

Экология, 2019, № 2, стр. 88-100
Эволюционная экология в XXI веке: новые концепции и перспективы развития
А. Г. Васильев *
Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: vag@ipae.uran.ru
Поступила в редакцию 14.07.2018
После доработки 07.08.2018
Принята к публикации 03.08.2018
Аннотация
Рассмотрены представления об эволюционной экологии XX в. и ее современном концептуальном пространстве, дополненные новыми представлениями о содержании популяционной и эволюционной синэкологии. Предложены: 1) популяционно-ценотический подход к анализу сопряженной изменчивости ценопопуляций симпатрических видов и таксоценов в пространстве и историческом времени; 2) подход к оценке внутригруппового морфоразнообразия ценопопуляций симпатрических видов и таксоценов как меры стабильности морфогенеза. Рассмотрено особое методологическое направление – экспериментальная эволюционная экология. Обсуждаются перспективы становления эволюционной экологии как междисциплинарной прикладной науки в связи с необходимостью прогнозирования региональных биоценотических кризисов и быстрых коэволюционных перестроек биотических сообществ в техногенной среде на фоне прогнозируемых неблагоприятных трендов климатических изменений.
В 2019 г. 1 апреля исполняется 100 лет со дня рождения академика Станислава Семеновича Шварца, а с момента выхода написанной им ключевой монографии “Эволюционная экология животных” [1] прошло ровно 50 лет. За это время в науке произошли существенные изменения в представлениях об эволюционной экологии (ЭЭ) как особом направлении экологических и эволюционно-биологических исследований. В конце XX в. и начале XXI в. происходило быстрое изменение состава биоты во всех регионах Земли, вызванное нарастающим техногенным загрязнением и урбанизацией территорий, истреблением массовых ресурсных видов животных и растений, уничтожением возобновляемых природных ресурсов и вымиранием уязвимых видов [2]. Благодаря сочетанному влиянию антропогенных и климатических факторов возросли число биологических инвазий чужеродных видов и формирование “гибридных” рекомбинантных биотических сообществ с новыми биологическими свойствами [3]. Выделена особая исторически недавно возникшая эпоха Земли–Антропоцен [4–6], когда воздействие технически и технологически вооруженного и численно растущего человечества на биосферу стало существенным фактором, значимо влияющим на ее функционирование. В результате, согласно существующим прогнозам, на Земле ожидается проявление региональных и глобального экологических кризисов [7–9]. Считается, что необходимость прогнозирования экологических кризисных изменений приведет к тому, что скоро эволюционная экология станет прикладной наукой и займет в биологии такое же лидирующее положение, какое в настоящее время занимает молекулярная биология [10–13].
В последние десятилетия в биологии произошел пересмотр многих теоретических представлений, доминировавших на протяжении XX в. Изменились содержание и направления исследований в генетике, биологии развития, экологии и эволюционной биологии, что в первую очередь связано с появлением новых технологических возможностей молекулярной генетики. Особое значение также имеют многочисленные прямые доказательства роли эпигеномных перестроек (метилирование ДНК, изменение локализации мобильных элементов генома и др.) в проявлении длящихся модификаций фенотипа, а также реальности трансгенерационного наследования эпигенетических изменений структуры и функционирования генома, вызванных стрессовыми средовыми эффектами [14–16]. Благодаря этим открытиям представления, ранее в XX в. казавшиеся “маловероятными” или “полностью ошибочными”, в частности проблема “наследования приобретенных признаков” [17], перешли в разряд “полностью доказанных” и широко обсуждаемых [14–16, 18–20]. По этой причине возникла необходимость ревизии эволюционных представлений, включая пересмотр геноцентрической синтетической теории эволюции – СТЭ (в англоязычной версии Modern Synthesis – MS).
Начиная с конца XX в. в качестве альтернативы СТЭ рассматривается эпигенетическая теория эволюции (ЭТЭ), предложенная М.А. Шишкиным [21, 22] и опирающаяся на представления И.И. Шмальгаузена [23, 24] и К. Х. Уоддингтона [17, 25]. Быстрое развитие эпигенетики в XXI в. и растущее осознание ее ведущей роли во многих областях биологии [16, 26], медицины [27] и сельского хозяйства [28] в последнее время создало для ЭТЭ прочный базис, опирающийся на фундаментальные представления и технологии в области эпигенетики и эпигеномики.
Стремление, с одной стороны, сохранить прежние лидирующие позиции СТЭ, а с другой, снять противоречивость геноцентрических представлений о механизмах эволюционного процесса привело к появлению концепции расширенного эволюционного синтеза – РЭС (Extended Evolutionary Synthesis – EES) [27, 29, 30]. Согласно РЭС, необходимо дополнить СТЭ эпигенетическими представлениями о феномене мягкой наследственности (soft heredity), основанной на эпигенетической наследственности и трансгенерационном наследовании измененных эпигенетических профилей ДНК. Предложено сочетать эти представления с теорией конструирования ниши – ТКН (Niche Construction Theory – NCT) [30, 31], т.е. связать экологические и эволюционные представления об активно формируемой нише с генетикой и эпигенетикой. Поскольку новая эпигенетическая трактовка механизмов эволюции в русле РЭС и ЭТЭ допускает быстрые эволюционно-экологические перестройки за относительно короткие исторические времена, появляется реальная возможность если не управлять микроэволюционным процессом, то попытаться его обнаруживать и прогнозировать.
Пересмотр теоретических представлений на основе РЭС и/или ЭТЭ должен касаться и эволюционной экологии (ЭЭ), значительный вклад в появление и развитие которой внес С.С. Шварц [1, 32]. Поскольку ЭЭ по его представлениям во многом опиралась именно на концепции СТЭ, необходимо оценить современное содержание ЭЭ в сравнении с исходными представлениями. Поэтому цель настоящей статьи заключается в сравнении исходных и современных концепций эволюционной экологии и оценке дальнейших направлений ее развития.
СТАНОВЛЕНИЕ ЭВОЛЮЦИОННОЙ ЭКОЛОГИИ
Эволюционная экология междисциплинарна: она частично объединяет эволюционистику (эволюционную теорию), экологию популяций и сообществ, биогеоценологию, историческую экологию, биогеографию, филогеографию и филоценогенетику. Вероятно, ЭЭ возникла несколько раньше, чем этому направлению было дано название. Признаки ЭЭ проявились уже в эволюционной теории Ж.-Б. Ламарка. Экологические и эволюционные представления, развивавшиеся в России К.Ф. Рулье, также можно причислить к ЭЭ. Первым “эволюционным экологом” следует считать Ч. Дарвина, предложившего идею борьбы за существование и вытекающий из нее эволюционно-экологический механизм естественного отбора. К. Петрусевич озаглавил одну из своих статей “Теория эволюции Дарвина есть теория экологическая” [33]. Д.Н. Кашкаров подчеркнул, что “…эколог может и должен интересоваться вопросами эволюции” [34, с. 196]. Значительный вклад в представления, ведущие к современной ЭЭ, внесли Ч. Элтон [35], Д.Н. Кашкаров [34], С.А. Северцов [36, 37], а позднее – Дж. Хатчинсон [38, 39] и Р. Макартур [40].
Работы С.А. Северцова 30–40-х годов XX в. могут рассматриваться в качестве пионерной версии эволюционной экологии [36, 41]. С.А. Северцов, вероятно, первым применил в своих работах словосочетание “эволюционная экология”. Изначально он определял цикл своих исследований как изучение эволюции “отношений со средой в связи с морфогенезом”, поскольку стремился обосновать взгляды А.Н. Северцова о биологическом прогрессе и морфологических закономерностях эволюции, включая экологическую трактовку явлений ароморфоза и идиоадаптации. Позднее он пришел к более широкому пониманию проблемы, определяя ее как исследование “эволюции экологических отношений и … изменений форм и интенсивности борьбы за существование в связи с адаптиогенезом” [37].
Интересно относительно раннее упоминание термина “эволюционная экология” В.Н. Беклемишевым [42]. Рассматривая проблемы экологической паразитологии, он развил представления о необходимости создания сравнительной экологии, использующей в качестве единиц сравнения жизненные схемы вида – “совокупность всех взаимоотношений вида со всеми элементами его среды обитания и в первую очередь – совокупность приспособлений вида к совокупности условий его существования” [42, с. 8]. В англоязычной литературе есть эквивалент “жизненной схемы” – “life history” (жизненный цикл). В.Н. Беклемишев полагал, что сравнительная экология жизненных схем видов может стать основой для создания эволюционной экологии. Однако он считал, что появление эволюционной экологии пока отдаленная задача, так как еще не создана ее основа – система сравнительной экологии. Эта версия “эволюционной экологии” контекстно была ориентирована на развитие эволюционной паразитологии и сравнительной экологии паразитов и в последующем автором больше не развивалась [43].
По мнению С.С. Шварца, заложившего основы наиболее известной в мире российской версии эволюционной экологии [1, 32, 44], одним из ее предтеч был Гордон Ориэнс [45], который полагал, что генеральной теорией экологии является теория естественного отбора. Он, вероятно, первым в англоязычной литературе употребил термин “эволюционная экология”, но, скорее, в метафорическом смысле. Г. Ориэнс взял за основу точку зрения Эрнста Майра о делении всей биологии на функциональную и эволюционную и разделил экологию на функциональную и эволюционную. Г. Ориэнс в качестве примера типичного носителя эволюционно-экологических взглядов рассматривал выдающегося эколога и эволюциониста – Дэвида Лэка. Сам Д. Лэк [46], которого С.С. Шварц считал одним из провозвестников ЭЭ, согласился с Г. Ориэнсом, в свою очередь сославшись на его статью, и в названии своей работы обозначил данное направление как “Evolutionary Ecology”. Он видел основную ее задачу в изучении эволюционной обусловленности экологической специфики видов и внутривидовых форм, а также становления и развития экологических адаптаций. По Д. Лэку следует различать непосредственные экологические особенности организмов в конкретных условиях и те, которые закреплены в процессе эволюции как собственно экологические адаптации.
С.С. Шварц стремился показать, что эволюционная экология нацелена на “изучение изменений взаимоотношений организмов в процессе филогенетического развития отдельных групп” [1, с. 9] и экологических механизмов эволюции, опираясь на экспериментальное изучение “зависимости между экологической и генетической структурой природных популяций, с одной стороны, и их продуктивностью и приспособляемостью – с другой” [Там же, с. 174]. Следует поэтому согласиться с Ю.И. Черновым [41] в том, что становление эволюционной экологии в мире должно быть связано в первую очередь с именами С.А. Северцова, Д. Лэка и С.С. Шварца.
С.С. Шварц принял в качестве генетической основы эволюционно-экологических представлений базовые идеи СТЭ. Он понимал, что неодарвинизм не может объяснить многие ключевые аспекты эволюции, включая видообразование (поскольку его нельзя свести только к внутривидовой дифференциации), а также макроэволюцию. С.С. Шварц также считал, что “косвенная роль фенотипических механизмов в эволюции, вероятно, более серьезна, чем это представляется неодарвинизмом” [1, с. 12]. При этом он подчеркивал, что “… Невнимание синтетической теории эволюции к вопросам физиологии развития, фенотипической реализации генотипа, к эпигенетике следует, вероятно, также отнести к числу ее недостатков, но этот недостаток не может быть преодолен на основе ламарковских концепций. Наоборот, применение к решению его некоторых принципов экологии может оказаться очень плодотворным” [1, с. 12]. Неполное соответствие ЭЭ (в понимании ее С.С. Шварцем) постулатам неодарвинизма следует из его утверждения: “Вопрос "вид – не вид” решается на экологическом, а не на физиологическом или генетическом уровне" [47, с. 15]. Еще более строго это отражено в другом его утверждении: “…виды не потому виды, что они не скрещиваются, а они потому не скрещиваются, что они виды” [1, с. 149]. Из этих утверждений явно следует примат экологии, а не генетики в процессе видообразования. С его точки зрения именно экологические механизмы и факторы в первую очередь обусловливают этот процесс. Последнее явно отличалось от представлений неодарвинизма середины XX в.
Тем не менее С.С. Шварц в теоретических построениях предпочел опираться на СТЭ как на наиболее развитую в те годы эволюционную теорию. Не случайно, говоря о предыстории становления ЭЭ, он отметил большое значение для ее развития представлений об экологической генетике, развивавшихся И.М. Лернером [48] и Э.Б. Фордом [49]. Он полагал, что это направление наиболее приближается к сущности ЭЭ в его понимании, но тем не менее лежит в русле генетики, а не экологии. Поскольку наиболее важными ключевыми блоками эволюционно-экологической теории видообразования С.С. Шварца [1, 32, 47] были начальные процессы популяционных микроэволюционных явлений, то неодарвинизм был им принят в качестве теоретической базы ЭЭ. По С.С. Шварцу “Популяция – биологическое единство, генетическое и экологическое проявление которого взаимообусловлены. Взаимосвязь экологического и генетического в популяции – это тот фон, на котором развертываются элементарные эволюционные явления” [1, с. 18]. Поэтому он считал, что главная задача ЭЭ заключается во всестороннем исследовании движущих сил эволюции на основе ее экологических механизмов [1].
Дальнейшее развитие эволюционно-экологических взглядов связывают с синэкологическими идеями, предложенными Э. Пианкой [50], П. Джиллером [51] и М.Л. Розенцвейгом [52].
Интересно сопоставить общие эволюционно-экологические представления С.С. Шварца [1, 32] с версией ЭЭ, изложенной Э. Пианкой [50]. Главная особенность ЭЭ в представлении С.С. Шварца заключается в том, что она была нацелена на использование методов популяционной биологии для изучения процесса эволюции, создание экологически ориентированной новой эволюционной теории. Эта направленность актуальна и в наше время. Эволюционная экология Э. Пианки была направлена на применение эволюционных понятий для объяснения популяционных явлений, изучение функционирования и происхождения сообществ. Ю.И. Чернов пришел к заключению, что ЭЭ в представлении Э. Пианки по существу – общая экология, исходя из мысли, что “…В определенном смысле экология – вся эволюционная” [53, с. 85].
Поскольку важны не только популяционно-экологические, но и синэкологические аспекты, Ю.И. Чернов [41] предложил включить в содержание ЭЭ три главных направления: 1) экологические факторы микроэволюционного процесса и видообразования; 2) адаптациогенез (включая адаптивную радиацию и захват новых адаптивных зон); 3) ценотическая эволюция (=эволюция на надорганизменном уровне). В последнем случае он имел в виду такие экологические формы организации, как “биоценозы, сообщества, популяционные и социоэкологические структуры, коадаптивные комплексы, морфоадаптивные типы и жизненные формы”.
По мнению Ю. И. Чернова, “По С.С. Шварцу эволюционная экология – это преимущественно эволюционная популяционная экология, т.е. исследование экологических механизмов и факторов микроэволюционного процесса, который совершается в популяциях” [53, с. 88], т.е. относится к первому направлению. Однако с этим можно согласиться лишь отчасти, поскольку С.С. Шварц и его школа много внимания уделяли и исследованиям в русле второго направления – адаптациогенеза. При этом они опирались на изучение изменчивости морфологических и морфофизиологических признаков большого числа видов в природных условиях и на эксперименты по параллельному выращиванию и скрещиванию внутривидовых форм и таксономически спорных видов из разных географических мест и природных зон [47, 54–57]. Вопросы эволюции на биоценотическом уровне (третье направление ЭЭ по Ю.И. Чернову) также привлекали С.С. Шварца, но он еще не видел путей для реального решения этой проблемы и заключил, что “Эволюция организмов сопровождается изменением структуры и организации их сообществ, в конечном итоге – биосферы. Какова взаимосвязь между этими процессами? Автор не считает возможным даже подойти к решению этой проблемы во всей ее многогранности. Можно полагать, что время это еще не пришло. Но оно придет завтра” [58, с. 213]. Тем не менее С.С. Шварц пришел к выводу, что “…понятие “эволюция” нельзя ограничивать филогенезом отдельных видов или групп организмов, оно включает в себя и эволюцию природных сообществ, изменение фауны и флоры в целом, эволюцию биосферы. Эволюционный процесс в обычном смысле слова в значительной степени детерминируется эволюцией биогеоценозов” [58, с. 222]. Поэтому можно заключить, что все три указанных Ю.И. Черновым направления ЭЭ были намечены в работах С.С. Шварца и его научной школы.
Для развития ЭЭ важной вехой стала появившаяся в начале XXI в. книга М.Дж. Вест-Эберхард [59] с революционными для того времени представлениями о роли фенотипической пластичности, модификаций и быстрых развитийных (developmental) перестроек в эволюции. Важны также обобщения Е. Яблонки и М. Лэмб [14, 18, 60] о трансгенерационном наследовании вызванных стрессом эпигенетических перестроек генома. Новые представления о роли эпигенетической наследственности, обеспечивающей возможность быстрых перестроек морфогенеза, требуют пересмотра основных представлений не только в области теории эволюции, но и в сфере ЭЭ. С.С. Шварц также обращал внимание на особое значение анализа фенотипической изменчивости (модификаций) и процессов морфогенеза в эволюционно-экологических исследованиях [47]. Опираясь на идею стабилизирующего отбора И.И. Шмальгаузена [61], он сформулировал в ЭЭ оригинальную концепцию оптимального фенотипа [54].
На рубеже XX–XXI вв. ключевое значение для развития второго и третьего направлений ЭЭ (по Ю.И. Чернову) имели новые представления в области теории коэволюции, сформулированные Дж.Н. Томпсоном [62, 63]. Следующим этапом развития ЭЭ можно считать обоснование М.Л. Розенцвейгом [52] представлений об “экологии согласования”, или “экологии примирения” (Reconciliation ecology), нацеленной на оценку согласованности размеров видовых ареалов (SPAR’s – species-area relationships) и площади их естественных сообществ, сохранившихся в относительно неизменном виде в результате исторической деятельности человечества. Концепция М.Л. Розенцвейга должна учитываться при оценке устойчивости биоразнообразия и прогнозировании экологических кризисных явлений при перестройках региональных сообществ и экосистем.
Ключевое значение для ЭЭ в последние годы имеет развитие концепции расширенного эволюционного синтеза – РЭС (EES) [11, 64]. Эти представления влияют на понимание механизмов быстрой микроэволюции и видообразования, дополняя их представлениями об эпигенетическом наследовании (soft heredity) и механизмах активного формирования ниши живыми организмами (в соответствии с ТКН). Тем не менее я полагаю, что до настоящего расширенного синтеза еще далеко, поскольку авторы РЭС пока почти не учитывают или только упоминают влияние биотических сообществ на эволюцию их компонентов, которое ранее широко обсуждалось, например, В.В. Жерихиным [7], при рассмотрении механизмов филоценогенеза.
В заключение краткого очерка становления ЭЭ следует упомянуть книгу А.С. Северцова [65] – “Эволюционная экология позвоночных животных”, которая содержит новые материалы и представления. В книге полно представлены взгляды автора на истоки и сущность данного направления, доказывается необходимость поддержания внутривидового и видового разнообразия для обеспечения феномена “эволюционного стазиса” и связь эволюционных процессов с филоценогенезом. А.С. Северцов обосновал важное соображение о том, что филоценогенез осуществляется не столько за счет специогенеза и экогенеза, сколько за счет смены жизненных форм и биоморфогенеза, необходимых для освоения новых адаптивных зон.
КОНЦЕПТУАЛЬНОЕ ПРОСТРАНСТВО ЭВОЛЮЦИОННОЙ ЭКОЛОГИИ
Эволюционная экология – часть экологии, но ее концептуальное пространство определено недостаточно строго. Разночтения есть и в видении концептуального пространства самой экологии. Д.Н. Кашкаров [35] выделял в экологии две стороны – аутэкологию и синэкологию. Н.П. Наумов [66] считал, что подразделение на аутэкологию и синэкологию не строгое и выделял экологию особей, популяций и сообществ (=биоценологию). С.С. Шварц [1, 32, 58] выделял аутэкологию, популяционную экологию, синэкологию и биогеоценологию. А.С. Северцов считал, что “Взаимодействия организмов с окружающей средой можно изучать на уровне особи – аутэкология или экофизиология, на уровне популяций и видов – популяционная экология или синэкология, на уровне экосистем – биоценология, на уровне всей биосферы” [65, с. 4]. И.А. Шилов попытался построить трехмерную схему экологии, где популяция была выделена в качестве начала координат [67, с. 241]. Вдоль одной оси им было показано направление от организма к популяции, вдоль второй – от популяции к биоценозу и биогеоценозу (=экосистеме), а вдоль третьей оси – от популяции к виду и далее к Царству. Особое место в этом пространстве занимала Биосфера, к которой тянутся связи от биогеоценозов, с одной стороны, и Царств, с другой. И.А. Шилов полагал, что популяция при этом выступает в двух ипостасях: 1) как биохорологическая единица (по Н.П. Наумову) и 2) как эволюционная единица (по С.С. Шварцу).
Эволюционная экология оперирует в качестве объектов исследований популяциями и видами, рассматривая их в разных условиях среды. Попытаемся построить трехмерное концептуальное пространство ЭЭ, используя три основные переменные: 1) популяции (число изучаемых популяций), 2) виды (число изучаемых видов) и 3) среда (градиент условий (состояний) среды) (рис. 1). На рисунке вдоль первой оси при изучении разного числа популяций фактически описывается направление, которое соответствует популяционной экологии (population ecology) в широком толковании. Вдоль второй оси для множества видов будет описываться направление, характеризующее синэкологию (synecology). Вдоль третьей оси помещаются состояния среды, по реакции на которые характеризуется направление исследований, соответствующее аутэкологии (autecology).
Рис. 1.
Концептуальное пространство эволюционной экологии и основных ее научных направлений в виде трехмерной модели (пояснения см. в тексте).
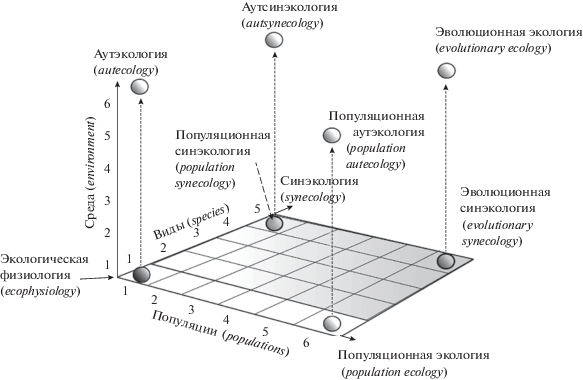
В начале координат, т.е. в нулевой позиции, мы поместили организм (см. рис. 1), поэтому если взять особи из определенной популяции какого-либо вида и оценить их физиологическую реакцию на изменение среды (вдоль третьей оси), то такое исследование будет соответствовать задачам экологической физиологии (ecophysiology) и/или организменной аутэкологии (organismic autecology). С момента возникновения термина аутэкология (от греч. autos – сам) его связывают как с изучением пределов условий, которые организмы выбирают для своего существования, так и с характеристикой взаимоотношений особей со средой. Поэтому нет противоречия в том, чтобы изучались аспекты аутэкологии не только отдельных особей, но и групп особей – популяций или сообществ, т.е. как самих надорганизменных систем. Это позволяет осознанно применять термин аутсинэкология для характеристики взаимоотношений со средой локальных популяций симпатрических видов, входящих в сообщество (таксоцен).
Рассмотрим (см. рис. 1) ситуацию, когда взят только один вид, одно состояние среды и одна популяция этого вида. В такой модели в одном месте и за короткий срок при сходных условиях среды будет изучаться разнообразие реакций особей из конкретной популяции. Такое исследование относится к популяционной экологии (population ecology). Однако изучение разнообразия реакций особей внутри популяции точнее будет характеризоваться как внутрипопуляционная экология (intrapopulation ecology) и фактически соответствует термину демэкология, которым иногда заменяют понятие популяционная экология. Из данного примера хорошо понятно, что так делать не следует, поскольку популяции состоят из элементарных поселений или демов (= микропопуляций). Двигаясь вдоль третьей оси, мы можем, например, рассмотреть сезонные или межгодовые изменения данной популяции, т.е. когда условия среды изменяются. Тогда это будет аутэкологическое популяционное исследование или точнее – популяционная аутэкология (population autecology). Если вдоль первой оси мы сравним несколько популяций одного вида, но в сходных биотопических условиях, причем за короткий срок (например, сравнение популяций рыжей полевки вдоль пойменных лесов реки Урал в июле), то исследование будет проводиться в рамках межпопуляционной экологии (interpopulation ecology). Усложнив модель за счет смещения вдоль третьей оси, получим ситуацию, когда изучаются несколько удаленных в пространстве и времени (например, по сезонам) популяций одного вида. Анализируя реакцию разных популяций одного вида на сезонные изменения среды, будем проводить исследование в рамках межпопуляционной аутэкологии (interpopulation autecology).
Теперь рассмотрим другую ситуацию, когда синтопно и синхронно изучены по одной ценопопуляции у нескольких видов, населяющих тот же биотоп в одном локалитете и в один сезон. При этом исследуется синэкологическая задача о том, как реагируют ценопопуляции разных симпатрических видов на одни и те же условия (например, засуху). Такое исследование лежит в русле направления, которое мы предлагаем назвать популяционная синэкология (population synecology). Общий замысел подобных исследований может быть иным. Например, мы можем изучить ценопопуляцию одного вида в разные годы при разном уровне численности и разной полноте видового состава таксоцена. При изучении реакции вида-доминанта Clethrionomys glareolus на высокую и низкую численность популяции и на разный видовой состав (олиго- и поливидовой) таксоцена в разные годы были обнаружены резкое переключение морфогенеза и изменение формы нижней челюсти при разных констелляциях численности и полноты видового состава таксоцена [68]. При дополнительном учете смещения вдоль третьей оси, т.е. изучении реакций ценопопуляций разных видов на разные условия среды (сезоны, годы, антропогенные воздействия), такое исследование (взаимодействия “вид × среда”) будет соответствовать популяционной аутсинэкологии (population autsynecology).
Представим далее ситуацию, когда относительно синхронно и синтопно анализируются ценопопуляции нескольких видов (например, землероек или грызунов) из географически удаленных локалитетов. Эти данные характеризуют задачу, которая позволяет выявить сопряженные реакции видов таксоцена на изменение экологических условий в удаленных биоценозах, оценить их коадаптивные свойства [69]. Такая ситуация соответствует направлению, которое я предлагаю называть эволюционная синэкология (evolutionary synecology). Если к этой задаче добавить анализ изменений среды вдоль третьей переменной, что происходит при изучении географически удаленных сообществ (таксоценов), то данное исследование будет формально соответствовать эволюционной аутсинэкологии (evolutionary autsynecology). Аспекты популяционной и эволюционной синэкологии с учетом аутсинэкологии отражают основную проблематику эволюционной экологии (evolutionary ecology) в ее широком понимании [41, 53].
В общей 3D модели отсутствует 4-е измерение – время, но изменение условий среды во времени отчасти компенсирует этот недостаток. Нетрудно представить, что при мысленном дополнении данной модели 4-й переменной времени 3D-схема анимируется: “популяции” будут возникать как шарики, флуктуировать по размерам (численности) и дрейфовать вдоль третьей оси (в разных условиях) и т.д.
Таким образом, данная 3D модель хорошо описывает концептуальное пространство эволюционной экологии и основных составляющих ее дисциплин: популяционной экологии, популяционной аутэкологии, популяционной и эволюционной синэкологии, а также популяционной и эволюционной аутсинэкологии. Дополнения в виде терминологической приставки аут- (aut-) в последних двух случаях, вероятно, не требуются, поскольку трудно представить полностью идентичные экологические условия при исследованиях в рамках популяционной и эволюционной синэкологии. Можно заключить, что концептуальное пространство эволюционной экологии действительно включает аутэкологические, популяционно-экологические и синэкологические компоненты исследований и все их возможные композиции.
Включение аспектов популяционной и эволюционной синэкологии в область интересов ЭЭ позволяет перейти к новым направлениям и возможностям исследований. Часть таких аспектов ЭЭ уже намечена. Известны: изучение разных аспектов коэволюции и, в частности, взаимной диффузной коэволюции видов в сообществе [62, 63]; анализ эволюционно-экологических механизмов симпатрического формообразования [70], включая быстрое возникновение и дифференциацию флоков (flocks) рыб [71, 72]; выявление обратных связей (feedbacks) между экологическими и эволюционными событиями в исторические характерные времена [13, 73]; изучение быстрых микроэволюционных событий в результате биологических инвазий [74, 75] или хронического антропогенного воздействия [76]; сопоставление действия принципа компенсации Ю.И. Чернова на разных уровнях биологической организации [68, 77]; сравнительный филогенетический анализ сообществ и путей их формирования [78]; решение задач урбанистической эволюционной экологии [13, 79] и др.
В целом для ЭЭ актуальна проблема быстрых морфогенетических перестроек популяций и сообществ при стрессирующих воздействиях, а также анализ быстрых перестроек морфогенеза на основе трансгенерационного наследования индуцированных стрессом эпигенетических изменений [14, 20]. В рамках данной статьи нет возможности рассмотреть весь комплекс быстро развивающихся эволюционно-экологических исследований, поэтому я остановлюсь лишь на некоторых из них.
ПОПУЛЯЦИОННАЯ И ЭВОЛЮЦИОННАЯ СИНЭКОЛОГИЯ
Проблемы синэкологии долго были вне интересов популяционных и эволюционных биологов, хотя эволюционные процессы осуществляются в сообществах и во многом, если не всегда, ими контролируются и направляются [7, 32, 53]. Важнейшей задачей ближайшего будущего следует считать внедрение в синэкологию [76, 80]: 1) популяционных представлений и методов, обеспечивающих переход к популяционной и эволюционной синэкологии (population and evolutionary synecology); 2) методов двухуровневой оценки экологического состояния ценопопуляций симпатрических видов и их сообществ, нацеленных на разработку популяционно-ценотических (population-coenotic) представлений. Необходимо также прогнозирование наступления кризисных ценотических явлений.
По моим представлениям, локальные сообщества таксономически близких симпатрических видов в пределах фации или урочища являются таксоценами (термин Г.Е. Хатчинсона [38]), т.е. таксономически близкими компонентами ценозов, выполняющими сходные, главным образом трофические и средопреобразующие функции. Таксоцены – это филогенетически близкородственные экологические гильдии, фрагменты сообществ, которые могут рассматриваться как их модели. Синтопные поселения каждого вида в таксоцене, населяющие локальный биотоп для относительно оседлых видов (например, моллюсков, многих групп насекомых, землероек, грызунов), – это ценопопуляции, которые экологически взаимодействуют друг с другом, поскольку обитают на одной территории и используют сходные ресурсы.
Термин ценопопуляция широко используется ботаниками и означает территориальную группировку вида, приуроченную к определенному биоценозу и соответственно биотопу. Зоологи предпочитают другие термины: дем, парцелла, локалитет или поселение. Подобные небольшие территориальные группировки, потенциально или реально связанные друг с другом мигрантами и формирующие единую популяцию (=метапопуляцию), принято также называть микропопуляциями [1, 32]. Они характеризуют только часть (территориальный фрагмент) популяции конкретного вида и формально не привязаны к биоценозу. Поэтому при сравнении особей из синтопных поселений симпатрических и таксономически близких видов, синхронный сбор которых приурочен к одним и тем же биотопам, мы сознательно используем по отношению к таким локальным группировкам термин ценопопуляция. В составе таксоцена ценопопуляции каждого вида населяют общий биотоп и одновременно могут рассматриваться как микропопуляции. Однако при оперировании выборками из синтопных группировок нескольких симпатрических видов, совпадающих в биотопическом и территориальном отношениях, только термин ценопопуляция представляется оправданным, а применение к ним территориального термина микропопуляция в этом случае теряет смысл. Если мы работаем исключительно с одним видом, то его локальные биотопические поселения логичнее называть микропопуляциями или демами.
Синхронный внутри- и межгрупповой анализ синтопных ценопопуляций симпатрических видов, входящих в таксоцен, позволяет в русле популяционной синэкологии сопоставить их реакции в виде увеличения изменчивости признаков в ответ на изменение условий. Он дает возможность оценить параллелизм или независимость проявления внутри- и межгрупповой сопряженной изменчивости морфогенетических, физиологических и поведенческих реакций, т.е. коэволюционный потенциал симпатрических видов [69]. Противоположные реакции могут указать на антагонизм экологических требований видов, а параллелизм ответов – на высокий коадаптивный потенциал, т.е. на общую адаптивную морфогенетическую и морфофизиологическую реакции. Анализ внутригруппового разнообразия позволяет оценить устойчивость ценопопуляции к констелляциям условий среды в разные сезоны и годы. Для нескольких синтопных и синхронно оцениваемых ценопопуляций симпатрических видов локального таксоцена тем же способом можно оценить изменение общего таксоценотического разнообразия во времени. При параллельном изучении нескольких таксоценов, включающих ценопопуляции одних и тех же видов, в географически удаленных локалитетах, т.е. в разных условиях, проводится аналогичное сравнение, но в этом случае уже не аллохронных, а аллотопных выборок из ценопопуляций нескольких видов. Например, с помощью методов геометрической морфометрии [69] можно разместить в общем морфопространстве ординаты особей разных видов, анализируя только изменчивость формы объектов.
Совмещение всех задач, т.е. параллельное сравнение географически удаленных, но синтопных, ценопопуляций нескольких симпатрических видов во времени и в пространстве относится уже к проблематике эволюционной синэкологии и/или эволюционной экологии в ее широком понимании [41]. При таком комплексном сопряженном анализе изменчивости свойств фенома в его самом широком толковании (от морфологических признаков до особенностей поведения особи на разных этапах онтогенеза) появляется возможность оценить, какой из видов-симпатриантов лучше адаптирован к условиям локального биотопа по проявлению изменчивости и разнообразия изученных признаков. В неблагоприятных для вида условиях его изменчивость по отдельным признакам или внутригрупповое разнообразие, оцененное по их совокупности, будут возрастать, а в благоприятных, напротив, уменьшаться [80, 81].
Феномен увеличения веера изменчивости признаков в неблагоприятной среде был экспериментально установлен и описан Н.В. Глотовым как эффект провокационного фона среды [82]. Мы предлагаем его назвать “принципом Н.В. Глотова”. С его помощью можно определить ценопопуляции видов с разным уровнем морфоразнообразия (morphological disparity). Вывод может быть получен после оценки степени рассеивания ординат в многомерном морфопространстве на основе методов геометрической морфометрии [83] при случайном выравнивании выборок по объему (процедура рарефакции). Например, установлено, что морфоразнообразие видов-доминантов значимо меньше, чем у видов-субдоминантов [80, 85]. При естественном восстановлении сообщества грызунов после неизбирательной элиминации на освобожденной территории у вида, бывшего в контроле малочисленным субдоминантом, на импактном участке значимо снизился уровень внутригруппового морфоразнообразия; это отражает снижение стрессированности морфогенеза. В другом исследовании [69] при сравнении сопряженной географической изменчивости трех симпатрических видов землероек рода Sorex у них проявился географический параллелизм трансформаций морфогенеза; это позволило с вероятностью выше 95% дискриминировать особей разных таксоценов независимо от их видовой принадлежности. Выявлено также снижение морфоразнообразия популяции вида-доминанта при наибольшей возможной полноте видового состава таксоцена, но значимое возрастание данного показателя при его неполноте [68]. Эти факты указывают на реальность влияния взаимодействий симпатрических видов локального сообщества на процессы морфогенеза и динамику как ценопопуляций, так и таксоцена.
Таким образом, приложение методов из арсенала популяционной биологии к задачам популяционной и эволюционной синэкологии позволяет оценить экологическое состояние как отдельных ценопопуляций симпатрических видов, так и локальных таксоценов. Развивая эту методологию, можно приблизиться к выявлению и прогнозированию региональных биоценотических кризисов.
ЭКСПЕРИМЕНТАЛЬНАЯ ЭВОЛЮЦИОННАЯ ЭКОЛОГИЯ
Объединение двух направлений – экспериментальных методов систематики, обсуждавшихся С.С. Шварцем [47, 86] с позиций эволюционной экологии, и экспериментальной экологии, о которой писали А. В. Покровский и В.Н. Большаков [87], – приводит к новому направлению, которое я предлагаю назвать экспериментальная эволюционная экология (ЭЭЭ). Однако ни С.С. Шварц, ни А.В. Покровский, ни В.Н. Большаков не писали об этом направлении и не применяли такой терминологии, хотя она во многом подразумевалась и непосредственно вытекает из многих их работ.
Идея была проста, но на практике работала эффективно. Если выращивать в сходных лабораторных условиях вивария представителей разных таксонов, взятых из разных условий или географических точек (например, из лесостепи и лесотундры), то морфофизиологическое или иное своеобразие их потомков будет отражать степень эволюционной дивергенции таксонов. Если межгрупповая изменчивость не проявится, это будет указывать на эволюционно-экологическую и феногенетическую близость сравниваемых форм, их историческую и филетическую общность.
В экспериментальной эволюционной экологии, как я полагаю, выделяются три основных направления: 1) экспериментальная оценка степени эволюционной дивергенции форм в лабораторных условиях (можно решать задачи экспериментальной систематики и эволюционно-экологические); 2) популяционно-экологические эксперименты в лаборатории и/или в природе (задачи из области популяционной экологии конкретных видов и эволюционно-экологических); 3) популяционно-ценотические “эксперименты” и слежение за реакциями видов и таксоценов. В случае изучения изменчивости и/или морфоразнообразия на уровне таксоценов проводится обобщенный анализ объектов без учета их видовой принадлежности, т.е. в единой совокупности синхронно полученных природных выборок симпатрических синтопных видов, случайным способом выравненных по объему наблюдений.
Рассмотрим конкретный пример, относящийся к ЭЭЭ. В виварии ИЭРиЖ УрО РАН под руководством А.В. Покровского [87] параллельно создали лабораторные колонии двух видов полевок – узкочерепной и полевки-экономки, каждый из которых был представлен двумя подвидами: северным и южным. Виды на значительной части их ареалов симпатрируют, причем симпатрическими формами являются северные и южные подвиды обоих видов. Поскольку животных разводили в выравненных условиях, можно было оценить их морфогенетическое своеобразие по комплексу 46 фенов неметрических признаков [88]. Дискриминантный канонический анализ значений главных компонент индивидуальных фенетических композиций северного (Lasiopodomys gregalis major) и южного (L. g. gregalis) подвидов узкочерепной полевки с северным (Alexandromys oeconomus hahlovi) и южным (A. o. oeconomus) подвидами полевки-экономки выявил значимые различия вдоль всех трех осей. Вдоль первой дискриминантной канонической функции проявились отчетливые межвидовые различия, а вдоль второй функции наблюдался однонаправленный параллельный сдвиг эллипсоидов, характеризующий изменчивость проявления фенетических композиций обоих южных подвидов по отношению к эллипсоидам обеих северных форм [88, см. рис. 1]. Другими словами, вдоль второй оси у обоих видов при их продвижении на юг и север наблюдаются во многом параллельные структурные изменения в проявлении целого ряда гомологичных фенов неметрических признаков.
Проявление фенов устойчиво к прямому действию различных экологических факторов. Это позволило использовать внутрииндивидуальную (феногенетическую) изменчивость дискретных морфоструктур для косвенной оценки уровня эпигенетических различий между сравниваемыми группами животных – меры их эпигенетической дивергенции [89]. Уровень эпигенетической дивергенции оценили как квадрат обобщенного расстояния Махаланобиса. Мера эпигенетической дивергенции аддитивна и включает в себя как минимум две компоненты: филогенетическую и эволюционно-экологическую. Филогенетическая компонента составила приблизительно 76% общей межгрупповой изменчивости. Эволюционно-экологическая компонента объединила приблизительно 24% межгрупповой дисперсии и подразделилась на видоспецифическую (взаимодействие “таксон” × “среда обитания”) – 9% и эколого-историческую – около 15% (параллелизм проявления фенов как результат исторического освоения таксонами сходных экологических условий). Филогенетическая компонента изменчивости многократно превысила эволюционно-экологическую. Но проявление существенной эколого-исторической компоненты указывает на то, что у обоих видов выработались сходные необратимые морфогенетические различия между северными и южными подвидами. Последнее может быть истолковано как проявление у симпатрических видов параллельных микроэволюционных перестроек морфологических структур осевого черепа и нижней челюсти, носящих в основном направленный адаптивный характер.
Популяционно-экологические эксперименты в лабораторных условиях и/или в природе как второе направление исследований в русле ЭЭЭ подразумевают следующие аспекты. Длительное слежение за популяциями в природных условиях, основанное на периодическом изъятии особей, мало отличается от аналогичного мониторинга в лабораторных условиях. Сочетание обеих техник популяционного мониторинга, т.е. параллельного анализа одновозрастных особей из природных популяций и происходящих от них лабораторных групп, позволяет получить наиболее интересные и содержательные результаты как эволюционной, так и популяционной экологии, поскольку в лабораторных условиях снимаются все межвидовые (ценотические) взаимодействия, упрощаются внутрипопуляционные взаимодействия, а также обеспечивается избыток ресурсов, что моделирует иную экологическую среду, чем в природе.
Преимущества слежения за природными популяциями, например животных, состоят в том, что особи находятся в естественной среде, имеются внутрипопуляционные и ценотические взаимодействия, питание, как правило, разнообразно и полноценно, но корм не является регулярно избыточным. Поскольку природные условия неконтролируемы, требуется длительно собирать материал в расчете на обнаружение сходных природных ситуаций. Если требуется выявить в популяции морфогенетическую реакцию разных структурно-функциональных групп – биотипов или морф [90] на те или иные условия среды в расчете на самый широкий их диапазон, нужно лишь обеспечить получение синхронных и синтопных выборок [91].
Популяционно-ценотические “эксперименты” и мониторинг таксоценов – еще одно направление в русле ЭЭЭ. Особый интерес в этом отношении представляет анализ разных форм сопряженной изменчивости симпатрических видов не в лабораторных, а в природных условиях. Это могут быть проявления сопряженной географической, хронографической или биотопической изменчивости у разных видов одного таксоцена. Параллельный анализ морфологической изменчивости синхронных выборок симпатрических видов, населяющих локальный биотоп, соответствует приведенному выше примеру параллельного изучения морфогенетических реакций разных форм в одних и тех же лабораторных условиях. Поэтому изучение синтопных аллохронных выборок представителей симпатрических видов – это прямая аналогия лабораторного сравнения, которое перенесено в естественные природные условия. Наиболее простым аналогом лабораторных исследований мониторинговым наблюдениям в природных условиях может быть сбор данных по аллохронным синтопным выборкам симпатрических видов в естественной среде, который регулярно (ежегодно) проводится в один и тот же сезон и временной отрезок [80]. Таким образом, простой параллельный мониторинг симпатрических видов таксоцена позволяет оценить эволюционно-экологические механизмы межвидовых взаимодействий и синэкологическую природу коэволюционных перестроек морфогенеза в сообществе.
ЗАКЛЮЧЕНИЕ
Академик С.С. Шварц, развивая полвека назад представления об эволюционной экологии, наметил широкий круг задач и общий путь исследований, который во многом актуален и сегодня в год его 100-летия [10, 12]. Несмотря на то, что теоретические представления о механизме видообразования, выдвинутые С.С. Шварцем [1, 32], должны быть сегодня откорректированы с учетом новых открытий, его эволюционно-экологическая основа изменилась не сильно. Мне представляется, что этапы видообразования, основанные по С.С. Шварцу на возникновении необратимых морфофизиологических особенностей, изменяющих отношение популяций к среде, а также включающие тканевые адаптации, напрямую связаны с экологическими эпигенетическими механизмами, широко обсуждаемыми в русле РЭС [16, 19, 20, 91]. Идея С.С. Шварца о регулярных гомеостатических колебаниях генетической структуры, объясняющая регулярные сезонные фенотипические перестройки, должна быть заменена представлениями об известных теперь гомеостатических перестройках эпигенетической системы, фенотипической пластичности и пороговых переключениях морфогенетических программ [18, 59, 60, 68]. Роль экологических механизмов в эволюционных изменениях, связанных с перестройками экологической структуры популяций, еще только начинает осознаваться [11, 19, 64].
Полагаю, что в связи с концепцией расширенного эволюционного синтеза экологические механизмы эволюции, намеченные С. С. Шварцем, следует дополнить. Необходимо изучение как эпигенетических последствий изменений соотношения всех структурно-функциональных групп в популяции, так и изменений соотношения видов в сообществе (таксоцене), а также обратных связей между экологическими и эволюционными процессами в популяциях и сообществах [13, 73, 92]. Рассмотренная С.С. Шварцем [1, 32] связь между микро- и макроэволюционными процессами по-прежнему широко обсуждается, но еще не решена [93]. Тем не менее с позиций РЭС и ЭТЭ ее решение представляется вполне реалистичным [92].
Можно полагать, что в недалеком будущем среди прочих направлений ЭЭ будет возрастать значение эволюционной синэкологии (ЭС), контуры которой еще только намечаются. Именно ЭС будет нацелена на изучение и моделирование коэволюции сообществ, выяснение механизмов симпатрического формообразования и прогнозирование быстрых перестроек популяций и сообществ. Основное преимущество ЭС – двухуровневый популяционно-ценотический подход [53, 81] к проявлениям сопряженной изменчивости и морфоразнообразия ценопопуляций в локальных таксоценах [80]. Представления экспериментальной эволюционной экологии хорошо согласуются с подходами ЭС и эпигенетической концепцией быстрых морфогенетических перестроек с позиций РЭС [94, 95], а также с представлениями о механизмах диффузной коэволюции [62, 63] и могут быть рассмотрены как особое методологическое направление ЭЭ.
Предложенные подходы к двухуровневому популяционно-ценотическому анализу симпатрических видов, в том числе на основе популяционной и эволюционной синэкологии, молекулярной генетики и геометрической морфометрии, перспективны для оценки экологического состояния и морфогенетических перестроек отдельных ценопопуляций и локальных таксоценов. Дальнейшее развитие эволюционно-экологических представлений о новых путях и методах прогноза региональных кризисных биоценотических явлений и экологических эпигенетических механизмах популяционно-ценотических перестроек позволит приблизиться к практическому решению задач управления начальными процессами эволюции, а также направленного изменения структуры и функциональных свойств ценозов [52]. Во многом это согласуется с высказанными С. С. Шварцем ожиданиями [1, 32] и даже отчасти их превосходит в отношении возможного конструирования гибридных (рекомбинантных) ценозов [3], включающих чужеродные инвазионные виды [74].
Обзор литературы и подготовка рукописи выполнены в рамках государственного задания Института экологии растений и животных УрО РАН, а также при частичной финансовой поддержке Комплексной программы исследований УрО РАН (проект № 18-4-4-28).
Автор признателен д.б.н. И. А. Васильевой за полезное обсуждение рукописи статьи и рецензенту за высказанные конструктивные критические замечания.
Список литературы
Шварц С.С. Эволюционная экология животных. Экологические механизмы эволюционного процесса. Свердловск, 1969. 198 с.
Павлов Д.С., Букварева Е.Н. Биоразнообразие, экосистемные функции и жизнеобеспечение человечества // Вестник РАН. 2007. Т. 77. № 11. С. 974–986.
Rotherham I.D. Recombinant ecology – a hybrid future? Sheffield, UK: Springer Briefs in Ecology, 2017. 85 p. doi 10.1007/978-3-319-49797-6_1
Crutzen P.J., Stoermer E.F. The Anthropocene // Global Change News. 2000. V. 41. P. 17–18.
Steffen W., Grinevald J., Crutzen P., McNeil J. The Anthropocene: conceptual and historical perspectives // Philosophical Transactions of The R. Soc. A. 2011. V. 369. P. 842–867.
Zalasiewicz J., Williams M., Steffen W., Crutzen P. The new world of the Anthropocene // Environ. Sci. Technol. 2010. V. 44. P. 2228–2231.
Жерихин В.В. Избранные труды по палеоэкологии и филоценогенетике. М.: Тов-во научн. изд. КМК, 2003. 542 с.
Moyne S., Neige P. The space-time relationship of taxonomic diversity and morphological disparity in the Middle Jurassic ammonite radiation // Palaeogeography, Palaeoclimatology, Palaeoecology. 2007. V. 248. P. 82–95.
Ivits E., Cherlet M., Mehl W., Sommer S. Ecosystem functional units characterized by satellite observed phenology and productivity gradients: A case study for Europe // Ecological Indicators. 2013. V. 27. P. 17–28.
Read A.F., Clark J.S. The next 20 years of ecology and evolution // Trends in Ecology and Evolution. 2006. V. 21. № 7. P. 354–355.
Schoener T.W. The newest synthesis: understanding the interplay of evolutionary and ecological dynamics // Science. 2011. V. 331. P. 426–429.
Sutherland W.J., Freckleton R.P., Goodfray H.Ch.J. et al. Identification of 100 fundamental ecological questions // J. Ecology. 2013. V. 101. P. 58–67.
Alberti M. Eco-evolutionary dynamics in an urbanizing planet // Trends in Ecology and Evolution. 2015. V. 30. № 2. P. 114–126.
Jablonka E., Lamb M.J. Transgenerational epigenetic inheritance // Evolution: the extended synthesis. Eds. Pigliucci M., Müller G.B. Cambridge MA: MIT Press, 2010. P. 137–174.
Bonduriansky R. Rethinking heredity, again // Trends in Ecology and Evolution. 2012. V. 27. № 6. P. 330–336.
Burggren W. Epigenetic inheritance and its role in evolutionary biology: re-evaluation and new perspectives // Biology. 2016. V. 5. № 24. P. 2–22.
Waddington C.H. Canalization of development and the inheritance of acquired characters // Nature. 1942. V. 150. P. 563–565.
Jablonka E., Lamb M.J. Epigenetic inheritance and evolution // Trends in Ecology and Evolution. 1996. V. 11. P. 266–267.
Ledón-Rettig C.C. Ecological epigenetics: an introduction to the symposium // Integrative and Comparative Biology. 2013. V. 53. P. 307–318.
Duncan E.J., Gluckman P.D., Dearden P.K. Epigenetics, plasticity and evolution: How do we link epigenetic change to phenotype? // J. Exp. Zool. Part B. Molecular and Developmental Evolution. 2014. V. 322. P. 208–220.
Шишкин М.А. Эпигенетическая система как объект селективного преобразования // Морфология и эволюция животных. М.: Наука, 1986. С. 63–73.
Шишкин М.А. Эволюция как эпигенетический процесс // Современная палеонтология. Т. 2. Общие закономерности эволюции органического мира. М.: Недра, 1988. С. 142–168.
Шмальгаузен И.И. Организм как целое в индивидуальном и историческом развитии. М.;Л.: Изд-во АН СССР, 1938. 144 с.
Шмальгаузен И.И. Стабилизирующий отбор и его место среди факторов эволюции // Журн. общ. биол. 1941. Т.2. № 3. С.307–354.
Waddington C.H. The epigenotype // Endeavour. 1942. V. 1. P. 18–20.
Dickins T.E., Rahman Q. The extended evolutionary synthesis and the role of soft inheritance in evolution // Rroceedings of The Royal Society. B. 2012. V. 279. P. 2913–2921.
Dupont C., Armant D.R., Brenner C.A. Epigenetics: definition, mechanisms and clinical perspective // Semin. Reprod. Med. 2009. V. 27. P. 403–408.
Bilichak A., Kovalchuk I. Transgenerational response to stress in plants and its application for breading //J. Exp. Biol. 2016. V. 67. P. 2081–2092.
Pigliucci M. Do we need an extended evolutionary synthesis? // Evolution. 2007. V. 61. № 2. P. 2743–2749.
Laland K., Matthews B., Feldman M.W. An introduction to niche construction theory // Evolutionary Ecology. 2016. V. 30. P. 191–202.
Laland K.N., Odling-Smee F.J., Feldman M.W. Evolutionary consequences of niche construction and their implications for ecology // Proceedings of the National Academy of Sciences. 1999. V. 96. № 18. P. 10 242–10 247.
Шварц С.С. Экологические закономерности эволюции. М.: Наука, 1980. 277 с.
Petrusewicz K. Teoria ewolucji Darwina jest teoria evologiczna // Ecol. Polska. 1959. Seria B. 5. T. 5. № 4. S. 297–263. [Petrusewicz K. Darwin’s evolution theory is an ecologie one // Ekol. Polska. 1959. V. 5. № 4. P. 297–263]
Кашкаров Д.Н. Среда и сообщество (основы синэкологии). М.: Гос. мед. изд-во, 1933. 244 с.
Elton C. Animal ecology and evolution. Oxford, 1930. 96 p.
Северцов С.А. Динамика населения и приспособительная эволюция животных. М.; Л.: Изд-во АН СССР, 1941. 316 с.
Северцов С.А. Проблемы экологии животных. Т. 1. М.: Изд-во АН СССР, 1951. 171 с.
Hutchinson G.E. Concluding remarks // Cold Spring Harb. Symp. Quant. Biol. 1957. V. 22. P. 415–427.
Hutchinson G.E. The ecological theater and evolutionary play. New Haven, Conn.: Yale Univ. Press, 1965. 139 p.
MacArthur R.H. The theory of island biogeography. Princeton, New Jersey: Princeton Univ. Press, 1967. 203 p.
Чернов Ю.И. Эволюционная экология – сущность и перспективы // Успехи современной биологии. 1996. Т. 116. Вып. 3. С. 277–291.
Беклемишев В.Н. О принципах сравнительной паразитологии в применении к кровососущим членистоногим // Мед. паразитол. и паразитар. болезни. 1945. Т. 14. Вып. 1. С. 3–11.
Мирзоян Э.Н. К истории глобальной экологии. Концепция Геомериды В.Н. Беклемишева. М.: Экологический центр ИИЭТ РАН, 2007. Вып. 1. 128 с.
Shvarts S.S. The evolutionary ecology of animals. Ecological mechanisms of the evolutionary process. N.Y.: Consultants Bureau, 1977. 292 p.
Orians G.H. Natural selection and ecological theory // The Amer. Naturalist. 1962. V. 96. No. 890. P. 257–263.
Lack D.J. Evolutionary Ecology // Anim. Ecol. 1965. V. 53. № 2. P. 237–245.
Шварц С.С. Проблема вида и новые методы систематики // Экспериментальные исследования проблемы вида. Свердловск, 1973. С. 3–18.
Lerner I.M. Ecological genetics (synthesis) // Genetics Today. V. 2. New York: Pergamon Press. Elmsford, 1965 [цит. пo Shvarts S.S., 1977].
Ford E.B. Ecological Genetics. London: Methuen, 1964. 493 p.
Пианка Э. Эволюционная экология. М.: Мир,1981. 399 с. [Pianka E.R. Evolutionary ecology. Sec. ed. New York, Hagerstown, San Francisco, London: Harpe and Row, Publishers. 1978]
Джиллер П. Структура сообществ и экологическая ниша. М.: Мир, 1988. 184 с.
Rosenzweig M.L. Reconciliation ecology and the future of species diversity // Oryx. 2003. V. 37. № 2. P. 194–205.
Чернов Ю.И. Экология и биогеография. Избранные работы. М.: Тов-во научн. изд. КМК, 2008. 580 с.
Шварц С.С. Принцип оптимального фенотипа // Журн. общ. биол. 1968. Т. 29. № 1. С. 12–24.
Шварц С.С., Смирнов В.С., Добринский Л.Н. Метод морфофизиологических индикаторов в экологии наземных позвоночных. Свердловск: УФАН СССР, 1968. 387 с.
Большаков В.Н. Пути приспособления мелких млекопитающих к горным условиям. М.: Наука,1972. 200 с.
Покровский А.В., Большаков В.Н. Экспериментальная экология полевок. М.: Наука, 1979. 147 с.
Шварц С.С. Эволюция и биосфера // Проблемы биогеоценологии. М.: Наука, 1973. С. 213–228.
West-Eberhard M.J. Developmental plasticity and evolution. Oxford: Oxford University Press, 2003. 816 p.
Jablonka E., Lamb M.J. Evolution in four dimensions. Genetic, epigenetic, behavioral, and symbolic variation in the history of life. The MIT Press: Cambridge, Massachusetts, London, England, 2005. 462 p.
Шмальгаузен И.И. Стабилизирующий отбор и его место среди факторов эволюции // Журн. общ. биол. 1941. Т. 2. № 3. С. 307–354.
Thompson J.N. Rapid evolution as an ecological process // Trends Ecol. and Evol. 1998. V. 13. P. 329–332.
Thompson J.N. Mutualistic webs of species // Science. 2006. V. 312. P. 372–373.
Laland K.N., Uller T., Feldman M.W., Sterelny K. et al. The extended evolutionary synthesis: its structure, assumptions and predictions // Philos. Trans. R. Soc. B.: Biol. Sci. 2015. V. 282. P. 1–14. doi 10.1098/rspb.2015.10
Северцов А.С. Эволюционная экология позвоночных животных. М.: Тов-во научн. изд. КМК, 2013. 347 с.
Наумов Н.П. Экология животных. М.: Высшая школа, 1963. 618 с.
Шилов И.А. Экология. М.: Высшая школа, 1997. 512 с.
Васильев А.Г., Васильева И.А., Городилова Ю.В., Добринский Н.Л. Принцип компенсации Ю.И. Чернова и влияние полноты состава сообщества грызунов на изменчивость популяции рыжей полевки (Clethrionomys glareolus) на Среднем Урале // Экология. 2017. № 2. С. 116–125. [Vasil’ev A.G., Vasil’eva I.A., Gorodilova Yu.V., Dobrinskii N.L. Chernov’s compensation principle and the effect of rodent community completeness on the variability of Bank Vole (Clethrionomys glareolus) population in the Middle Urals // Rus. J. of Ecology. 2017. V. 48. №. 2. P. 161–169]
Vasil’ev A.G., Vasil’eva I.A., Kourova T.P. Analysis of coupled geographic variation of three shrew species from southern and northern Ural taxocenes // Rus. J. Ecology. 2015. V. 46. № 6. P. 552–558.
Bolnick D.I., Fitzpatrick B.M. Sympatric speciation: models and empirical evidence // Annual Review of Ecology, Evolution, and Systematics. 2007. V. 38. P. 459–487.
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. Lake Tana large barbs: phenetics, growth and diversification // J. Fish Biology. 1996. V. 48. P. 383–404.
Albertson R.C., Kocher T.D. Genetic and developmental basis of cichlid trophic diversity // Heredity. 2006. V. 97. P. 211–221.
Post D.M., Palkovacs E.P. Eco-evolutionary feedbacks in community and ecosystem ecology: interactions between the ecological theatre and the evolutionary play // Philos. Trans. Royal. Soc. Lond. B. Biol. Sci. 2009. V. 364. P. 1629–1640.
Facon B., Genton B.J., Shykoff J. et al. A general eco-evolutionary framework for understanding bioinvasions // Trends in Ecology and Evolution. 2008. V. 21. № 3. P. 130–135.
Vasil’ev A.G., Bol’shakov V.N., Vasil’eva I.A., Sineva N.V. Aftereffects of muskrat introduction in Western Siberia: Morphological and functional aspects // Rus. J. Biological Invasions. 2017. V. 8. № 1. P. 1–9.
Васильев А.Г., Васильева И.А., Городилова Ю.В., Чибиряк М.В. Сопряженная техногенная морфологическая изменчивость двух симпатрических видов грызунов в зоне влияния Восточно-Уральского радиоактивного следа // Вопросы радиационной безопасности. Спецвыпуск. 2013. С. 4–13.
Чернов Ю.И. Видовое разнообразие и компенсационные явления в сообществах и биотических системах // Зоол. журн. 2005. Т. 84. Вып. 10. С. 1221–1238.
Webb C.O., Ackerly D.D., McPeek M.A., Donoghue M.J. Phylogenies and community ecology // Annu. Rev. Ecol. Syst. 2002. V. 33. P. 475–505.
Marzluff J.M. Urban evolutionary ecology // Stud. Avian Biol. 2012. V. 45. P. 287–308.
Васильев А.Г., Васильева И.А., Городилова Ю.В., Чибиряк М.В. Соотношение морфологического и таксономического разнообразия сообществ грызунов в зоне влияния Восточно-Уральского радиоактивного следа на Южном Урале // Экология. 2010. № 2. С. 119–125. [Vasil’ev A.G., Vasil’eva I.A., Gorodilova Y.V., Chibiryak M.V. Relationship between morphological disparity and taxonomic diversity in rodent communities in the zone of influence from the Eastern Ural radioactive trace in the Southern Urals // Rus. J. of Ecology. 2010. V. 41. № 2. С. 153–158]
Букварева Е.Н., Алещенко Г.М. Принцип оптимального разнообразия биосистем. М.: Тов-во научн. изд. КМК, 2013. 522 с.
Глотов Н.В. Генетическая гетерогенность природных популяций по количественным признакам: Автореф. дис. … докт. биол. наук. Ленинград: Ленингр. гос. ун-т, 1983. 33 с.
Zelditch M.L., Swiderski D.L., Sheets H.D., Fink W.L. Geometric morphometrics for biologists: a primer. New York: Elsevier Academic Press, 2004. 437 p.
Васильев А.Г., Большаков В.Н., Васильева И.А. и др. Морфогенетические эффекты переселения представителей южной популяции обыкновенной слепушонки (Ellobius talpinus Pall., 1770) на северную границу ареала // Докл. РАН. 2018. Т. 478. № 5. С. 604–607. [Vasil’ev A.G., Bol’shakov V.N., Vasil’eva I.A. et al. Morphogenetic effects of resettlement of Mole Voles (Ellobius talpinus Pall., 1770) from the southern population to the northern boundary of the species range // Doklady Biol. Sci. 2018. V. 478. P. 26–28]
Васильев А.Г., Большаков В.Н., Васильева И.А. и др. Оценка эффектов неизбирательной элиминации в сообществе грызунов методами геометрической морфометрии // Экология. 2016. № 4. С. 290–299. [Vasil’ev A.G., Bol’shakov V.N., Vasil’eva I.A. et al. Assessment of nonselective elimination effects in rodent communities by methods of geometric morphometrics // Rus. J. Ecology. 2016. V. 47. № 4. P. 383–391]
Шварц С.С. Экспериментальные методы исследования начальных стадий микроэволюционного процесса (постановка проблемы) // Внутривидовая изменчивость наземных позвоночных животных и микроэволюция. Свердловск, 1965. С. 21–32.
Покровский А.В., Большаков В.Н. Экспериментальная экология полевок. М.: Наука, 1979. 147 с.
Васильев А.Г., Васильева И.А., Большаков В.Н. Эволюционно-экологический анализ закономерностей феногенетической изменчивости гомологичных морфоструктур: от популяций до экологических рядов видов // Экология. 2010. № 5. С. 323–329. [Vasilyev A.G., Vasilyeva I.A., Bol’shakov V.N. Evolutionary-ecological analysis of trends in phenogenetic variation of homologous morphological structures: from populations to ecological series of species // Rus. J. Ecology. 2010. V. 41. № 5. P. 365–371]
Васильев А.Г., Васильева И.А. Гомологическая изменчивость морфологических структур и эпигенетическая дивергенция таксонов: основы популяционной мерономии. М.: Тов-во научн. изд. КМК, 2009. 511 с.
Васильев А.Г. Эпигенетические основы фенетики: на пути к популяционной мерономии. Екатеринбург: Академкнига, 2005. 639 с.
Васильев А.Г., Васильева И.А., Шкурихин А.О. Геометрическая морфометрия: от теории к практике. М.: Тов-во научн. изд. КМК, 2018. 471 с.
Haloin J.R., Strauss Sh.Y. Interplay between ecological communities and evolution review of feedbacks from microevolutionary to macroevolutionary scales // Annals of the New York Academy of Sciences. 2008. V. 1133. P. 87–125.
Иорданский Н.Н. Макроэволюция. Системная теория. М.: Наука, 1994. 112 с.
Васильев А.Г., Васильева И.А. Эпигенетические перестройки популяций как вероятный механизм наступления биоценотического кризиса // Вестн. Нижегород. гос. ун-та им. Н.М. Лобачевского. Сер. Биол. 2005. № 1(9). С. 27–38.
Skinner M.K. Environmental epigenetics and unified theory of the molecular aspects of evolution: a neo-Lamarckian concept that facilitates neo-Darwinian evolution // Genome Biol. Evol. 2015. V. 7. P. 1296–1302.
Дополнительные материалы отсутствуют.