАЛЬНЫХ МАРКЁРОВ
Л. Э. Ялковская 1, *, Ю. Я. Хрунык 2, М. А. Крохалева 1, член-корреспондент РАН В. Д. Богданов 1, П. А. Сибиряков 1, С. Б. Ракитин 1, А. В. Бородин 1
1 Институт экологии растений и животных Уральского отделения Российской Академии наук
Екатеринбург, Россия
2 Уральский федеральный университет им. Б.Н. Ельцина
Екатеринбург, Россия
* E-mail: lida@ipae.uran.ru
Поступила в редакцию 17.05.2019
После доработки 17.05.2019
Принята к публикации 17.05.2019
Аннотация
Впервые проведена реконструкция филогенетических связей налима (Lota lota L., 1758) Волжско-Камского речного бассейна. Материалом послужили последовательности гена цитохрома b и контрольного региона мтДНК 44 образцов из притоков Волги – р. Кама и р. Межевая Утка. В ходе исследования для обоих маркёров описаны новые гаплотипы. Результаты филогенетической реконструкции как по цитохрому b, так и контрольному региону мтДНК не противоречат современным представлениям о филогенетической структуре вида и указывают на принадлежность налима Волжско-Камского речного бассейна к Евразийской гаплогруппе. Согласно полученным данным, Волжско-Камский бассейн мог играть важную роль в формировании генетического разнообразия налима в Европе, а в определенные периоды служить коридором, соединяющим речные системы европейской и азиатской частей видового ареала.
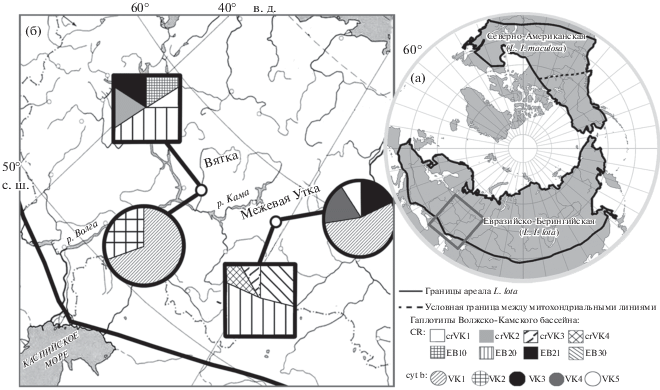
Генетические исследования широко распространенных видов на основе анализа митохондриальных маркёров успешно используются при решении вопросов внутривидовой дифференциации, систематики и филогении, а также для реконструкции эволюционной истории отдельных таксонов и биомов в целом. Налим (Lota lota L., 1758) – единственный пресноводный представитель семейства Gadidae. Циркумполярный ареал вида представлен реками и озерами Евразии и Северной Америки вплоть до 40° с.ш. (рис. 1). Исследования генетической структуры L. lota с использованием гена цитохрома b (cyt b) и контрольного региона (CR) мтДНК охватывают территории обоих континентов [1–6]. Показано существование двух крупных митохондриальных линий, соответствующих двум подвидам. Северо-Американская линия (L. l. maculosa) занимает юго-восточную часть ареала вида в Северной Америке, а Евразийско-Берингийская (L. l. lota) – северо-западную часть Северной Америки и территорию всей Евразии, и, собственно, она имеет циркумполярный ареал (рис. 1).
Рис. 1.
Ареал L. lota и условная граница между митохондриальными линиями (а), места сбора образцов из Волжско-Камского речного бассейна и распределение гаплотипов cyt b и CR мтДНК (б).
Анализ генетического разнообразия и демографической истории Евразийско-Берингийской линии свидетельствует о формировании границ её современной области распространения в постледниковый период. Согласно филогенетическим реконструкциям, в пределах линии дифференцируются группы гаплотипов, распространение которых ограничено периферийными частями ареала, а их базальные гаплотипы являются производными от гаплотипов широко распространенной Евразийской группы, занимающей территорию Западной и Северной Европы, а также Западной и Восточной Сибири [1–3, 6]. Формирование Евразийской группы связывают с существованием нескольких рефугиумов. Для Европы такое значение мог иметь Дунайский бассейн [1–3]. Но, как показали наши исследования генетического разнообразия налима Обь-Тазовского бассейна [6], реконструкции эволюционной истории налима нельзя считать обоснованными без учета возможного вклада крупных речных систем центральной и восточной Евразии, данные по которым практически отсутствуют. Так, в генетических исследованиях налима не представлен крупнейший в Европе Волжско-Камский речной бассейн, который впадает в бессточное (внутреннее) Каспийское море и в формировании которого участвуют как европейские, так азиатские реки. Согласно геологической истории бассейна, в его недавнем прошлом были периоды соединения как с европейскими, так и с сибирскими гидрологическими системами [7]. В этой связи нами были проведены исследования филогенетических связей налима Волжско-Камского речного бассейна на основе анализа двух митохондриальных маркёров cyt b и CR.
В работе использованы образцы мышечной ткани, зафиксированной в 96%-м этаноле, от 44 особей налима из притоков Волги второго и третьего порядка – р. Вятка (57°38' с.ш. 59°04' в.д.) и р. Межевая Утка (56°07' с.ш. 51°20' в.д.) соответственно (рис. 1). Выделение ДНК, амплификация (условия ПЦР одинаковы для обоих маркёров), секвенирование фрагментов мтДНК, содержащих cyt b и CR, обработка последовательностей, оценка генетического разнообразия и филогенетические реконструкции проведены с применением методов, подробно описанных ранее [6]. Для амплификации cyt b использовали праймеры L14299Gm и H14942Gm [1], CR – LProF и 12S5R [6].
Для филогенетических построений использованы данные GenBank. Для cyt b – 39 гаплотипов (AY226383-444 [1], KM201364 [8], KM363244 [9], KC44053, KU244689-92, KT327178 (неопубликованные данные)), большинство из которых представлены последовательностями длиной 1063 пн: фрагмент от 5'-конца – 567 пн и фрагмент от 3'- конца – 496 пн [1]. Для CR – 104 гаплотипа (AY656840-915 [2], DQ630963-1411 [3], EU873154-65 [4], KX017626-48 [6], KM201364 [8], KM363244 [9], KJ381204 [10], KC44053, EF191381-84, EF191387 (неопубликованные данные) длиной 409 п.н. от 5'-конца, что позволило включить в анализ максимальный объём данных, имеющихся в GenBank.
Полные последовательности cyt b (1140 пн) налима Волжско-Камского бассейна получены от 21 особи: 10 особей – р. Вятка, 11 – р. Межевая Утка. Описано пять гаплотипов (депонированы в базе данных GenBank (NCBI) под номерами MN708969–MN708973), четыре из которых являются новыми. Гаплотип VK1 выявлен у большинства особей в обеих реках (семь и шесть особей – р. Вятка и р. Межевая Утка соответственно), VK2 (три особи) обнаружен только в р. Вятка, VK3–VK5 – в р. Межевая Утка (VK3 – две особи, VK4 – две особи, VK5 – одна особь) (рис. 1).
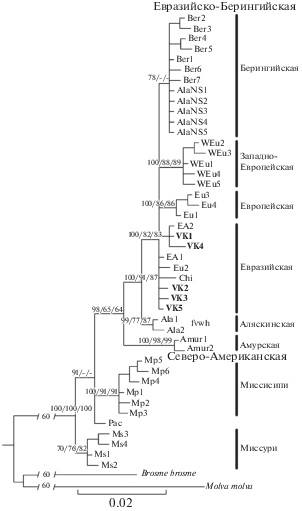
Филогенетические реконструкции по данным cyt b (рис. 2) показали, что гаплотипы налима Волжско-Камского бассейна, как и ожидалось согласно представлениям о распространении митохондриальных линий, относятся к Евразийско-Берингийской линии, структура которой в целом соответствует предложенным ранее схемам [1, 5]. Гаплотипы Волжско-Камского бассейна не образуют отдельной группы и относятся к Евразийской гаплогруппе, распространенной от Швеции, Финляндии, Онежского озера до оз. Байкал и бассейна р. Лены (рис. 2).
Рис. 2.
Филогенетическое дерево (метод Байесова анализа (BI)) гаплотипов cyt b (1063 пн) L. lota. Шрифтом выделены гаплотипы, обнаруженные в Волжско-Камском речном бассейне. Над ветвями даны вероятности поддержки гаплогрупп (BI > 60/метод максимального правдоподобия (ML) > 50/метод ближайшего соседа (NJ) > 50), прочерком обозначена низкая поддержка при данном методе построения. В разрывах на ветвях указано сокращение их длины (%).
Для 23 особей налима Волжско-Камского бассейна (12 особей – р. Вятка, 11 – р. Межевая Утка) секвенированы полные последовательности CR (804 п.н.), описано 11 гаплотипов, среди которых восемь являются новыми (депонированы в базе данных GenBank (NCBI) под номерами MN685694–MN685704). Использование при филогенетических реконструкциях только небольшого участка CR (409 пн) привело к уменьшению общего числа гаплотипов с 11 до 8: четыре гаплотипа crVK1-crVK4 новые; а четыре гаплотипа EB10, EB20, EB21, EB30 описаны ранее в пределах Евразийско-Берингийской линии [2]. Гаплотип EB20 обнаружен в обеих реках (пять и шесть особей – р. Вятка и р. Межевая Утка соответственно), ЕВ10 (две особи), EB21 (одна особь), crVK1 (одна особь) и crVK2 (одна особь) – только в р. Вятка, а EB30 (три особи), crVK3 (одна особь) и crVK4 (одна особь) – только в р. Межевая Утка (рис. 1).
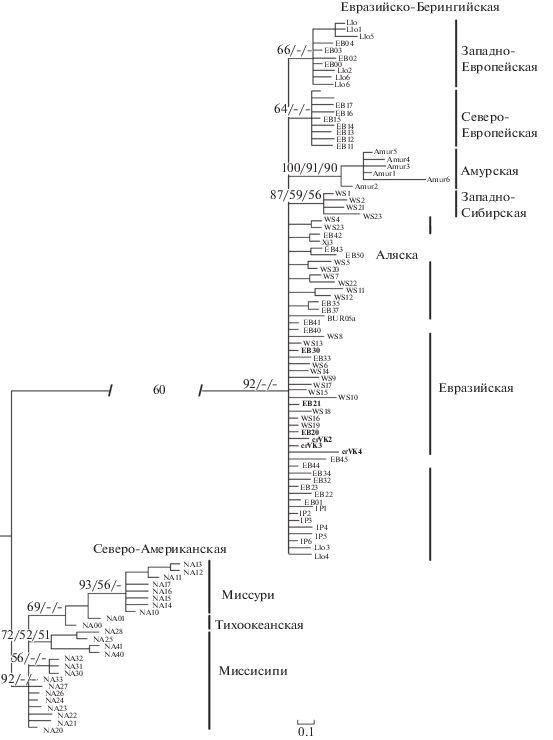
Результаты филогенетической реконструкции на основе последовательностей CR (рис. 3) согласуются с данными, полученными по cyt b, и не противоречат имеющимся представлениям о филогеографической структуре вида [2, 5, 6]. Обнаруженные нами новые гаплотипы (crVK1–crVK4) не образуют самостоятельной клады. Среди описанных ранее три гаплотипа EB20, EB21, EB30 принадлежат Евразийской гаплогруппе и имеют широкое распространение: EB20 – Дунайский речной бассейн, Онежское озеро и р. Кюрёнйоки (Финляндия); EB21 – Дунайский речной бассейн, р. Висла, р. Обь; EB30 – Центральная Европа, Западная и Восточная Сибирь, считается одним из базальных гаплотипов для всей Евразийско-Берингийской линии [2]. Четвертый гаплотип EB10, обнаруженный в р. Вятка, ранее описан на территории Скандинавии [2] и рассматривается как базальный для Северо-Европейской группы, в которую также оказался включён один из вятских гаплотипов crVK1 (рис. 3).
Рис. 3.
Филогенетическое дерево (метод BI) гаплотипов CR (409 пн) L. lota. Шрифтом выделены гаплотипы, обнаруженные в Волжско-Камском речном бассейне. Над ветвями даны вероятности поддержки гаплогрупп BI > 60/ML > > 50/NJ > 50, прочерком обозначена низкая поддержка при данном методе построения. В разрывах на ветвях указано сокращение их длины (%).
Относительно высокое гаплотипическое разнообразие (cyt b – h = 0.605, CR – h = 0.759) при низких значениях нуклеотидного разнообразия (cyt b – π(%) = 0.108, CR – π(%) = 0.424) налима Волжско-Камского речного бассейна указывает на его недавнее расселение в пределах современной акватории бассейна, что согласуется с представлениями о формировании генетической дифференциации Евразийско-Берингийской линии в целом.
Таким образом, анализ филогенетических связей налима Волжско-Камского речного бассейна позволяет предположить, что он, как и Дунайский бассейн, играл важную роль в формировании генетического разнообразия вида в Европе, а в определенные периоды мог служить своеобразным коридором, соединяющим речные системы европейской и азиатской частей видового ареала.
Список литературы
Van Houdt J.K.J., Hellemans B., Volckaert F.A.M. Phylogenetic Relationships among Palearctic and Nearctic Burbot (Lota lota): Pleistocene Extinctions and Recolonization // Molecular Phylogenetics and Evolution. 2003. V. 29. P. 599–612.
Van Houdt J.K.J., De Cleyn L., Perretti A., et al. A Mitogenic View on the Evolutionary History of the Holarctic Freshwater Gadoid, Burbot (Lota lota) // Molecular Ecology. 2005. V. 14. P. 2445–2457.
Barluenga M., Sanetra M., Meyer A. Genetic Admixture of Burbot (Teleostei: Lota lota) in Lake Constance from Two European Glacial Refugia // Molecular Ecology. 2006. V. 15. P. 3583–3600.
Elmer K.R., Van Houdt J.K.J., Meyer A., et al. Population Genetic Structure of North American Burbot (Lota lota maculosa) across the Nearctic and at Its Contact Zone with Eurasian Burbot (Lota lota lota) // Canadian J. of Fisheries and Aquatic Sciences. 2008. V. 65. № 11. P. 2412–2426.
Fang H., Zhang J., Song N., et al. Population Genetic Structure and Geographical Differentiation of Burbot (Lota lota) in China // Russian J. Genetics. 2013. V. 49. № 10. P. 1047–1056.
Хрунык Ю.Я., Богданов В.Д., Ялковская Л.Э. и др. Генетическое разнообразие налима (Lota lota L., 1758) Западной Сибири по результатам анализа полиморфизма контрольного региона мтДНК // Генетика. 2017. Т. 53. № 2. С. 214–222.
Застрожнов А.С., Шкатова В.К., Минина Е.А. и др. Карта четвертичных образований масштаба 1 : 2 500 000 территории Российской Федерации. Пояснительная записка. Минприроды России, Роснедра, ФГУП “ВСЕГЕИ”, ФГУП “ВНИИОкеангеология”, 2010. 220 с.
Zhang N., Song N., Gao T. The Complete Mitochondrial Genome of Lota lota (Gadiformes: Gadidae) // Mitochondrial DNA. 2016. V. 27. № 2. P. 1437–1438.
Lu Z., Zhang N., Song N., et al. The Complete Mitochondrial Genome of Lota lota (Gadiformes: Gadidae) from the Burqin River in China // Mitochondrial DNA. 2016. V. 27. № 3. P. 1662–1663.
Worthington T.A., Van Houdt J.K.J., Hull J.M., et al. Thermal Refugia and the Survival of Species in Changing Environments: New Evidence from a Nationally Extinct Freshwater Fish // Ecology of Freshwater Fish. 2017. V. 26. P. 415–423.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни