

Экология, 2019, № 3, стр. 182-188
Сезонное формирование годичных колец лиственницы сибирской и сосны обыкновенной в зоне южной тайги Средней Сибири
Е. В. Калинина a, А. А. Кнорре a, b, М. В. Фонти a, *, Е. А. Ваганов a, c
a ФГАОУ ВО “Сибирский федеральный университет”
660041 Красноярск, просп. Свободный, 79, Россия
b Государственный природный заповедник “Столбы”
660006 Красноярск, ул. Карьерная, 26а, Россия
c Институт леса им. В.Н. Сукачева СО РАН
660036 Красноярск, Академгородок, 50, стр. 28, Россия
* E-mail: marina.fonti@mail.ru
Поступила в редакцию 14.06.2018
После доработки 30.06.2018
Принята к публикации 09.07.2018
Аннотация
Приведен сравнительный анализ сезонного формирования годичных колец лиственницы сибирской (Larix sibirica Ledeb.) и сосны обыкновенной (Pinus sylvestris L.), произрастающих в зоне южной тайги Средней Сибири на двух участках высотного трансекта (200 и 536 м над ур. м.). Отбор образцов (кернов) проводился в течение 2012 г. с апреля по сентябрь. Результаты наблюдений показали, что начало и продолжительность различных фаз формирования годичных колец лиственницы и сосны обусловлены географическим положением участков и связанным с этим градиентом температур. Следует также отметить влияние видовых особенностей роста деревьев на формирование клеток ксилемы в течение вегетационного периода и, как следствие, на размеры анатомических параметров трахеид.
Прогнозы изменения температуры воздуха, количества атмосферных осадков и увеличения атмосферного СО2 являются основой для ожидаемых изменений в составе, структуре и продуктивности лесных экосистем [1]. Исследования структуры годичных колец, скорости и продолжительности различных фаз формирования ксилемы позволяют установить связь между изменениями окружающей среды и радиальным ростом деревьев, так как фиксируют условия разных промежутков сезона роста и по-разному регистрируют изменения в окружающей среде [2, 3]. В свою очередь структура ксилемы отражает функциональное равновесие между эффективностью водного транспорта (для достижения оптимального роста), затратами для построения клеточных стенок трахеид (для оптимизации и сохранности водопроводящей системы), механической поддержкой ассимиляционного аппарата, а также содержанием воды и неструктурных углеводов (для устойчивости к стрессовым факторам) [4, 5].
Влияние факторов среды на сезонный рост хвойных и лиственных видов древесных растений исследовали с применением различных методов в разных климатических и природных зонах [6–8]. Наибольший интерес представляют работы, рассматривающие продолжительность сезона роста деревьев бореальной зоны на основе исследования формирования древесины [3, 6, 9]. В данной работе мы рассматриваем сезонное формирование годичных колец лиственницы сибирской (хвойного листопадного вида) и сосны обыкновенной (хвойного вечнозеленого вида), произрастающих в низкогорном и среднегорном поясах южной тайги, что обусловливает различный температурный режим территории. Проведение исследований на участках вдоль высотных трансектов [10, 11] имеет особое экологическое значение для интерпретации отклика деревьев на прошлые и прогнозируемые изменения климата, так как представляет собой “природный эксперимент”, при котором начало и продолжительность роста деревьев меняются в течение сезона в зависимости от положения (высоты над ур. м.) исследуемых древостоев и обусловленных этим температур воздуха (до 0.5°С каждые 100 м высоты). Для условий, где фиксируется значительное потепление климата весеннего периода в последние десятилетия и температура воздуха становится определяющим фактором при установлении сроков начала и продолжительности вегетационного периода [12], такой дизайн исследований позволяет оценить изменения прироста основных лесообразующих пород деревьев, которые могут быть вызваны потеплением климата.
Основная цель настоящей работы – установить влияние температуры воздуха на сроки инициации камбиальной активности древесных растений и продолжительность процессов растяжения и утолщения вторичных стенок трахеид, что в свою очередь оказывает влияние на ширину годичных колец и их анатомическую структуру. В задачи работы входило оценить различия в скорости формирования годичных колец лиственницы и сосны в рассматриваемых местообитаниях и выявить основные факторы, определяющие сезонную динамику формирования колец.
МАТЕРИАЛ И МЕТОДЫ
Исследования лиственницы сибирской (Larix sibirica Ledeb.) и сосны обыкновенной (Pinus sylvestris L.) проводили на базе государственного природного заповедника “Столбы” (55°53′ с.ш., 92°46′ в.д., 200 и 536 м над ур. м.). Климат района – резко-континентальный, среднегодовая температура –0.13°С, количество осадков 660 мм/год (данные метеостанции “Столбы” за период 1947–2016 гг.). Год исследования (2012 г.) отличается от среднего за весь период наблюдений более высокими температурами воздуха (13.9 и 12.0°С соответственно) и меньшим количеством осадков (191.4 и 369.4 мм соответственно) в течение сезона роста – с мая по сентябрь.
В 2012 г. были заложены временные пробные площади для изучения внутрисезонных и погодичных изменений процессов формирования ксилемы хвойных пород и оценки связи данных процессов с сезонными фазами вегетации растений. Участки отбирали по принципу приуроченности к постоянным фенологическим площадкам заповедника и двум локальным метеостанциям, расположенных на кордонах Лалетино и Нарым. Участок Лалетино (LAL, 200 м над ур. м.): LAL-1 – сосново-березовый разнотравно-злаково-осочковый лес; LAL-2 – светлохвойный злаково-осочковый лес. Участок Нарым (NAR, 536 м над ур. м.) – светлохвойно-березовый крупнотравно-злаково-осочковый лес. Согласно данным, полученным с локальных метеостанций, в 2012 г. на участках Лалетино и Нарым сумма осадков за вегетационный период составила 81.4 и 191.4 мм, среднегодовая температура воздуха 0.06 и –0.37°С, температура поверхности почвы 4.4 и 3.2°С соответственно. Температура почвы на глубине 20 см (измерения проводили только для участка Нарым) составила 4.0°С.
На участке NAR были выбраны 4 доминантных дерева каждого вида (сосна и лиственница), на участке LAL-1 – 4 дерева сосны, а на LAL-2 – 4 лиственницы. Возраст деревьев для NAR и LAL был одинаков и составил 90 лет у сосны и 105 лет у лиственницы. Высота и диаметр деревьев 25 м и 30 см для лиственницы, 22 м и 43 см для сосны соответственно. С середины апреля по середину сентября 2012 г. с каждого дерева были взяты керны древесины длиной около 1 см (включающие 2–3 годичных кольца последних лет роста) возрастным шведским буравом Hagloff спирально по окружности ствола с шагом 5 см на высоте 1.3 м от поверхности почвы с периодичностью один раз в неделю. Непосредственно сразу после сбора образцы помещали в пробирки с фиксирующей жидкостью глицерин-спирт-вода в соотношении 1 : 1 : 1. Всего в течение вегетационного периода сбор сезонных образцов древесины проводился 22 раза.
Параметры формирующихся годичных колец измеряли на тонких поперечных срезах древесины, полученных с помощью санного микротома МС-2. Срезы помещали в раствор астра-голубого (2%-ный раствор) и сафранина (1%-ный раствор) для контрастного окрашивания лигнифицированных и нелигнифицированных тканей. Размеры различных зон формирующихся колец, число и размеры клеток измеряли при помощи системы анализа изображений (Carl Zeiss, Jena, Germany) и программного пакета AxioVision SE64 Rel. 4.9.1, позволяющих определять параметры клеточных структур в разных частях годичного кольца в полуавтоматическом режиме. В каждом годичном кольце измеряли 3 ряда клеток от внутренней границы к внешней, затем значения усредняли.
Нами выбрана следующая схема разделения клеток на зоны: 1) камбиальная; 2) зона растяжения клеток; 3) зона первичной лигнификации клеточной стенки; 4) зона зрелых трахеид. Подробное деление формирующегося кольца с выделением отдельных зон растяжения и первичной лигнификации было возможно благодаря применению поляризационного фильтра.
Для последнего сезонного образца измерены параметры трахеид полностью сформированного годичного кольца 2012 г. Выбраны 5 рядов клеток с наибольшей площадью поперечного сечения. Измеряли радиальный размер люмена (LD) и толщину двойной клеточной стенки (2CWT), по которым затем рассчитывали радиальный размер трахеид (D = = LD + 2CWT). Cогласно индексу Морка [13], на основе соотношения ширины двойной клеточной стенки и диаметра люмена в каждом кольце были установлены размеры зон ранней (EW, 2CWT < LD) и поздней (LW, 2CWT > LD) древесины.
РЕЗУЛЬТАТЫ
Камбиальная зона исследуемых деревьев в состоянии покоя (весной и осенью) включала от 4 до 8 клеток (рис. 1б, г, д, е). Начало камбиальной активности в сторону ксилемы на участке LAL (200 м над ур. м.) и у лиственницы, и у сосны было синхронным и наблюдалось с 12 мая. Сумма температур воздуха за 10 дней, предшествующих делению камбиальных клеток, составила 79.34°C, а сумма температур поверхности почвы 38.17°C (данные с локальной метеостанции LAL). На участке NAR (536 м над ур. м.) камбиальная активность лиственницы началась 18 мая, а у сосны на неделю позже при сумме средних температур воздуха и поверхности почвы 64.64 и 46.89°C соответственно (для периода с 15 по 24 мая, данные с локальной метеостанции NAR).
Рис. 1.
Суточные изменения температуры воздуха (T), температуры поверхности почвы (tsoil) и количества осадков (P) с локальных метеостанций на участках Лалетино (LAL (а)) и Нарым (NAR (б), пробел в метеоданных с 23.04 по 14.05). Кругами обозначены фазы распускания хвои лиственницы: светло-серый – распускание почек, серый – начало развертывания хвои, черный – полное развертывание. Усредненное (количество деревьев/вид/участок) количество клеток (± стандартная ошибка) в различных зонах годичного кольца лиственницы сибирской (в, г) и сосны обыкновенной (д, е) в течение сезона роста: 1 – камбиальная зона, 2 – зона растяжения клеток, 3 – зона первичной лигнификации клеточной стенки, 4 – зона зрелых трахеид, 2 + 3 + 4 – общее количество клеток ксилемы.
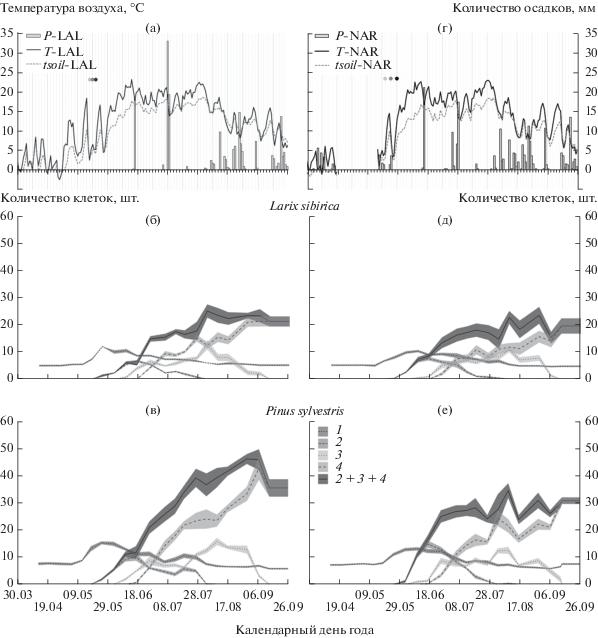
Камбиальная активность наблюдалась в течение 63 дней для двух видов с участка NAR и 70 и 77 дней – для лиственницы и сосны соответственно с участка LAL (табл. 1). В этот период среднее количество клеток в зоне для деревьев лиственницы составило 9 и 11 – для сосны на каждом из участков. Максимальная активность камбиальной зоны деревьев на участке NAR наблюдалась на неделю раньше (25 мая) по сравнению с участком LAL (2 июня), в это же время появились первые клетки ксилемы в зоне растяжения (см. рис. 1б, г, д, е). Окончание камбиальной активности было зафиксировано в конце июля и совпало с периодом максимальных среднесуточных температур воздуха (до 22.16°С, рис. 1а, б). Процесс растяжения закончился у лиственницы на участке Лалетино 27 июля, а у остальных деревьев на двух участках одновременно 3 августа.
Таблица 1.
Продолжительность различных фаз формирования годичных колец лиственницы сибирской и сосны обыкновенной в 2012 г.
| Учас-ток | Вид | 13.04 | 20.04 | 27.04 | 05.05 | 12.05 | 18.05 | 25.05 | 02.06 | 10.06 | 17.06 | 26.06 | 06.07 | 13.07 | 19.07 | 27.07 | 03.08 | 10.08 | 17.08 | 30.08 | 07.09 | 14.09 | 27.09 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LAL | LASI | Кн | Рн | Лн | Зн | Ко | Ро | Ло | Зо | ||||||||||||||
| PISY | Кн | Рн | Лн | Зн | Ко | Ро | Ло | Зо | |||||||||||||||
| NAR | LASI | Кн | Рн | Лн | Зн | Ко | Ро | Ло | Зо | ||||||||||||||
| PISY | Кн | Рн | Лн | Зн | Ко | Ро | Ло | Зо |
Примечание. LAL – Лалетино; NAR – Нарым; LASI – лиственница сибирская, PISY – сосна обыкновенная; Кн и Ко – начало и окончание камбиальной активности; Рн и Ро – начало и окончание фазы растяжения трахеид; Лн и Ло – начало и окончание фазы лигнификации клеточной стенки; Зн и Зо – начало и окончание появления зрелых трахеид в ксилеме.
Появление клеток в зоне первичной лигнификации и утолщение клеточных стенок зафиксированы 10 июня для всех исследуемых видов (см. табл. 1). Последние клетки в этой зоне на участке LAL у лиственницы наблюдались на неделю раньше, чем у сосны (30 августа и 7 сентября соответственно), на участке NAR – 7 сентября у лиственницы и 14 сентября у сосны.
Появление зрелых трахеид с полностью сформированной вторичной клеточной стенкой в ранней древесине сосны наблюдалось 17 июня на участке LAL и было синхронным (26 июня) у деревьев разных видов на участке NAR и лиственницы с участка LAL (см. табл. 1). Завершилось формирование годичного кольца на участке LAL у лиственницы 7 сентября, у сосны – на неделю позже, на участке NAR – 14 сентября у лиственницы и 27 сентября у сосны. Несмотря на положение исследуемых участков вдоль высотного трансекта и вызванные этим различия температурного режима, продолжительность сезона роста (от активации камбия до завершения лигнификации клеточных стенок трахеид поздней древесины) для низкогорного (LAL) и среднегорного (NAR) участков была одинакова и составила 118 дней для деревьев лиственницы и 125 дней – для деревьев сосны.
В течение сезона роста 2012 г. относительно широкие кольца (более 1 мм) были сформированы деревьями сосны на двух исследуемых участках (табл. 2). В связи с высокой корреляцией между шириной кольца и количеством клеток, его образующих, максимальное количество клеток (56) также соответствует данному виду. В среднем количество сформированных трахеид в годичном кольце сосны для четырех деревьев составило 33. При этом параметры трахеид (толщина клеточной стенки и радиальный размер клетки) отличались у деревьев с разных участков не только по средним значениям для кольца (табл. 2, рис. 2), но и отдельно для зоны ранней (Dew = 35.6 и 37.0 мкм, CWTew = 3.3 и 3.0 мкм) и поздней (Dlw = 20.2 и 17.3 мкм, CWTlw = 5.6 и 4.8 мкм) древесины для участков LAL и NAR соответственно, что в свою очередь отразилось на ширине каждой зоны.
Таблица 2.
Среднеарифметические показатели параметров трахеид лиственницы сибирской и сосны обыкновенной, сформированных в 2012 г.
| Участок | Вид | TRW, мкм | Число клеток в ряду | LD | D | CWT | EW/LW, % |
|---|---|---|---|---|---|---|---|
| мкм | |||||||
| LAL | LASI | 819 | 24 | 22 | 34 | 6.1 | 45/55 |
| PISY | 1022 | 33 | 23 | 31 | 4.0 | 68/32 | |
| NAR | LASI | 571 | 17 | 23 | 34 | 5.5 | 56/44 |
| PISY | 1026 | 34 | 24 | 31 | 3.5 | 72/28 | |
Рис. 2.
Диаграмма размаха (boxplot) изменений радиального размера трахеид (1) и толщины их клеточной стенки (2) внутри годичных колец лиственницы и сосны, сформированных в 2012 г., с шагом 5% относительно расстояния внутри годичного кольца: а – участок Лалетино, б – участок Нарым. Линией внутри “ящика” показана медиана, размер “ящика” определен нижним и верхним квартилями (25 и 75% выборки соответственно), “усы” – минимальное и максимальное значение выборки, точки за пределами – выбросы.
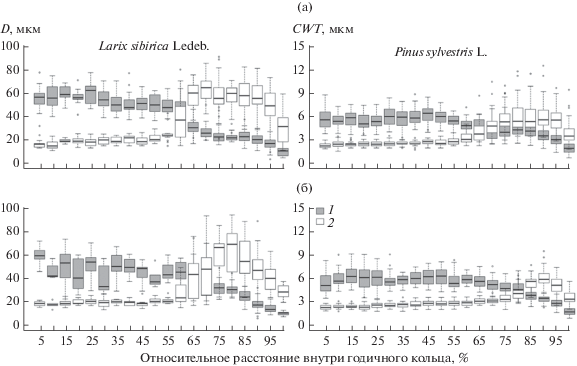
Минимальная ширина годичных колец отмечена для деревьев лиственницы на участке Нарым. Она была на 20% меньше, чем на участке Лалетино, что связано с размерами трахеид ранней (Dew = 52.5 и 47.3 мкм, CWTew = 3.4 и 3.5 мкм) и поздней (Dlw = 20.7 и 19.7 мкм, CWTlw = 8.1 и 7.6 мкм) древесины для LAL и NAR соответственно. При этом процент ранней древесины был выше для лиственницы с участка Нарым (56%), чем для деревьев с участка Лалетино (45%).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Ранее для периода с 1947 по 2011 г. было установлено [14, 15], что рост сосны обыкновенной в районе исследования определяется весенними температурами (R = 0.30–0.42, p < 0.05), тогда как для лиственницы сибирской значимую роль играет температура июня (R = 0.33, p < 0.05). Такие различия авторы связывают с тем, что физиологически активные процессы (водопроведение, фотосинтез и др.) у сосны и лиственницы могут начинаться в разное время за счет необходимости ежегодного формирования нового фотосинтезирующего аппарата у последней. Однако начало именно камбиальной активности, по полученным нами данным, наступает практически одновременно у двух видов (участок Лалетино), либо фиксируется запаздывание прироста сосны на одну неделю по сравнению с лиственницей (участок Нарым). Это соответствует результатам, указывающим на более ранние сроки наступления фенологических этапов развития хвои у лиственницы по сравнению с сосной [12]. Данное явление может быть обусловлено не только наличием или отсутствием фотосинтетического аппарата у деревьев, но и перераспределением резервных ассимилятов внутри растения при инициации тех или иных процессов [16]. Количество осадков оказывает значимое влияние на прирост сосны в июне (R = 0.30, p < 0.05), тогда как для лиственницы значимых корреляций между шириной годичного кольца и осадками не обнаружено.
Влияние температуры почвы на начало камбиальной активности и прирост древесных растений имеет сложный характер [17–19], несмотря на то, что при росте температуры воздуха соответственно увеличивается и температура почвы. Нижний порог этого параметра широко известен в пределах 3–5°С [20], хотя для трех хвойных видов Канады, основываясь на прямых измерениях, вероятность наступления камбиальной активности была рассчитана при минимальной, средней и максимальной температуре почвы на глубине 15 см равной 0.2–4.5, 2.6–7.5 и 8.3–12.6°С соответственно [18]. Полученные нами данные для участков LAL и NAR, где среднесуточная температура поверхности почвы за десятидневный период до начала камбиальной активности составила 3.8 и 4.7°С соответственно, а температура почвы на глубине 20 см на участке Нарым была равна 0.6°С, схожи с ранее опубликованными результатами исследований [18]. Данные о начале сезона роста сосны и лиственницы при средней за 10‑дневный период температуре воздуха 7.9 и 6.5°С на участках Лалетино и Нарым также соответствуют ранее опубликованным результатам для деревьев бореальной зоны [18, 21], когда начало камбиальной активности происходит при достижении среднесуточных температур 6–8°С.
Ранее для лиственницы европейской вдоль высотного трансекта в Альпах было установлено, что запаздывание начала камбиальной активности составило 3–4 дня на каждые 100 м над ур. м. [11]. Схожие результаты получены нами для деревьев сосны, когда начало сезона роста было зафиксировано для высокогорного участка на 13 дней позже, чем для низкогорного. Для лиственницы запаздывание составило только 6 дней.
Согласно ряду исследований [22–24], влияние температуры воздуха на реактивацию камбиальных тканей происходит опосредованно через активацию и перенос ауксина внутри стебля, при этом низкие весенние температуры могут замедлять как его биосинтез, так и транспорт. Источниками ауксина являются апикальные меристемы, в том числе почки, развивающаяся хвоя и побеги [25, 26], в связи с чем согласованность фенологии хвои и начала ксилогенеза представляет собой особый интерес [27, 28]. Несмотря на очевидную связь этих двух процессов, ранее полученные результаты противоречат друг другу. Исследование лиственницы европейской на высотном трансекте в Швейцарских Альпах [11] показало, что радиальный рост деревьев начинается на 3–4 недели после распускания хвои, тогда как для хвойных Италии и Канады обнаружена другая последовательность [27, 29]. В нашем исследовании на участке Лалетино деление камбиальных клеток лиственницы было зафиксировано на неделю раньше начала распускания хвои [30], тогда как для лиственницы с участка Нарым эти две фазы совпали (см. рис. 1б, г). В целом периоды между фазами распускания хвои лиственницы (распускание почек, начало развертывания и полное развертывание хвои) для участка LAL были короче (2 дня), чем для участка NAR (4 дня).
Следует отметить, что ранее опубликованные данные о лимитировании роста деревьев температурой воздуха в конце вегетационного периода, когда она опускается до среднесуточных значений 8–9°С [27], соответствуют только для сосны с участка Нарым. Непосредственно перед окончанием сезона роста лиственницы с двух участков, а также сосны с участка Лалетино (когда все клетки полностью закончили лигнификацию клеточных стенок) средняя температура все еще достигала 12–13°С.
В связи с тем, что основным лимитирующим рост деревьев фактором в зоне южной тайги является температура воздуха, достаточно сложно установить влияние количества осадков на начало и продолжительность камбиальной активности, а также на процессы формирования годичного кольца деревьев. Чтобы получить достоверные результаты, необходимо продолжить мониторинг сезонного роста годичных колец и использовать наблюдения для нескольких лет (или сезонов роста).
Несмотря на широко известную видовую особенность изменчивости радиального размера люмена и клеточной стенки трахеид, при которой сосна характеризуется меньшим размером клеток ранней древесины, но более широкой переходной зоной, тогда как для лиственницы как таковая переходная между ранней и поздней древесиной зона почти отсутствует, о внутрисезонной динамике анатомических параметров ксилемы для одного и того же вида вдоль высотного трансекта известно мало. Так, основные различия как сосны, так и лиственницы для разных участков наблюдались в соотношении зон ранней и поздней древесины, обусловленные изменением толщины клеточной стенки при относительно схожих размерах люмена (см. табл. 2, рис. 2). При этом более широкая клеточная стенка трахеид и в целом ширина поздней древесины были характерны для деревьев на участке LAL. На основании ранее полученных результатов о влиянии климатических условий на радиальный рост исследуемых видов [14, 15] и том факте, что в 2012 г. период с мая по август был на 3°С теплее и на 42% суше по сравнению со средним для 1947–2016 гг., можно предположить, что при оптимальных условиях радиальный прирост деревьев и анатомическая структура годичных колец сосны и лиственницы будет значительно отличаться от сформированных в 2012 г.
Таким образом, полученные нами данные подтверждают влияние температуры воздуха и почвы на начало ксилогенеза сосны обыкновенной и лиственницы сибирской, при этом для среднегорного участка деление камбиальных клеток начинается при относительно низких температурах по сравнению с низкогорным. Завершение камбиальной активности совпадает с периодом максимальных температур воздуха. Полученные связи и климатические данные в начале и конце сезона роста могут быть использованы в дальнейшем как входящие параметры имитационной модели роста древесных растений Ваганова-Шашкина [2] и позволят получить биологически обусловленную согласованность между исходными древесно-кольцевыми хронологиями и имитационными кривыми. Несмотря на различия в продолжительности камбиальной активности до двух недель, в целом длина сезона роста для двух участков была схожей и составила 118 дней для лиственницы и 125 дней для сосны. Данные анатомических параметров трахеид исследуемых видов подтверждают как видовую специфичность, так и влияние условий произрастания на рост сосны обыкновенной и лиственницы сибирской вдоль высотного трансекта.
Работа выполнена при поддержке РФФИ (проект № 17-04-00610-а).
Список литературы
IPCC V. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Eds. Stocker T.F., Qin D., Plattner G.-K., Tignor M., Allen S.K., Boschung J., Nauels A., Xia Y., Bex V., Midgley P.M. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 2013. 1535 p.
Ваганов Е.А., Шашкин А.В. Рост и структура годичных колец хвойных. Новосибирск: Наука, 2000. 232 с.
Rossi S., Anfodillo T., Čufar K. et al. Pattern of xylem phenology in conifers of cold ecosystems at the northern hemisphere // Global Change Biology. 2016. V. 22. P. 3804–3813. https://doi.org/10.1111/gcb.13317
Свидерская И.В., Суховольский В.Г., Радостева Е.Ю., Кирдянов А.В. Модельная оценка оптимального соотношения между толщиной клеточной стенки и размером люмена у трахеид хвойных // Журн. Сибирского федерального ун-та. Биология. 2011. Т. 2. № 4. С. 183–196.
Fonti P., Jansen S. Xylem plasticity in response to climate // New Phytol. 2012. V. 195(4). P. 734–736. doi.org/ https://doi.org/10.1111/j.1469-8137.2012.04252.x
Antonova G.F., Stasova V.V. Effects of environmental factors on wood formation in Scots pine stems // Trees. 1993. V. 7. P. 214–219. https://doi.org/10.1007/ BF00202076
Deslauriers A., Morin H. Intra-annual tracheid production in balsam fir stems and the effect of meteorological variables // Trees. 2005. V. 19(4). P. 402–408. https://doi.org/10.1007/s00468-004-0398-8
Eilmann B., Zweifel R., Buchmann N. et al. Drought-induced adaptation of the xylem in Scots pine and pubescent oak // Tree Physiol. 2009. V. 29. P. 1011–1020. https://doi.org/10.1093/treephys/tpp035
Брюханова М.В., Кирдянов А.В., Прокушкин А.С., Силкин П.П. Особенности ксилогенеза Larix gmelinii (Rupr.) Rupr. в условиях криолитозоны Средней Сибири // Экология. 2013. № 5. С. 323–329. [Bryukhanova M.V., Kirdyanov A.V., Prokushkin A.S., Silkin P.P. Specific features of xylogenesis in Dahurian larch, Larix gmelinii (Rupr.) Rupr., growing on permafrost soils in Middle Siberia // Rus. J. of Ecology. 2013. V. 44. № 5. P. 361–366. doi 10.1134/S1067413613050044]
Körner C. The use of ‘altitude’ in ecological research // Trends Ecol. Evol. 2007. V. 22. P. 569–574. https://doi.org/10.1016/j.tree.2007.09.006
Moser L., Fonti P., Büntgen U. et al. Timing and duration of European larch growing season along altitudinal gradients in the Swiss Alps // Tree Physiol. 2010. V. 30. P. 225–233. https://doi.org/10.1093/treephys/tpp108
Овчинникова Т.М., Фомина В.А., Андреева Е.Б. и др. Анализ изменений сроков сезонных явлений у древесных растений заповедника “Столбы” в связи с климатическими факторами // Хвойные бореальной зоны. 2011. №1–2. С. 47–54.
Denne M.P. Definition of latewood according to Mork (1928) // IAWA Bulletin. 1989. V. 10. P. 59–62. https://doi.org/10.1163/22941932-90001112
Кнорре А.А., Конурбаева Р.У. Влияние рекреации на формирование радиального прироста сосны обыкновенной (Pinus sylvestris L.) в заповеднике “Столбы” // Научные исследования в заповедниках и национальных парках Южной Сибири. Новосибирск: Изд-во СО РАН, 2012. Вып. 2. С. 160–165.
Конурбаева Р.У. Реакция основных древесных видов горно-таежного пояса Восточного Саяна на изменение климата: Дис. на соиск. степени магистра. Красноярск: Сибирский федер. ун-т, 2012. 81 с.
Chapin F.S., Schulze E., Mooney H.A. The ecology and economics of storage in plants // Ann. Rev. Ecol. Syst. 1990. V. 21. P. 423–447. https://doi.org/10.1146/annurev.es.21.110190.002231
Landhausser S.M., DesRochers A., Lieffers V.J. A comparison of growth and physiology in Picea glauca and Populus tremuloides at different soil temperatures // Can. J. For. Res. 2001. V. 31(11). P. 1922–1929. https://doi.org/10.1139/x01-129
Rossi S., Deslauriers A., Anfodillo T., Carraro V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes // Oecologia. 2007. V. 152. P. 1–12. https://doi.org/10.1007/s00442-006-0625-7
Lupi C., Morin H., Deslauriers A., Rossi S. Xylogenesis in black spruce: does soil temperature matter? // Tree Physiol. 2012. V. 32. P. 74–82. https://doi.org/10.1093/treephys/tpr132
Körner C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Berlin, Heidelberg: Springer-Verlag, 2003. 349 p. https://doi.org/10.1007/978-3-642-18970-8
Rossi S., Deslauriers A., Gričar J. et al. Critical temperatures for xylogenesis in conifers of cold climates // Global Ecol. Biogeogr. 2008. V. 17. P. 696–707. https://doi.org/10.1111/j.1466-8238.2008.00417.x
Oribe Y., Kubo T. Effect of heat on cambial reactivation during winter dormancy in evergreen and deciduous conifers // Tree Physiol. 1997. V. 17. P. 81–87. https://doi.org/10.1093/treephys/17.2.81
Uggla C., Magel E., Moritz T., Sundberg B. Function and dynamics of auxin and carbohydrates during earlywood/latewood transition in Scots pine // Plant Physiol. 2001. V. 125(4). P. 2029–2039. https://doi.org/10.1104/pp.125.4.2029
Oribe Y., Funada R., Kubo T. Relationships between cambial activity, cell differentiation and the localization of starch in storage tissues around the cambium in locally heated stems of Abies sachalinensis (Schmidt) Masters // Trees: Struct. Funct. 2003. V. 17. P. 185–192. https://doi.org/10.1007/s00468-002-0231-1
Savidge R.A. Auxin and ethylene regulation of diameter growth in trees // Tree Physiol. 1988. V. 4(4). P. 401–414. https://doi.org/10.1093/treephys/4.4.401
Sundberg B., Uggla C. Origin and dynamics of indoleacetic acid under polar transport in Pinus sylvestris // Physiol. Plant. 1998. V. 104(1). P. 22–29. https://doi.org/10.1034/j.1399-3054.1998.1040104.x
Rossi S., Rathgeber C.B.K., Deslauriers A. Comparing needle and shoot phenology with xylem development on three conifer species in Italy // Ann. For. Sci. 2009. V. 66(2). P. 206. https://doi.org/10.1051/forest/2008088
Swidrak I., Schuster R., Oberhuber W. Comparing growth phenology of co-occurring deciduous and evergreen conifers exposed to drought // Flora. 2013. V. 208. P. 609–617. https://doi.org/10.1016/j.flora.2013.09.004
Zhai L., Bergeron Y., Huang J.G., Berninger F. Variation in intra-annual wood formation, and foliage and shoot development of three major Canadian boreal tree species // Amer. J. Bot. 2012. V. 99(5). P. 827–837. https://doi.org/10.3732/ajb.1100235
Должковая Н.П. Календарь растений // Летопись природы государственного природного заповедника “Столбы”. 2012. Кн. 70. С. 61–65.
Дополнительные материалы отсутствуют.