

Доклады Российской академии наук. Науки о жизни, 2023, T. 510, № 1, стр. 308-315
EPTESICUS NILSSONII VARANGUS SUBSP. NOV. (VESPERTILIONIDAE, CHIROPTERA) ИЗ НИЖНЕГО ПЛЕЙСТОЦЕНА ПЕЩЕРЫ ТАВРИДА В КРЫМУ
Академик РАН А. В. Лопатин 1, *
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 30.01.2023
После доработки 07.02.2023
Принята к публикации 10.02.2023
- EDN: QHRGYE
- DOI: 10.31857/S2686738923600073
Аннотация
По неполному черепу и фрагменту нижнечелюстной кости из нижнеплейстоценовых отложений пещеры Таврида в центральном Крыму описан новый подвид северных кожанков Eptesicus nilssonii varangus subsp. nov. Это древнейшая находка данного вида. Присутствие E. nilssonii (Keyserling et Blasius, 1839) в составе раннеплейстоценовой ассоциации рукокрылых пещеры Таврида указывает, что до вселения в Центральную и Юго-Восточную Европу этот вид обитал на юге Восточной Европы.
Среди раннеплейстоценовых рукокрылых из пещеры Таврида в центральном Крыму (Белогорский район, пос. Зуя) ранее были описаны Rhino-lophus macrorhinus cimmerius Lopatin, 2022, R. mehelyi scythotauricus Lopatin, 2023 (Rhinolophidae) и Eptesicus praeglacialis Kormos, 1930, а также отмечены представители родов Myotis и Plecotus (Vespertilionidae) [1–3]. Сборы 2022 г. позволили значительно увеличить коллекцию ископаемых летучих мышей из пещеры Таврида [3] и дополнить таксономический состав ассоциации несколькими формами (включая мелкие виды Myotis и Vespertilio). Одной из важных новых находок является северный кожанок Eptesicus nilssonii (Keyserling et Blasius, 1839). Этот бореальный вид в настоящее время обитает в Северной Евразии. В европейской части ареала он распространен на юге до стран Центральной Европы. В палеонтологической летописи E. nilssonii отмечен с конца раннего плейстоцена Европы [4–8].
Кожанок из раннего плейстоцена Крыма ниже описан как Eptesicus nilssonii varangus subsp. nov. Фаунистический комплекс пещеры Таврида датирован поздним виллафранком, около 1.8–1.5 млн л.н. [9, 10].
Изученный материал хранится в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН) в г. Москве. Для сравнения использованы черепа современных E. nilssonii из коллекции Зоологического музея Московского государственного университета имени М.В. Ломоносова (ЗМ МГУ). Измерения сделаны с помощью окуляр-микрометра стереомикроскопа Leica MZ6, приведены в мм. Иллюстрации подготовлены при помощи цифрового фотоаппарата Nikon D800 с объективом AF-S Micro NIKKOR 60mm f/2.8G ED, сканирующих электронных микроскопов Tescan Vega II XMU, Tescan Vega 3 XMU и рентгеновского микротомографа Neoscan N80 в ПИН.
Отряд Chiroptera Blumenbach, 1779
Семейство Vespertilionidae Gray, 1821
Подсемейство Vespertilioninae Gray, 1821
Триба Eptesicini Volleth et Heller, 1994
Род Eptesicus Rafinesque, 1820
Eptesicus nilssonii (Keyserling et Blasius, 1839)
Eptesicus nilssonii varangus Lopatin, subsp. nov.
Название подвида – лат. varangus, варяг, чужеземный наемник.
Голотип – ПИН, № 5644/680, неполный череп с правыми M1–M3 и фрагментом левого P4; Россия, Республика Крым, Белогорский район, местонахождение Таврида (точка “Ванин грот”); нижний плейстоцен.
Описание (рис. 1–4). Размеры черепа как у современных E. nilssonii nilssonii, но ростральный отдел шире, а зубы немного крупнее. Череп компактный, мозговая коробка округлая, выпуклая, несколько шире и намного выше лицевого отдела. Заглазничное сужение четко выражено.
Рис. 1.
Eptesicus nilssonii varangus Lopatin, subsp. nov.: а–д – голотип ПИН, № 5644/680, неполный череп с правыми M1–M3 и фрагментом левого P4: а – с дорсальной стороны, б – с вентральной стороны, в – с левой латеральной стороны, г – с правой латеральной стороны, д – с ростральной стороны; е–з – экз. ПИН, № 5644/681, фрагмент левой нижнечелюстной кости с P4 и M2: е – с лабиальной стороны, ж – с окклюзиальной стороны, з – с лингвальной стороны; Россия, Крым, пещера Таврида; нижний плейстоцен.
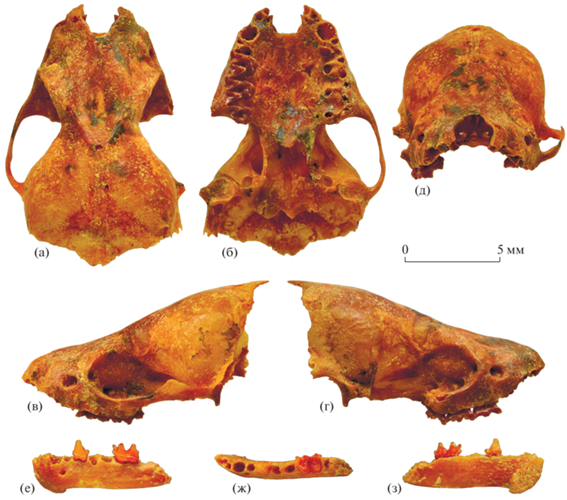
Рис. 2.
Eptesicus nilssonii varangus Lopatin, subsp. nov., голотип ПИН, № 5644/680, неполный череп с правыми M1–M3 и фрагментом левого P4 с вентральной стороны: а – ростральная часть черепа; б – то же с прорисовкой следов небных валиков; в, г – костное небо (СЭМ Tescan Vega II XMU); Россия, Крым, пещера Таврида; нижний плейстоцен.
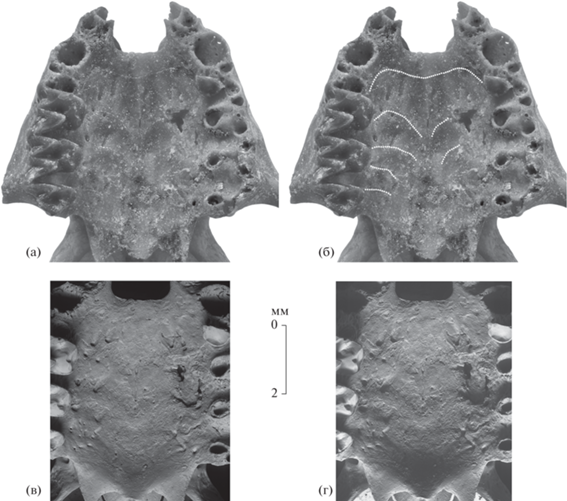
Рис. 3.
Eptesicus nilssonii varangus Lopatin, subsp. nov., голотип ПИН, № 5644/680, неполный череп с правыми M1–M3 и фрагментом левого P4, компьютерная модель: а – с вентральной стороны; б – левая половина носовой полости, сагиттальный срез вблизи носовой перегородки (видны этмотурбиналии I, II, III и максиллотурбиналия); в – носовая полость, поперечный срез на уровне передней части M1; Россия, Крым, пещера Таврида; нижний плейстоцен.
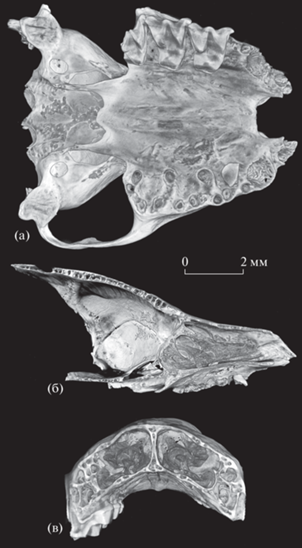
Рис. 4.
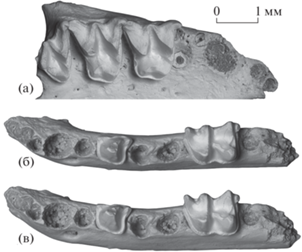
Eptesicus nilssonii varangus Lopatin, subsp. nov., зубные ряды (СЭМ Tescan Vega 3 XMU): а – голотип ПИН, № 5644/680, правые M1–M3 и альвеолы I1–P4 с окклюзиальной стороны; б, в – экз. ПИН, № 5644/681, левые P4 и M2 и альвеолы I1–P2, M1 и M3: б – с окклюзиальной стороны, в – с окклюзиальной стороны с лингвальным наклоном; Россия, Крым, пещера Таврида; нижний плейстоцен.
Лицевой отдел черепа массивный, ростральная часть короткая, широкая, с дорсальной стороны отчетливо вогнутая, с узкой углубленной продольной срединной впадиной между глазницами (рис. 1). Конец рострума плавно изогнут, перед клыками имеются отчетливые антеролатеральные впадины. Вырезка носового отверстия глубокая, V-образная. Лобные кости расширены над глазницами, предглазничные отростки отсутствуют, слезные вздутия хорошо выражены. Надглазничные гребни отчетливые, сагиттальный гребень не развит. Теменные отверстия крупные.
Подглазничное отверстие крупное, округлое, расположено на уровне задней части P4 и передней части M1. Отделяющий подглазничное отверстие от глазницы костный мостик довольно широкий. Расположенное в передней стенке глазницы мелкое округлое слезное отверстие находится на уровне задней части M1. Скуловая дуга плавно изогнутая, относительно высокая, с крупным треугольным заглазничным отростком.
Костное небо широкое. Задний край широкой U-образной передней небной вырезки находится на уровне передней части альвеол верхних клыков C1. Альвеолы C1 широко разнесены. Срединный шип костного неба короткий и широкий. Мезоптеригоидная ямка широкая. Суставные впадины поперечно короткие, округлые.
На костном небе сохранились следы пяти из семи характерных для вида (см. [11–13]) небных валиков (поперечных небных складок, rugae palatinae transversae vel plicae palatinae), включая небные валики II–VI. Эти следы имеют вид субпараллельных изогнутых тонких линий (поверхностных бороздок) белесого цвета на месте оснований небных валиков (рис. 1б, 2а, 2б) (см. [1]). Непрерывная линия небного валика II дважды волнообразно изогнута, ее плавно прогнутая назад срединная часть протягивается до уровня супротивно расположенных промежутков C1/P4, выступающие вперед парные лопасти достигают уровня заднего края альвеол правого и левого C1, а латеральные окончания находятся у средней части лингвальных альвеол противолежащих P4. Линии небных валиков III–VI сильно прогнуты дистально и прерываются в осевой части костного неба; таким образом, каждая из них имеет вид разомкнутой в центре двойной дуги. Парные дуги сильно изгибаются вперед у линии небного валика III и становятся все более пологими в последовательности небных валиков IV–VI. Мезиальные выступы парных дуг линии небного валика III находятся на уровне протокона противолежащих M1, а их латеральные окончания достигают уровня выступа гипокона M1. Латеральные концы дуг линии небного валика IV располагаются на уровне переднего края противолежащих M2, небного валика V – на уровне выступа гипокона M2, небного валика VI – на уровне переднего края противолежащих M3. На месте небных валиков III–V имеются трудно различимые шевронообразные утолщения костного неба (видны на СЭМ, см. рис. 2в, 2г). Следы терминальных небных валиков I и VII не сохранились.
Носовые раковины (турбиналии), изученные с помощью компьютерной томографии (рис. 3), демонстрируют характерное для Vespertilionidae строение [14]. Четко идентифицируются крупные этмотурбиналии I, II и III, занимающие значительную часть объема носовой полости, и продольно вытянутая максиллотурбиналия, расположенная вблизи дна носовой полости (см. [2]).
Верхний зубной ряд включает два резца, клык, один премоляр и три моляра (рис. 4а). Судя по альвеолам I1 и I2, верхние резцы тесно сближены и I1 несколько превосходит по величине I2. Альвеола C1 округлой формы; она отделена от альвеолы I2 небольшим промежутком, но тесно сближена с альвеолой антеролабиального корня P4.
Верхние моляры M1 и M2 массивные, с окклюзиальной поверхностью трапециевидных очертаний и крупными выступающими стилями. M2 расширен поперечно сильнее, чем M1, парастиль и метастиль на M2 немного крупнее. Лингвальная доля M1 и M2 заметно сжата в продольном направлении, особенно на M2. Протокон на каждом из этих зубов выдается лингвально сильнее, чем выступ гипокона. Выступ гипокона хорошо развитый, с бугорковидной вершиной. Цингулюм тонкий. Задняя вырезка очень слабая. M3 слабо редуцированный, широко-треугольный, с крупным парастилярным выступом. Метакон хорошо развит, метастиль отсутствует. Цингулюм тонкий, но четкий, непрерывный.
Горизонтальная ветвь нижнечелюстной кости относительно высокая, с хорошо развитым подбородочным выступом. Подбородочное отверстие мелкое, округлое, расположено под промежутком C1/P2. Симфиз крупный, достигает уровня передней части альвеолы P2.
Нижний зубной ряд состоит из трех резцов, клыка, двух премоляров и трех моляров (рис. 4б, 4в). Судя по альвеолам, нижние резцы мелкие, наклонены вперед и скошены лабиально. Альвеола I2 расположена строго позади альвеолы I1 и непосредственно лингвальнее передней части альвеолы I3 и, таким образом, смещена лингвально относительно осевой линии ряда резцов. Альвеола C1 крупная, овальная, альвеола P2 в 1.5 раза меньше ее.
P4 массивный, основание коронки трапециевидной формы, с непрерывным мощным цингулидом. Антеролингвальный и постеролингвальный дополнительные бугорки хорошо развитые, сильно выступающие. Лингвальная сторона основания коронки между этими дополнительными бугорками почти прямая, с небольшой впадиной на уровне вершины главного бугорка. Вершина главного бугорка трехгранная, с лингвальной фасеткой стирания, лабиальная сторона главного бугорка выпуклая, задняя сторона немного вогнутая, основание постеролингвального ребра с заметным утолщением.
M2 массивный, компактный, с лабиальными бугорками (протоконидом и гипоконидом) приблизительно равной высоты (при умеренном стирании). Талонид немного шире тригонида. Энтокристид неполный, в передней части этот мощный гребень резко понижается и соединяется с основанием тонкого дистального метакристида, восходящего на заднюю стенку метаконида. При этом глубокая V-образная вырезка между дистальным метакристидом и энтокристидом немного не достигает дна талонидного бассейна, и он остается закрытым с лингвальной стороны. Гипоконулид умеренно развит, заметно выступает постеролингвально. Эктоцингулид мощный, энтоцингулид выражен в виде короткого тонкого гребешка, расположенного базальнее выхода из тригонидного бассейна.
Размеры в мм. Голотип ПИН, № 5644/680: длина рострума (до заднего края небного отростка верхнечелюстной кости) – 6.2; длина костного неба (от передней небной вырезки до основания срединного шипа) – 5.8; длина суставной впадины (dex – правая, sin – левая) – 1.4 (dex, sin); ширина суставной впадины – 1.6 (dex, sin); межглазничная ширина (в заглазничном сужении) – 4.15; наименьшее расстояние от сагиттального шва до латерального выступа левой скуловой дуги – 5.1 (реконструируемая ширина черепа в скуловых дугах – около 10.2); ширина мозговой коробки – 8.2; ширина на уровне альвеол клыков – 5.35; наименьшее расстояние от лабиального края выступа метастиля M2 до уровня срединного небного шва – 3.5 (реконструируемая ширина на уровне M2–M2 – около 7.0); наименьшее расстояние от лабиального края выступа парастиля M3 до уровня срединного небного шва – 3.5 (реконструируемая ширина на уровне M3–M3 – около 7.0); ширина мезоптеригоидной ямки (между основаниями крыловидных отростков) – 2.55; альвеолярная длина C1–M3 – 5.7 (sin); длина M1–M3 – 3.85 (dex); размеры зубов (L – наибольшая длина, LabL – лабиальная длина, LinL – лингвальная длина, W – наибольшая ширина): M1: LabL – 1.6; LinL – 1.05; W – 1.7; M2: LabL – 1.55; LinL – 0.95; W – 1.8; M3: L – 0.95; W – 1.75.
Экз. ПИН, № 5644/681: альвеолярная длина I1–M3 – около 6.9; высота горизонтальной ветви в области симфиза – 2.17; под P4 – 1.5; под M2 – 1.5; размеры зубов: P4: L – 0.88; W – 0.73; M2: L – 1.45; ширина тригонида – 0.9; ширина талонида – 0.92.
Сравнение и замечания. Отнесение раннеплейстоценового мелкого кожана из пещеры Таврида к виду E. nilssonii основывается на строении и размерах черепа и зубов. Вместе с тем от современных представителей E. nilssonii (в составе подвидов E. nilssonii nilssonii и E. nilssonii parvus Kishida, 1932) крымская ископаемая форма отличается более широкой ростральной частью черепа (особенно хорошо это заметно по ширине черепа на уровне клыков и моляров, см. [15, 17] и табл. 1). В строении зубов E. nilssonii varangus subsp. nov. отличается от современного E. nilssonii nilssonii продольно сжатыми лингвальными долями M1–M2 (различия по параметрам LinL/LabL см. в табл. 1) и более тонким цингулюмом верхних моляров, а также неполным энтокристидом M2 (у E. nilssonii nilssonii энтокристид полностью слит с дистальным метакристидом и образует вместе с ним высокий продольный гребень с умеренно развитой вырезкой).
Таблица 1.
Размеры (в мм) Eptesicus nilssonii (Keyserling et Blasius, 1839) (n – число измеренных экземпляров; * – образцы из разных регионов России (Калининградская, Ленинградская, Новгородская, Тверская, Самарская, Нижегородская области, Ханты-Мансийский автономный округ – Югра, Республика Саха (Якутия), Забайкальский край), коллекция ЗМ МГУ, измерения автора)
| Параметры | E. nilssonii varangus subsp. nov. | E. nilssonii nilssonii, современные [5, 15, 17] |
||
|---|---|---|---|---|
| n | пределы | среднее | ||
| Длина рострума | 6.2 | 12* | 6.0–6.8 | 6.3 |
| Длина костного неба | 5.8 | 7* | 5.5–6.1 | 5.77 |
| Длина суставной впадины | 1.4 | 25* | 1.3–1.6 | 1.45 |
| Ширина суставной впадины | 1.6 | 18* | 1.5–1.7 | 1.64 |
| Межглазничная ширина | 4.15 | 42 [17] 24 [15] 14* |
3.8–4.5 3.71–4.28 3.9–4.55 |
4.18 4.04 4.2 |
| Ширина черепа в скуловых дугах | ~10.2 | 26 [17] 8* |
9.5–10.5 9.9–10.7 |
9.95 10.27 |
| Ширина мозговой коробки | 8.2 | 24 [15] 14* |
7.57–8.38 7.75–8.45 |
7.89 8.07 |
| Ширина на уровне C1–C1 | 5.35 (по альвеолам) | 40 [17] 24 [15] 13* |
4.6–5.2 4.51–5.11 4.55–5.2 |
4.88 4.84 4.88 |
| Ширина на уровне M2–M2 | ~7.0 | 14* | 6.05–6.65 | 6.36 |
| Ширина на уровне M3–M3 | ~7.0 | 41 [17] 24 [15] 13* |
5.9–6.9 6.02–6.63 6.2–6.7 |
6.26 6.38 6.47 |
| Ширина мезоптеригоидной ямки | 2.55 | 12* | 2.4–2.6 | 2.49 |
| Длина C1–M3 | 5.7 (по альвеолам) | 40 [17] 24 [15] 27* |
5.0–5.8 5.21–5.74 5.3–5.95 |
5.42 5.53 5.6 |
| Длина M1–M3 | 3.85 | 28* | 3.35–3.8 | 3.59 |
| Лабиальная длина (LabL) M1 | 1.6 | 28* | 1.3–1.55 | 1.41 |
| Лингвальная длина (LinL) M1 | 1.05 | 28* | 0.97–1.1 | 1.05 |
| LinL/LabL M1, % | 65.63 | 28* | 70.0–81.48 | 74.93 |
| Ширина M1 | 1.7 | 28* | 1.45–1.7 | 1.59 |
| Лабиальная длина M2 | 1.55 | 28* | 1.3–1.5 | 1.41 |
| Лингвальная длина M2 | 0.95 | 28* | 0.9–1.1 | 0.98 |
| LinL/LabL M2, % | 61.29 | 28* | 64.29–78.57 | 69.46 |
| Ширина M2 | 1.8 | 28* | 1.65–1.85 | 1.77 |
| Длина M3 | 0.95 | 27* | 0.85–1.0 | 0.95 |
| Ширина M3 | 1.75 | 27* | 1.6–1.75 | 1.7 |
| Длина I1–M3 | ~6.9 (по альвеолам) | 28* | 6.7–7.3 | 6.98 |
| Длина P4 | 0.88 | 6 [5] 28* |
0.78–0.9 0.7–0.85 |
0.85 0.79 |
| Ширина P4 | 0.73 | 6 [5] 28* |
0.74–0.78 0.65–0.75 |
0.76 0.71 |
| Длина M2 | 1.45 | 28* | 1.3–1.45 | 1.39 |
| Ширина талонида M2 | 0.92 | 28* | 0.9–1.05 | 0.97 |
Показатели ширины ростральной части черепа на уровне клыков (WC1–C1) и на уровне последних моляров (WM3–M3) у новой формы значительно превосходят средние значения этих параметров у современных мелких кожанов Палеарктики – E. nilssonii (табл. 1) и E. gobiensis Bobrinskoy, 1926 ([15], табл. 1). При этом по WC1–C1 различия крымской ископаемой формы с E. gobiensis существенно больше, чем с E. nilssonii nilssonii. У E. nilssonii varangus subsp. nov. этот показатель (в мм, n – число экземпляров) составляет 5.35 (по альвеолам), у E. nilssonii nilssonii – 4.51–5.11 (среднее – 4.84, n = 24; см. также табл. 1), у E. gobiensis gobiensis – 4.12–4.83 (среднее – 4.59, n = 16), у E. gobiensis bobrinskoi Kuzyakin, 1935 –4.07–4.39 (среднее – 4.21, n = 14). Наглядны также различия в отношении WC1–C1 к WM3–M3 (по средним значениям), отражающем степень сужения рострума: у E. nilssonii varangus subsp. nov. данный показатель (WC1–C1/WM3–M3, %) составляет около 76.4, у E. nilssonii nilssonii – 75.9, тогда как у E. gobiensis gobiensis – 69.8, а у E. gobiensis bobrinskoi – 68.8. Отношение WC1–C1 к длине C1–M3 (%): E. nilssonii varangus subsp. nov. (по альвеолам) – 93.9, E. nilssonii nilssonii – 82.6–91.7 (среднее – 87.5, n = 24), E. gobiensis gobiensis – 78.7–86.9 (среднее – 82.5, n = 16), E. gobiensis bobrinskoi – 76.3–84.6 (среднее – 80.8, n = 14). Вместе с тем недостаток материалов в настоящее время не позволяет охарактеризовать изменчивость размеров E. nilssonii varangus subsp. nov.
Eptesicus nilssonii varangus subsp. nov. намного мельче E. praeglacialis, также обнаруженного в местонахождении Таврида (у последнего ширина черепа в скуловых дугах – 14.2–15.6 мм, ширина на уровне C1–C1 – 7.0–7.65 мм, ширина на уровне M2–M2 – 8.6–9.6 мм, длина C1–M3 – 8.2–8.75 мм, длина I1–M3 – 10.4 мм, см. [2]).
Древнейшие известные ранее ископаемые остатки E. nilssonii датированы концом раннего плейстоцена [4–8]. В Центральной Европе находки этого вида редки в нижнем и среднем плейстоцене, но более обычны в верхнем плейстоцене и голоцене, что отражает увеличение числа бореальных видов летучих мышей в Европе на фоне общего похолодания конца плейстоцена [16]. По размерам E. nilssonii из плейстоцена Европы значимо не отличается от современных популяций этого вида [5, 17].
Ранее было установлено [18], что у современных и субфоссильных E. nilssonii иногда бывает развит рудиментарный верхний малый премоляр P2 (с одной или обеих сторон). Этот факт был интерпретирован как примитивная черта, противопоставляющая E. nilssonii прочим видам рода [18]. Отсутствие P2 у раннеплейстоценового E. nilssonii varangus subsp. nov., самого древнего представителя вида, показывает, что в линии Eptesicus утрата данного зуба произошла на более ранней эволюционной стадии; таким образом, появление P2 у некоторых современных особей E. nilssonii имеет атавистический характер.
Современный северный кожанок Eptesicus nilssonii – самый устойчивый к холоду вид летучих мышей, широко распространенный в северных районах Евразии (до 70°25′ с.ш. в Норвегии) [16, 19]. Он обитает в лесах, горных местностях, встречается в различных открытых биотопах; зимует в постройках, иногда в пещерах и шахтах [16], но не относится к числу троглофильных форм. Присутствие E. nilssonii в составе раннеплейстоценовой ассоциации рукокрылых пещеры Таврида указывает, что до вселения в Центральную и Юго-Восточную Европу этот вид обитал на юге Восточной Европы. Находка остатков E. nilssonii varangus subsp. nov. и E. praeglacialis [2] в данном местонахождении согласуется с представлениями о возникновении на рубеже плиоцена и плейстоцена у некоторых лесных форм европейских летучих мышей (включая Eptesicus) климатически обусловленной экологической адаптации – гибернации в пещерах (см. [20]).
Материал. Помимо голотипа (сборы 2022 г.), из пещеры Таврида фрагмент левой нижнечелюстной кости с P4 и M2, альвеолами I1–P2, M1 и M3 (экз. ПИН, № 5644/681; сборы 2020 г.).
Список литературы
Лопатин А.В. Раннеплейстоценовый подковонос Rhinolophus macrorhinus cimmerius subsp. nov. (Rhinolophidae, Chiroptera) из пещеры Таврида в Крыму // Доклады Российской академии наук. Науки о жизни. 2022. Т. 506. С. 396–405.
Лопатин А.В. Раннеплейстоценовый кожан Eptesicus praeglacialis (Vespertilionidae, Chiroptera) из пещеры Таврида в Крыму // Доклады Российской академии наук. Науки о жизни. 2023. Т. 508. С. 95–104.
Лопатин А.В. Rhinolophus mehelyi scythotauricus subsp. nov. (Rhinolophidae, Chiroptera) из нижнего плейстоцена пещеры Таврида в Крыму // Доклады Российской академии наук. Науки о жизни. 2023. Т. 509. С. 109–114.
Jánossy D. Pleistocene vertebrate faunas of Hungary. Budapest: Akadémiai Kiadó, 1986.
Wołoszyn B.W. Pliocene and Pleistocene bats of Poland // Acta Palaeontologica Polonica. 1987. V. 32. № 3–4. P. 207–325.
Salari L., Kotsakis T. Late Pleistocene bat assemblage with Eptesicus nilssonii from Grotta del Fossellone (Southern Latium, Central Italy) // Alpine and Mediterranean Quaternary. 2017. V. 30. № 2. P. 125–136.
Pazonyi P., Virág A., Gere K. et al. Sedimentological, taphonomical and palaeoecological aspects of the late early Pleistocene vertebrate fauna from the Somssich Hill 2 site (South Hungary) // Comptes Rendus Palevol. 2018. V. 17. P. 296–309.
Pazonyi P., Trembeczki M., Mészáros L., Szentesi Z. Preliminary report on the Early Pleistocene vertebrate sites of Beremend Crystal Cave (Beremend 16, South Hungary) and on their palaeoecological importance // Fragmenta Palaeontologica Hungarica. 2019. V. 36. P. 115–140.
Лопатин А.В., Вислобокова И.А., Лавров А.В. и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Доклады Академии наук. 2019. Т. 485. № 3. С. 381–385.
Лопатин А.В., Тесаков А.С. Раннеплейстоценовая белозубка Crocidura kornfeldi (Lipotyphla, Soricidae) из Крыма // Доклады Российской академии наук. Науки о жизни. 2021. Т. 501. № 1. С. 499–504.
Огнев С.И. Звери Восточной Европы и Северной Азии. Т. 1. Насекомоядные и летучие мыши. Москва– Ленинград: Главнаука, 1928.
Shimizu M., Miyao T. A study of bats. II. The palatal ridges of the bat // Journal of the Faculty of Liberal Arts, Shinshu University. 1967. Pt 2. № 1. P. 1–25.
Eisentraut M. Das Gaumenfaltenmuster der Säugetiere und seine Bedeutung für stammesgeschichtliche und taxonomische Untersuchungen // Bonner Zoologische Monographien. 1976. № 8. P. 1–214.
Ito K., Tu V.T., Eiting T.P. et al. On the embryonic development of the nasal turbinals and their homology in bats // Frontiers in Cell and Developmental Biology. 2021. V. 9. Art. 613545. P. 1–19.
Benda P., Mashkour M. A finding of Eptesicus gobiensis in an ancient salt mine in Iran and notes on the status of this bat in the Middle East (Mammalia: Chiroptera) // Journal of the National Museum (Prague). Natural History Ser. 2021. V. 190. № 1. P. 61–72.
Rydell J. Eptesicus nilssonii // Mammalian Species. 1993. № 430. P. 1–7.
Spitzenberger F. Die Nordfledermaus (Eptesicus nilssoni Keyserling & Blasius, 1839) in Österreich. Mammalia austriaca 10 (Mammalia, Chiroptera) // Annalen des Naturhistorischen Museums in Wien. 1986. V. 87B. P. 117–130.
Fadeeva T.V., Kruskop S.V. Rudimentary small upper premolar in the Northern bat, Eptesicus nilssoni (Vespertilionidae, Chiroptera) // Russian Journal of Theriology. 2008. V. 7. № 2. P. 77–80.
Rydell J., Strann K.B., Speakman J.R. First record of breeding bats above the Arctic Circle: northern bats at 68°–70° N in Norway // Journal of Zoology. 1994. V. 233. P. 335–339.
Rosina V.V., Sinitsa M.V. Bats (Chiroptera, Mammalia) from the Turolian of the Ukraine: phylogenetic and biostratigraphic considerations // Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen. 2014. V. 272. № 2. P. 147–166.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни