

Доклады Российской академии наук. Науки о жизни, 2021, T. 500, № 1, стр. 391-401
КРУПНЫЙ ДИКОБРАЗ Hystrix refossa (RODENTIA, HYSTRICIDAE) ИЗ РАННЕПЛЕЙСТОЦЕНОВОГО МЕСТОНАХОЖДЕНИЯ ТАВРИДА В КРЫМУ
Академик РАН А. В. Лопатин 1, *
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 27.04.2021
После доработки 31.05.2021
Принята к публикации 31.05.2021
Аннотация
Остатки крупных дикобразов (неполный череп, черепные и нижнечелюстные фрагменты, изолированные зубы и кости посткраниального скелета) из раннеплейстоценового пещерного местонахождения Таврида в центральном Крыму (Белогорский район, пос. Зуя) отнесены к виду Hystrix (Hystrix) refossa Gervais, 1852, ранее из Крыма не известному. На основании стадий прорезывания, смены и стирания нижних зубов установлено присутствие в выборке особей шести возрастных групп – от ювенильной до сенильной. Одна нижнечелюстная кость демонстрирует прижизненную утрату щечных зубов, предположительно связанную с аномальным недоразвитием их корней и альвеол.
В 2019 г. мной были описаны остатки мелкого дикобраза Hystrix (Acanthion) vinogradovi Argyropulo, 1941, обнаруженные годом ранее в раннеплейстоценовом пещерном местонахождении Таврида в центральном Крыму (Белогорский район, пос. Зуя) [1]. Сборы 2020 г. в южном коридоре пещеры Таврида пополнили коллекцию ископаемых позвоночных новыми материалами по H. (A.) vinogradovi и многочисленными находками остатков еще одного вымершего вида дикобразов – крупного Hystrix (Hystrix) refossa Gervais, 1852. Этот вид, широко распространенный в плейстоцене Евразии [2–4], в Крыму ранее известен не был. Указание [4, табл. 1] на находку зубов H. refossa в нижнем плейстоцене м. Тарханкут (оригинальное определение [5] – Hystrix sp.) ошибочно – судя по мелким размерам [5], эти остатки принадлежат H. (A.) vinogradovi [1].
Таблица 1.
Размеры (в мм) черепа Hystrix refossa Gervais, 1852 из нижнего плейстоцена пещеры Таврида, Крым (dex – правый; sin – левый)
| Параметр | Экземпляр ПИН, № | ||
|---|---|---|---|
| 5644/555 | 5644/500 | 5644/550 | |
| Длина лобных костей | 55.9 | – | – |
| Длина теменных костей (от лобно-теменного шва до заднего края затылочного гребня) | 36.0 | 30.5 | 42.4 |
| Длина сагиттального гребня | 20.8 | 21.0 | 23.7 |
| Расстояние от заднего края твердого неба до затылочных мыщелков | 78.0 | – | – |
| Расстояние от переднего края P4 до заднего края твердого неба | 31.0 | – | – |
| Альвеолярная длина P4–M3 | 38.7 (dex); 39.0 (sin) | – | – |
| Окклюзиальная длина P4–M3 | 37.7 (dex); 37.8 (sin) | – | – |
| Расстояние между зубными рядами на уровне P4 | 13.6 | – | – |
| Расстояние между зубными рядами на уровне M3 | 13.1 | – | – |
| Высота черепа над M3 | 79.1 | – | – |
| Высота затылка по средней линии | 52.6 | 51.9 | 55.7 |
| Наибольшая ширина затылка выше foramen magnum | 61.5 | 64.0 | 68.9 |
| Расстояние между отверстиями наружных слуховых проходов | 64.3 | 64.5 | 71.0 |
| Наибольшая ширина затылочных мыщелков (между наружными краями) | 38.3 | – | 41.7 |
| Наибольшая ширина foramen magnum | 18.0 | – | 19.5 |
Описанный ниже материал по H. refossa из пещеры Таврида хранится в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН) в Москве. Терминология зубных структур по [1], обозначения стадий стирания зубов по [6, 7]. Размеры в мм. Обозначения: AL – альвеолярная длина, AW – альвеолярная ширина, DL – длина диастемы, L – наибольшая длина, LD – продольный диаметр, OL – окклюзиальная длина, OW – окклюзиальная ширина, TD – поперечный диаметр, W – наибольшая ширина; n – число экземпляров, μ – среднее значение; dex – правый, sin – левый.
Отряд Rodentia Bowdich, 1821
Семейство Hystricidae Fischer von Waldheim, 1817
Подсемейство Hystricinae Fischer von Waldheim, 1817
Род Hystrix Linnaeus, 1758
Подрод Hystrix Linnaeus, 1758
Hystrix (Hystrix) refossa Gervais, 1852
Hystrix refossa: [8, с. 8, табл. 48, фиг. 11].
Голотип – Лаборатория палеонтологии Национального музея естественной истории, Париж; фрагмент правой нижнечелюстной кости с P4 и альвеолами M1 [2, табл. I, фиг. 1a, 1b]; Франция, Перье, Этуэр (Les Etouaires); верхний плиоцен, MN16b.
Рис. 1.
Hystrix refossa Gervais, 1852: а–д – экз. ПИН, № 5644/555, неполный череп: а – с дорсальной стороны, б – с вентральной стороны, в – с правой латеральной стороны, г – с затылочной стороны, д – верхние щечные зубные ряды; е, ж – экз. ПИН, № 5644/502, изолированный нестертый левый P4: е – с окклюзиальной стороны, ж – с лингвальной стороны; Крым, пещера Таврида; нижний плейстоцен.

Описание (рис. 1–4). Дикобраз крупных размеров с гипсодонтными зубами. Череп с умеренно высокой выпуклой крышей лобной области, с относительно высоким и широким затылком (экз. ПИН, № 5644/555; рис. 1, табл. 1). Носовые кости умеренно длинные, достигают уровня переднего корня скуловых дуг (сохранилась область носо-лобных швов). Передний корень скуловой дуги расположен антеролатеральнее P4. Лобные кости длинные, лобно-теменные швы находятся намного каудальнее заднего корня скуловых дуг. Теменная область широкая. Сагиттальный гребень высокий, тонкий, сравнительно короткий. Имеется короткая и широкая треугольная площадка позади лобных костей, образованная мощными теменными гребнями, косо расходящимися от переднего конца сагиттального гребня. Затылочный гребень резкий. Затылочные мыщелки широкие и низкие, длинная ось их сочленовной поверхности субвертикальная. Заднее отверстие подглазничного канала находится на уровне передней части M2. На небной части верхнечелюстных костей перед P4 имеются большие округлые впадины для m. buccinator [см. 9]. Передний край небно-верхнечелюстного шва расположен на уровне средней части M2. Задний край твердого неба находится на уровне передней части M3.
Рис. 2.
Hystrix refossa Gervais, 1852: а – экз. ПИН, № 5644/503, парные нижнечелюстные кости с I1 и P4–M3 с окклюзиальной стороны; б, в – экз. ПИН, № 5644/504, парные нижнечелюстные кости с I1 и P4–M3: б – с окклюзиальной стороны, в – левая нижнечелюстная кость с лабиальной стороны; г – экз. ПИН, № 5644/524, левая нижнечелюстная кость с I1 и M1–M2 с лабиальной стороны; д–ж – экз. ПИН, № 5644/525, правая нижнечелюстная кость с I1 и P4–M3: д – с лабиальной стороны, е – с окклюзиальной стороны, ж – с лингвальной стороны; з – экз. ПИН, № 5644/509, правая нижнечелюстная кость с P4–M2 с лабиальной стороны; и – экз. ПИН, № 5644/531, левая нижнечелюстная кость с I1 с лабиальной стороны; Крым, пещера Таврида; нижний плейстоцен.

Рис. 3.
Hystrix refossa Gervais, 1852, нижние щечные зубные ряды разных стадий стирания с окклюзиальной стороны: а – экз. ПИН, № 5644/524, левые M1–M2 (стадия A/B1), альвеолы DP4 и M3; б – экз. ПИН, № 5644/526, левые DP4–M3 (стадия B1/B2); в – экз. ПИН, № 5644/527, правые M1–M3 (стадия B1/B2), альвеолы DP4; г – экз. ПИН, № 5644/534, правые M1–M3 (стадия B1/B2), альвеолы DP4; д, е – экз. ПИН, № 5644/503, левые M1–M3 (P4 внутри альвеолы) и правые P4–M3 (стадия C); ж – экз. ПИН, № 5644/535, правые P4–M3 (стадия C/D1); з – экз. ПИН, № 5644/512, правые P4–M3 (стадия D1); и, к – экз. ПИН, № 5644/504, левые и правые P4–M3 (стадия D2); л – экз. ПИН, № 5644/522, левые P4 и M2–M3 (стадия D2/E), альвеолы M1; м – экз. ПИН, № 5644/525, правые P4–M3 (стадия D2/E); н – экз. ПИН, № 5644/509, правые P4–M2 (стадия E), альвеола M3; о – экз. ПИН, № 5644/521, левые P4–M3 (стадия E); п – экз. ПИН, № 5644/523, правые M1–M3 (стадия E), альвеолы P4; р – экз. ПИН, № 5644/531, частично заросшие альвеолы левых P4–M3; Крым, пещера Таврида; нижний плейстоцен.

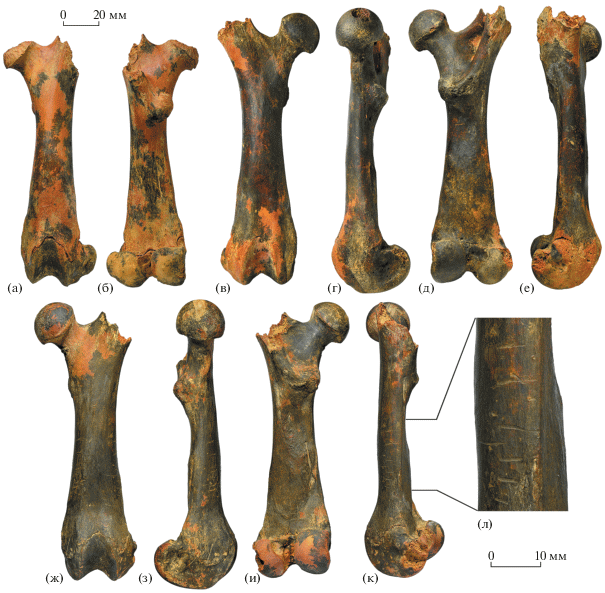
Рис. 4.
Hystrix refossa Gervais, 1852: а, б – экз. ПИН, № 5644/508, левая бедренная кость: а – с передней стороны, б – с задней стороны; в–е – экз. ПИН, № 5644/510, правая бедренная кость: в – с передней стороны, г – с медиальной стороны, д – с задней стороны, е – с латеральной стороны; ж–л – экз. ПИН, № 5644/511, левая бедренная кость: ж – с передней стороны, з – с медиальной стороны, и – с задней стороны, к – с латеральной стороны, л – то же, следы (ямки и бороздки) зубов хищника; Крым, пещера Таврида; нижний плейстоцен.
Верхние щечные зубные ряды параллельны друг другу (рис. 1б), по длине P4 = M2 > M1 > M3. P4 в нестертом (экз. ПИН, № 5644/502) и слабо стертом (экз. ПИН, № 5644/555) состоянии имеет замкнутые парафлексию и постфлексию и широко открытые переднюю мезофлексию и гипофлексию. Антерофлексия (складка, выходящая на переднюю сторону) и задняя мезофлексия открыты на нестертом зубе. Передняя мезофлексия лингвальным краем касается эмалевой стенки гипофлексии либо объединяется с этой складкой. Постфоссетта и задняя мезофоссетта соединены с лингвальной мезофоссеттой и образуют вместе с ней единую фоссетту подковообразной формы. На нестертом P4 лингвальная эмалевая высота коронки превосходит длину жевательной поверхности в 2.3 раза, а высота гипострии составляет менее 20% от лингвальной эмалевой высоты коронки.
На значительно стертых М1 (стадия стирания D) либо все складки замкнутые, либо остаются открытыми гипофлексия и передняя мезофлексия при очень коротких бороздках на соответствующих боковых сторонах (экз. ПИН, № 5644/555). Постфоссетта обособлена от более передней по положению загнутой назад фоссетты, объединяющей заднюю мезофоссетту и лингвальную мезофоссетту. На М2 сходного строения гипофлексия открытая, передняя мезофлексия имеет узкий выход или замкнутая, эмалевые стенки задней мезофлексии сомкнуты у выхода, но при этом сохраняется четкая лабиальная бороздка (задняя мезострия). Парафоссетта слегка изогнута вперед и сужена в средней части. Для M3 характерно наличие замкнутых гипофоссетты, парафоссетты и постфоссетты при открытых передней мезофлексии и задней мезофлексии. Парафоссетта сужена в средней части. Задняя мезофлексия соединена с лингвальной мезофоссеттой, которая дистально объединена с постфоссеттой.
Нижнечелюстная кость массивная, но относительно низкая (рис. 2). Диастема по длине от 64–75% (у молодых особей) до 81–92% длины ряда щечных зубов (табл. 2). Сочленовный отросток немного выше венечного отростка. Венечный отросток с сильно утолщенной вершиной. Крупное подбородочное отверстие расположено у окончания диастемы, близко к дорсальному краю кости. Нижний массетерный гребень высокий, спереди оканчивается под M1, у некоторых сенильных особей с сильно стертыми щечными зубами достигает середины P4 (экз. ПИН, №№ 5644/521, 522). Симфиз простирается назад до уровня задней части нижнего премоляра (DP4/P4). Альвеолярный бугор нижнего резца умеренно вздутый.
Таблица 2.
Размеры (в мм) нижних зубов и нижнечелюстной диастемы Hystrix refossa Gervais, 1852 из нижнего плейстоцена пещеры Таврида, Крым (AL – альвеолярная длина; DL – длина диастемы; LD – продольный диаметр; OL – окклюзиальная длина; OW – окклюзиальная ширина; TD – поперечный диаметр; dex – правая кость; sin – левая кость)
| Экземпляр ПИН, № (стадия стирания) | Зубной ряд и диастема | Зубы (резец: LD × TD; щечные зубы: OL × OW) |
|---|---|---|
| 5644/524 (A/B1) | DL: 30.0 | M1 – 11.8 × 9.1 |
| 5644/526 (B1/B2) | AL DP4–M3: 41.3; DL: 29.3 | DP4 – 9.9 × 7.1; M1 – 10.6 × 7.8; M2 – 10.7 × 7.0 |
| 5644/527 (B1/B2) | AL DP4–M3: 41.6 | M1 – 10.9 × 7.6; M2 – 10.8 × 7.2 |
| 5644/534 (B1/B2) | AL DP4–M3: 43.5; DL: 28.0 | M1 – 11.8 × 8.5; M2 – 10.0 × 6.5 |
| 5644/536 (B1/B2) | – | M1 – 11.8 × 8.0 |
| 5644/503 (C) | AL DP4–M3: 44.0 (dex), 40.1 (sin); DL: 32.9 (dex), 34.5 (sin) | I1: 7.5 × 6.6 (dex), 7.6 × 6.7 (sin); M1: 10.3 × 8.1 (dex), 10.0 × 8.2 (sin); M2: 11.0 × 8.5 (dex), 11.0 × 8.1 (sin); M3: 9.0 × 6.5 (dex) |
| 5644/535 (C/D1) | AL P4–M3: 44.0; OL P4–M3: 41.7; DL: 33.5 | I1 – 8.6 × ?; P4 – 11.0 × 7.2; M1 – 10.0 × 8.1; M2 – 11.0 × 8.2; M3 – 9.5 × 6.1 |
| 5644/512 (D1) | AL P4–M3: 46.2; OL P4–M3: 43.0; DL: 33.5 | P4 – 12.0 × 8.8; M1 – 10.5 × 9.2; M2 – 11.1 × 9.6; M3 – 9.5 × 8.0 |
| 5644/504 (D2) | AL P4–M3: 46.5 (dex), 45.0 (sin); OL P4–M3: 42.7 (dex), 42.0 (sin); DL: 38.2 (dex), 39.4 (sin) | I1 – 8.0 × 7.0 (dex, sin); P4 – 11.5 × 8.8 (dex), 11.5 × 8.6 (sin); M1: 10.2 × 8.0 (dex), 10.3 × 8.0 (sin); M2: 11.5 × 8.8 (dex), 11.7 × 9.2 (sin); M3: 10.4 × 7.8 (dex), 10.0 × 7.9 (sin) |
| 5644/522 (D2/E) | AL P4–M3: 46.0; OL P4–M3: 42.4 | P4 – 13.0 × 9.8; M2 – 11.0 × 11.0; M3 – 9.7 × 9.6 |
| 5644/525 (D2/E) | AL P4–M3: 39.2; OL P4–M3: 38.5; DL: 36.2 | P4 – 11.5 × 8.6; M1 – 7.9 × 8.7; M2 – 9.9 × 8.8; M3 – 9.0 × 7.1 |
| 5644/509 (E) | AL P4–M3: 42.9; DL: 38.5 | P4 – 13.4 × 10.0; M1 – 8.2 × 9.4; M2 – 10.6 × 9.7 |
| 5644/521 (E) | AL P4–M3: 44.5; OL P4–M3: 42.4; DL: 35.9 | P4 – 12.8 × 9.0; M1 – 9.7 × 8.7; M2 – 11.0 × 10.0; M3 – 9.8 × 8.0 |
| 5644/523 (E) | AL P4–M3: 45.0; DL: 28.5 | I1 – 8.6 × 7.3; M1 – 10.2 × 8.6; M2 – 11.5 × 9.0; M3 – 10.0 × 7.9 |
| 5644/531 (без щечных зубов) | AL P4–M3: ~38.8 | I1 – 8.8 × 7.6 |
DP4 сравнительно небольшой, с узкой передней частью. Р4 крупный, массивный, удлиненный. M1 крупный, но в стертом состоянии уступает по окклюзиальной длине и ширине другим нижним щечным зубам. M2 крупнее прочих моляров. M3 относительно узкий в задней части.
У наиболее молодой особи (экз. ПИН, № 5644/524) жевательная поверхность недавно прорезавшегося M2 ниже таковой M1 (рис. 2г). M1 слабо стертый, M2 в состоянии начального стирания (стадия A/B1). На молярах с лингвальной стороны открыты три складки – передний мезофлексид, задний мезофлексид и постерофлексид (рис. 3а).
На стадии B1/B2 (экз. ПИН, №№ 5644/526, 527, 534, 536) DP4 сильно стертый, M1 и M2 слабо стертые, M3 в состоянии прорезывания, едва выступает над альвеолой (рис. 3б–3г). На DP4 (экз. ПИН, № 5644/526) шесть маленьких эмалевых островков (по три на передней и задней долях жевательной поверхности). На M1 передний мезофлексид имеет узкий выход или замкнут, изолирован или соединен с антерофоссеттидом в единую складку подковообразной формы; лабиальный мезофоссеттид имеет обособленное положение; задний мезофлексид и гипофлексид разобщены; постерофлексид имеет узкий выход или замкнут. На M2 открытый передний мезофлексид в лабиальной части сообщается с круговым фоссеттидом, полностью окружающим центральный конид; задний мезофлексид с широким выходом, лабиально может быть связан с круговым фоссеттидом (экз. ПИН, №№ 5644/527, 534); широкий постерофлексид объединен с гипофлексидом.
На экз. ПИН, № 5644/503 (стадия C) в левом и правом зубных рядах DP4 выпали, P4 находятся внутри альвеол, правый P4 в состоянии начального прорезывания (рис. 3д, 3е). M1 сильно стертые, с пятью эмалевыми островками и гипофлексидом. M2 умеренно стертые, имеют три незамкнутые складки – глубокий гипофлексид, хорошо развитый задний мезофлексид и постерофлексид с узким устьем. Коронка M3 в состоянии прорезывания возвышается над лабиальной стенкой альвеолы, но ниже ее лингвальной стенки.
На экз. ПИН, № 5644/535 (стадия C/D1) P4 полностью прорезавшийся, практически не стертый (рис. 3ж). M1 с пятью замкнутыми долинками и глубоким гипофлексидом. Передний мезофоссеттид слит с лабиальным мезофоссеттидом в единую долинку. На M2 открыты гипофлексид и задний мезофлексид, постерофоссеттид замкнут. В передней половине жевательной поверхности находятся две крупных замкнутых долинки и маленький эмалевый островок. M3 в состоянии начального стирания.
Стадия D1 (экз. ПИН, № 5644/512) характеризуется замкнутыми долинками при сохранении глубоких гипофлексидов на P4–M2. На P4 четыре обширных субпоперечных долинки, на M1 – пять сравнительно небольших долинок, на M2 – три мелких эмалевых островка в антеролингвальной части, косая длинная долинка постеролабиальнее от них и две широкие субпоперечные долинки в задней части жевательной поверхности (рис. 3з). На M3 узкие выходы имеют задний мезофлесид и постерофлексид, гипофлексид глубокий.
На экз. ПИН, № 5644/504 левый и правый нижние зубные ряды демонстрируют стадию стирания D2 (рис. 3и, 3к). На P4 четыре эмалевых долинки и незамкнутый гипофлексид; на M1 – пять эмалевых складок (на левом зубе замкнутый гипофоссеттид, на правом зубе выположенный гипофлексид с узким устьем, совмещенным с короткой бороздкой на лабиальной стороне); на M2 – четыре-пять эмалевых островков и глубокий гипофлексид; на M3 – пять эмалевых островков и гипофлексид.
На стадии D2/E (экз. ПИН, №№ 5644/522, 525) гипофлексид сохраняется на P4 и M3 (или только на P4), но замкнут в гипофоссеттид на M1 и M2 (рис. 3л, 3м).
На заключительной стадии E (экз. ПИН, №№ 5644/509, 521, 523) все эмалевые долинки (обычно их пять-шесть) замкнутые и имеют небольшие размеры (рис. 3н–3п).
Кости посткраниального скелета массивные, крупные. Бедренная кость (рис. 4) относительно широкая в диафизе, с крупной головкой, характерной относительно короткой шейкой, широким большим вертелом, мощными малым вертелом и межвертельным гребнем и массивным дистальным эпифизом – как у молодых (рис. 4а, 4б), так и у взрослых особей.
Размеры черепа приведены в табл. 1 , нижней челюсти и зубов – в табл. 2, бедренной кости – в табл. 3.
Таблица 3.
Размеры (в мм) бедренной кости Hystrix refossa Gervais, 1852 из нижнего плейстоцена пещеры Таврида, Крым
| Параметр | Экземпляр ПИН, № | ||
|---|---|---|---|
| 5644/508 | 5644/510 | 5644/511 | |
| Длина от головки до края дистального эпифиза | 135.6 | ~148.0 | 151.0 |
| Поперечный диаметр головки | 21.3 | 25.8 | 24.7 |
| Минимальный поперечный диаметр диафиза | 20.0 | 22.1 | 21.9 |
| Минимальный передне-задний диаметр диафиза | 13.5 | 15.0 | 14.8 |
| Максимальный передне-задний диаметр дистального эпифиза | 43.0 | 44.5 | 40.9 |
Экз. ПИН, № 5644/555, OL × OW: P4 – 9.8 × 7.8 (dex), 10.0 × 8.0 (sin); M1 – 9.9 × 9.5 (dex), 9.8 × 9.6 (sin); M2 – 10.1 × 9.6 (dex), 10.3 × 9.8 (sin); M3 – 8.7 × 7.0 (dex), 8.8 × 7.0 (sin). Экз. ПИН, № 5644/501: AL DP4–M3 – 37.0; AL × AW: DP4 – 8.5 × 11.5, M1 – 9.0 × 10.0, M2 – 9.5 × 9.8, M3 – 9.5 × 7.7.
I1 (экз. ПИН, № 5644/544): LD – 8.9, TD – 7.5. Стертый DP4 (экз. ПИН, № 5644/532): OL – 11.1, OW – 14.1. Нестертый P4 (экз. ПИН, № 5644/502): OL – 10.0; L – 11.8; OW – 7.8; W – 10.0; лингвальная эмалевая высота коронки – 23.2; высота гипострии – 4.0.
Нижнечелюстные кости: длина от переднего края резца до заднего края сочленовного отростка – 110.5 (экз. ПИН, № 5644/503, dex), 125.5 (экз. ПИН, № 5644/504, dex), 140.0 (экз. ПИН, № 5644/525); высота в сочленовном отростке – 41.2 (экз. ПИН, № 5644/534), 47.7 (экз. ПИН, № 5644/512), 50.8 (экз. ПИН, № 5644/525); высота с лингвальной стороны под DP4/P4 – 22.2–31.9, под M1 – 22.6–29.2 (n = 15); толщина спереди от нижнего массетерного гребня – 12.5–17.1 (n = 15).
Изменчивость. Отмеченные выше стадии прорезывания, смены и стирания зубов (рис. 3) позволяют разделить нижнечелюстной материал из пещеры Таврида на возрастные группы. Смена DP4 на P4 у современных дикобразов рода Hystrix происходит в возрасте около 1.5 лет, когда они достигают половой зрелости; с этой онтогенетической стадией соотносится стадия зубного стирания C [8]. По аналогии с современными H. cristata Linnaeus, 1758 и H. africaeaustralis Peters, 1852 стадии A, B1, B2, D1, D2, E сопоставляются соответственно с возрастными группами 0–5, 5–8, 8–18, 18–30, 30–48 мес и 4–12 лет [8]. С учетом данных по зубным возрастным классам H. africaeaustralis [10, табл. 1 ], описанные выше стадии H. refossa рассматриваются в пределах шести возрастных групп: 5–6 мес (A/B1, ювенильные особи), 8–11 мес (B1/B2, молодые неполовозрелые особи), 18–23 (C, C/D1), 24–30 (D1), 30–48 (D2) мес (C–D2, взрослые особи) и 4–12 лет (D2/E, E, стареющие взрослые и сенильные особи).
Нижнечелюстные кости более возрастных особей характеризуются большей общей длиной (стадия C – 110.5; стадия D2 – 125.5; стадия D2/E – 140.0), более длинной диастемой (табл. 2), большей высотой в сочленовном отростке (стадия B1/B2 – 41.2; стадия D1 – 47.7; стадия D2/E – 50.8) и на уровне передних щечных зубов (стадии A/B1 и B1/B2: под DP4/P4 – 22.2–25.6, под M1 – 22.6–24.1; стадии С–E: под DP4/P4 – 25.0–31.9, под M1 – 26.5–29.2), а также большей массивностью и толщиной горизонтальной ветви (стадии A/B1 и B1/B2 – 12.5–13.7; стадии С–E – 14.1–17.1).
Экз. ПИН, № 5644/531 (рис. 2и, 3р) демонстрирует аномально неглубокие альвеолы выпавших левых P4–M3, частично заполненные пористой костной тканью. Особенно сильно выположена лунка M1. Очевидно, данная особь при жизни утратила все щечные зубы в левой половине нижней челюсти, и зубные лунки начали залечиваться. Судя по размерам (высота горизонтальной ветви под P4 – 29.0, под M1 – 28.0, толщина – 16.2), это животное принадлежало к взрослой возрастной группе, однако причина выпадения у него щечных зубов неясна. Признаков периодонтита или травмы нет. С территории Сицилии (Италия) описан череп взрослого самца современного дикобраза H. cristata (Музей естественной истории Феррары, экз. № 21B) с зубными лунками левых P4–M2 аномально малой глубины (сами зубы утрачены при жизни) [11, рис. 2, 3]. В нижней челюсти отсутствуют правый P4 и его альвеола, а оба M1, их корни и лунки сильно редуцированы (причем левый M1 выпал при жизни). Можно предположить, что в обоих случаях (экз. № 21B; экз. ПИН, № 5644/531) из-за генетических отклонений сформировались аномально короткие зубные корни, что привело к прижизненной утрате щечных зубов с последующим частичным зарастанием (цикатризацией) зубных лунок [см. 11, с. 223].
Сравнение и замечания. Согласно Д. Ван Веерсу [3], синонимами H. refossa являются H. major Gervais, 1859 (плейстоцен, Европа), H. crassidens Lydekker, 1886 (плейстоцен, Индия), H. etrusca Bosco, 1898 (ранний плейстоцен, Европа), H. angressi Frenkel, 1970 (поздний плейстоцен, Израиль), H. gigantea Van Weers, 1985 (поздний плейстоцен, Ява, Индонезия) и H. magna Pei, 1987 (плейстоцен, Китай). H. makapanensis Greenwood, 1958 из плио-плейстоцена Южной и Восточной Африки, также иногда рассматривавшийся как синоним H. refossa [2], в настоящее время обычно считается самостоятельным видом [12]. H. refossa четко отличается от других плейстоценовых дикобразов сочетанием крупных размеров и гипсодонтных щечных зубов. По степени гипсодонтности крупный дикобраз из Крыма (отношение эмалевой высоты коронки к длине жевательной поверхности для нестертого P4 – 2.32) входит в интервал значений, свойственных H. refossa (1.5–2.6) [2].
По строению и размерам черепа крупный дикобраз из пещеры Таврида соответствует раннеплейстоценовым H. refossa из Европы [см. 13], но отличается от позднеплейстоценового H. refossa (= H. angressi) из Израиля [см. 6, 7]. В числе отмеченных отличий более крупные размеры, субпараллельное положение верхних зубных рядов и субвертикальное положение длинной оси затылочных мыщелков. В строении зубов крымский большой дикобраз отличается от израильской формы закрытой парафоссеттой на нестертых и слабо стертых P4, а также изолированным положением лабиального мезофоссеттида и отсутствием связи заднего мезофлексида с гипофлексидом на слабо стертых M1.
По длине зубных рядов (AL P4–M3 – 37.0–39.0; OL P4–M3 – 37.7–37.8; AL P4–M3 – 39.2–46.5; OL P4–M3 – 38.5–43.0) большой дикобраз из Тавриды соответствует H. refossa (AL P4–M3 – 37.0–40.3; OL P4–M3 – 39.7; AL P4–M3 – 37.5–42.3; OL P4–M3 – 39.8–42.0), но в случае нижних зубных рядов превосходит все известные выборки по максимальным и средним значениям [6, 7, 14, 15] (табл. 4). Нужно отметить, что некоторые сводные данные о размерах H. refossa [15, табл. 1 ] неточны (в частности, окклюзиальная длина P4–M3 – 43.5), так как, видимо, частично основаны на экземплярах других крупных вымерших видов дикобразов – H. primigenia (Wagner, 1848) [см. 2, табл. 2] и H. depereti Sen, 2001 [см. 16].
Таблица 4.
Размеры (в мм) зубов Hystrix refossa Gervais, 1852 из плейстоцена Европы и Передней Азии (AL – альвеолярная длина; OL – окклюзиальная длина; n – число экземпляров; μ – среднее значение)
| Параметр | Крым: пещера Таврида | Италия: Вальдарно [2], Пирро-Норд [18] | Израиль: пещера Геула [6, 7] |
|---|---|---|---|
| AL P4–M3 | 37.0–39.0 (n = 3, μ = 38.2) | 37.0–40.3 (n = 2, μ = 38.7) [2] | 37.7–39.5 (n = 4, μ = 38.7) |
| OL P4–M3 | 37.7–37.8 (n = 2, μ = 37.75) | 39.7 (n = 1) [2] | – |
| AL P4–M3 | 39.2–46.5 (n = 9, μ = 44.7) | 41.1 (n = 1) [2]; 39.8–42.3 (n = 2, μ = 41.0) [18] | 37.5–40.6 (n = 9, μ = 38.5) |
| OL P4–M3 | 38.5–43.0 (n = 7, μ = 41.8) | 39.8 (n = 1) [2]; 40.6 (n = 1) [18] | – |
| OL P4 | 9.8–11.1 (n = 4, μ = 10.2) | 10.6–11.95 (n = 2, μ = 11.3) [18] | 9.2–11.0 (n = 24, μ = 10.3) |
| OL M1 | 9.8–9.9 (n = 2, μ = 9.85) | M1–2: 6.7–10.85 (n = 13, μ = 9.2) [18] | 6.8–10.0 (n = 24, μ = 9.0) |
| OL M2 | 10.1–10.3 (n = 2, μ = 10.2) | 7.8–11.0 (n = 29, μ = 9.7) | |
| OL M3 | 8.7–8.8 (n = 2, μ = 8.75) | 6.7–10.05 (n = 7, μ = 8.9) [18] | 7.1–10.5 (n = 28, μ = 9.0) |
| OL P4 | 11.0–13.4 (n = 8, μ = 12.1) | 11.9 (n = 1) [18] | 9.2–13.0 (n = 42, μ = 10.9) |
| OL M1 | 7.9–11.8 (n = 15, μ = 10.3) | M1-2: 8.75–10.4 (n = 2, μ = 9.6) [18] | 7.1–10.7 (n = 28, μ = 9.1) |
| OL M2 | 9.9–11.7 (n = 14, μ = 10.9) | 8.4–11.0 (n = 26, μ = 9.7) | |
| OL M3 | 9.0–10.4 (n = 9, μ = 9.65) | 9.85–10.5 (n = 3, μ = 10.1) [18] | 8.3–11.0 (n = 42, μ = 9.9) |
Длина отдельных зубов (P4 – 9.8–11.1; M1–2 – 9.8–10.3; M3 – 8.7–8.8; P4 – 11.0–13.4; M1–2 – 7.9–11.8; M3 – 9.0–10.4) в целом отвечает крупным экземплярам H. refossa (длина P4 – 8.7–11.95; M1–2 – 6.7–11.0; M3 – 6.7–10.5; P4 – 9.2–13.0; M1–2 – 7.1–11.6; M3 – 8.3–11.0) [6, 7, 15, 17, 18], хотя превосходит их по максимальным и средним значениям P4 и M1–2 (см. табл. 4). Значение изменчивости в размерах следует оценить при получении дополнительных материалов по европейским представителям вида.
Hystrix refossa примерно на 20% крупнее современных H. indica (Kerr, 1792) [4] и H. cristata L., 1758. Кроме того, для H. refossa характерны очень массивные кости посткраниального скелета, даже по сравнению с крупными позднемиоценовыми формами H. primigenia и H. depereti. Вероятно, более тяжелое телосложение (как и гипсодонтные зубы) H. refossa связано с адаптацией к жизни в сухих и открытых ландшафтах [17, 18]. При этом по величине зубов H. refossa уступал H. depereti из терминального миоцена Франции (AL P4–M3 – 49.5–50.5; OL M1–2 – 10.9–14.0, n = 10, μ = 12.1), H. paukensis из мио-плиоцена Мьянмы (OL P4 – 15.06; OL M1 – 11.97; OL M2 – 11.56; OL M3 – 11.13) и Xenohystrix crassidens из плиоцена Южной и Восточной Африки (OL P4 – 14.0–17.0, n = 3, μ = = 15.7; OL M1 – 14.0; OL M2 – 14.5) [16, 19, 20].
Остатки нескольких разновозрастных особей H. refossa (экз. ПИН, №№ 5644/503, 504, 508) были найдены в пещере Таврида совместно, что позволяет предположить их первичную концентрацию в пределах логова или норы плейстоценовых дикобразов. Часть костей (например, экз. ПИН, №№ 5644/504, 509, 522, 524, 527, 531) несет следы погрызов, видимо, оставленные H. (A.) vinogradovi и мелкими грызунами (рис. 2в, 2г, 2з, 2и); очевидно, до захоронения в осадке кости некоторое время оставались на поверхности. Особенно примечательна находка бедренной кости (экз. ПИН, № 5644/511) с явственными отметинами (ямками и бороздками) от зубов хищного млекопитающего (рис. 4ж–4л).
Распространение. Поздний плиоцен (MN16a) – плейстоцен Евразии.
Материал. Из пещеры Таврида неполный череп с обоими рядами щечных зубов (экз. ПИН, № 5644/555), фрагменты черепов (экз. ПИН, №№ 5644/500, 550), фрагмент предчелюстной кости с I1 (экз. ПИН, № 5644/544), фрагменты верхнечелюстных костей без зубов (экз. ПИН, №№ 5644/501, 543), изолированный DP4 (экз. ПИН, № 5644/532), изолированный P4 (экз. ПИН, № 5644/502), парные нижнечелюстные кости (экз. ПИН, №№ 5644/503, 504), нижнечелюстные кости и их фрагменты (экз. ПИН, №№ 5644/509, 512, 521–527, 531, 534–536), атлант (экз. ПИН, № 5644/505), эпистрофей (экз. ПИН, № 5644/506), фрагмент плечевой кости (экз. ПИН, № 5644/515), локтевые кости и их фрагменты (экз. ПИН, №№ 5644/507, 513, 516, 519), бедренные кости и их фрагменты (экз. ПИН, №№ 5644/508, 510, 511, 518) и другие кости посткраниального скелета.
Список литературы
Лопатин А.В. Дикобраз Hystrix (Acanthion) vinogradovi (Rodentia, Hystricidae) из раннеплейстоценового местонахождения Таврида в Крыму // Доклады Академии наук. 2019. Т. 486. № 6. С. 756–762.
Van Weers D.J. The porcupine Hystrix refossa Gervais, 1852 from the Plio-Pleistocene of Europe, with notes on other fossil and extant species of the Hystrix // Scripta Geologica. 1994. V. 106. P. 35–52.
Van Weers D.J. A taxonomic revision of the Pleistocene Hystrix (Hystricidae, Rodentia) from Eurasia with notes on the evolution of the family // Contributions to Zoology. 2005. V. 74. № 3/4. P. 301–312.
Lazaridis G., Tsoukala E., Maul L.C. The earliest Hystrix refossa: a new Early Villafranchian record from Milia (Grevena, Macedonia, Greece) // Hystrix, the Italian Journal of Mammalogy. 2019. V. 30. № 1. P. 12–18.
Топачевский В.А. Грызуны таманского фаунистического комплекса Крыма. Київ: Наук. думка, 1973. 235 с.
Frenkel H. Hystrix angressi sp. nov. a large fossil porcupine from the Levalloiso-Mousterian of the Geula Cave // Israel Journal of Zoology. 1970. V. 19. № 1. P. 51–82.
Monchot H., Fernandez P., Gaillard J.-M. Paleodemographic analysis of a fossil porcupine (Hystrix refossa Gervais, 1852) population from the Upper Pleistocene site of Geula Cave (Mount Carmel, Israel) // Journal of Archaeological Science. 2012. V. 39. P. 3027–3038.
Gervais P. Fossiles dans les alluvions sous-volcanique de la montagne de Perrier, aux environs d’Issoire (Puy-de-Dôme) // Zoologie et paléontologie francaise. V. 1–3. Paris: A. Bertrand, 1848–1852. P. 1–271.
Wang B.Y., Qiu Z.D. New Hystrix (Hystricidae, Rodentia) from the Neogene of Linxia Basin, Gansu, China // Vertebrata PalAsiatica. 2020. V. 58. № 3. P. 204–220.
Van Aarde R.J. Age determination of Cape porcupines, Hystrix africaeaustralis // South African Journal of Zoology. 1985. V. 20. № 4. P. 232–236.
Angelici F.M., Luiselli L. Extra teeth and dental anomalies in the crested porcupine Hystrix cristata, from Sicily // Acta Theriologica. 1999. V. 44. № 2. P. 219–223.
Adams J.W. Craniodental and postcranial remains of the extinct porcupine Hystrix makapanensis Greenwood, 1958 (Rodentia: Hystricidae) from Gondolin, South Africa // Annals of the Ditsong National Museum of Natural History. 2012. V. 2. № 1. P. 7–17.
Bosco C. Hystrix etrusca n. sp. // Paleontografia Italica. 1889. V. 4. P. 141–153.
Vekua A., Bendukidze O., Bukhsianidze M., et al. Porcupine in the Late Neogene and Quaternary of Georgia // Bulletin of the Georgian National Academy of Sciences. 2010. V. 4. № 3. P. 140–149.
Salari L., Sardella R. Il genere Hystrix Linnaeus, 1758 in Italia // Atti della Società Toscana di Scienze Naturali, Memorie Ser. A. 2011. V. 116. P. 171–178.
Sen S. Early Pliocene porcupine (Mammalia, Rodentia) from Perpignan, France: a new systematic study // Geodiversitas. 2001. V. 23. № 2. P. 303–312.
Rook L., Sardella R. Hystrix refossa Gervais, 1852 from Pirro Nord (Early Pleistocene, Southern Italy) // Rivista Italiana di Paleontologia e Stratigrafia. 2005. V. 111. № 3. P. 489–496.
Rook L., Sardella R. New data on the Early Pleistocene large sized porcupine from Pirro Nord (Apricena, Apulia, Italy) // Palaeontographica. Abt. A. 2013. V. 298. № 1–6. P. 87–94.
Nishioka Y., Zin-Maung-Maung-Thein, Egi N., et al. New Hystrix (Mammalia, Rodentia) from the Late Miocene/Early Pliocene of Myanmar // Journal of Vertebrate Paleontology. 2011. V. 31. № 4. P. 919–924.
Greenwood M. Fossil Hystricoidea from the Makapan Valley, Transvaal // Palaeontologia Africana. 1955. V. 3. P. 77–85.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни