

Экология, 2019, № 3, стр. 233-235
Фактор солености в микроэволюции рыб и распределение возрастных групп алтайского османа Oreoleuciscus potanini (Cyprinidae) в озерах Айраг и Хяргас (Монголия)
А. Н. Мироновский a, b, *, А. С. Маврин b, А. В. Кожара b, Ю. В. Слынько c
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
b Институт биологии внутренних вод им. И.Д. Папанина РАН
152743 Ярославская обл., Некоузский р-н, п. Борок, 109, Россия
c Институт морских биологических исследований им. А.О. Ковалевского РАН
299011 Севастополь, просп. Нахимова, 2, Россия
* E-mail: adissa@mail.ru
Поступила в редакцию 13.07.2018
После доработки 04.09.2018
Принята к публикации 18.09.2018
Изменения экологических факторов среды обитания влекут за собой генетические и, как следствие, фенетические перестройки популяций [1]. Для гидробионтов важным эволюционным фактором является соленость воды [2]. В частности, показано, что популяции карповых рыб, использующие для нагула солоноватые воды Каспия либо вселенные в минерализованные озера Казахстана, демонстрируют морфологические и генетические отличия от пресноводных популяций [3, 4].
Вместе с тем сравнение алтайских османов O. potanini, обитающих в солоноватом (7.63‰) оз. Хяргас и пресноводном оз. Айраг, достоверных фенетических различий не выявило, а по генетическим показателям выборки Айрага оказались ближе к выборкам Хяргаса, чем к выборкам пресноводного оз. Ногон той же речной системы [5]. Отсутствие выраженной дивергенции между конспецифичными группами, обитающими в озерах разной степени солености, противоречит данным предыдущих работ [3, 4]. Чтобы прояснить этот вопрос, мы провели дополнительное исследование, включающее определение и анализ возрастного состава выборок экологических форм османа (растительноядной и рыбоядной), анализ биотопических особенностей озер Айраг и Хяргас и особенностей ландшафта изучаемого района.
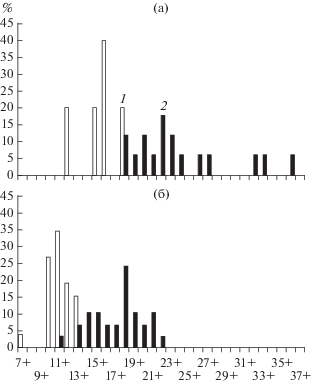
Возраст рыб оценивали по operculum с контролем по cleithrum. Кости рассматривали под бинокулярным микроскопом в проходящем свете, предварительно просветляя в 96%-ном этиловом спирте. Как следует из рис. 1, распределение особей по возрастным группам в озерах Айраг и Хяргас неодинаково. Если в первом предельный возраст рыбоядной особи равен 18+, то возрастной ряд рыбоядных османов оз. Хяргас начинается с 18+. Распределения рыб растительноядной формы в сравниваемых озерах перекрываются, однако и здесь их различие четко выражено.
Рис. 1.
Возрастной состав выборок алтайского османа (а – рыбоядная форма; б – растительноядная форма) из озер Айраг и Хяргас: 1 – возрастные группы оз. Айраг (31 особь), 2 – возрастные группы оз. Хяргас (46 особей).
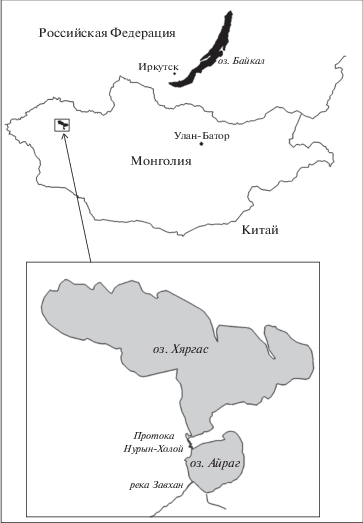
На наш взгляд, из всех возможных интерпретаций полученных результатов наименьших допущений требует гипотеза, исходящая из пресноводного происхождения рода Oreoleuciscus. По данным академика А. Дулмаа [6], “Османы обитают в водоемах …. разной степени осолонения … во время нереста наблюдается миграция из осолоненных озер в пресные реки” (c. 220). В нашем случае пресное оз. Айраг и осолоненное оз. Хяргас завершают водную систему р. Завхан, которая впадает в оз. Айраг, и далее по 5-километровой протоке Нурын-Холой вода поступает в бессточное оз. Хяргас, уровень которого на 2 м ниже уровня оз. Айраг (рис. 2). Ширина протоки – до 100 м, глубина – до 11 м. Другими словами, Нурын-Холой – это полноводная пресная река. Войдя в нее на нерест из минерализованного оз. Хяргас, османы через 5 км оказываются в оз. Айраг. Известно, что алтайские османы откладывают икру на любой субстрат, но при наличии выбора предпочитают грунты с растительностью [6, 7]. Следовательно, в оз. Айраг, входящем в число основных тростниковых озер региона, условия размножения для них лучше, чем в оз. Хяргас, где водорослевые заросли наблюдаются лишь вдоль северного берега, а высшая водная растительность практически отсутствует [6, 8]. Мало пригодна для нереста и протока Нурын-Холой с ее каменистым дном и почти отвесными скальными берегами.
Говоря о худших по сравнению с оз. Айраг условиях размножения в оз. Хяргас, необходимо учитывать и чисто физиологические последствия высокой минерализации последнего. Величина 7.63‰ лежит у верхнего порога критической солености 5–8‰, который считается физиологической границей, отделяющей пресноводную фауну от морской [9]. В частности, эмбрионы пресноводных рыб после выклева способны развиваться в воде с соленостью не более 5–7‰ [10].
Размножение в пресноводной части ареала (обычное явление у рыб, освоивших для нагула соленые воды) в изучаемой ситуации объясняет отсутствие фенотипических и генетических проявлений действия солености. Выклюнувшиеся из икринок на нерестилищах оз. Айраг османы часть жизни проводят в пресной воде, далее мигрируют в оз. Хяргас, а для размножения возвращаются в оз. Айраг. Нерестуют все рыбы, видимо, на одних нерестилищах, что не позволяет закрепиться дивергенции признаков, если таковая возникает в силу разной солености озер. В случае каждой экологической формы (растительноядной и рыбоядной) алтайские османы озерной системы “Айраг-Хяргас” представляют собой репродуктивно и соответственно генетически и фенетически единую популяцию, характеризующуюся пространственной дифференциацией возрастных групп.
Изложенная выше трактовка полученных результатов основана на косвенных данных, подтвердить которые могут лишь прямые наблюдения за миграциями и нерестом рыб. Вместе с тем совокупность косвенных данных представляется достаточно убедительной, чтобы рассматривать описанную схему жизненного цикла османов озерной системы “Айраг–Хяргас” в качестве рабочей гипотезы.
В заключение отметим наглядность изучаемой ситуации как примера того, что наряду с оценкой фенетических и генетических параметров в исследованиях биологического разнообразия методически важно анализировать и структуру сравниваемых популяций, в частности возрастную.
Авторы искренне благодарны М. Эрдэнэбату (Институт геоэкологии Академии наук Монголии), Ю.Ю. Дгебуадзе (ИПЭЭ РАН), А.В. Крылову и Д.П. Карабанову (ИБВВ РАН) за помощь при сборе материала. Работа выполнена при финансовой поддержке РФФИ (проекты № 08-04-00061а и 14-04-00022). А.Н. Мироновским работа выполнена в рамках раздела Государственного задания ИПЭЭ РАН (№ 0109-2018-0076), Ю.В. Слынько – в рамках бюджетного финансирования ИМБИ РАН по теме № АААА-А18-118020890074-2, А.С. Мавриным – в рамках темы Государственного задания ИБВВ РАН № АААА-А18-118012690101-2, А.В. Кожарой – в рамках темы Государственного задания ИБВВ РАН № АААА-А18-118012690222-4.
Список литературы
Шварц С.С. Экологические закономерности эволюции. М.: Наука, 1980. С. 227.
Одум Ю. Основы экологии. М.: Мир, 1975. С. 740.
Мироновский А.Н., Кожара А.В., Яковлев В.Н. Закономерности формирования популяционной структуры карповых рыб Волго-Каспийского района // Экология. 1989. № 1. С. 21–27. [A.N. Mironovskii, A.V. Kozhara, V.N. Yakovlev. Principles of formation of population structure in cyprinid fishes of the Volga-Caspian region // Sov. J. of Ecology. 1989. V. 20. № 1. P. 17–22.]
Слынько Ю.В., Мироновский А.Н. Популяционно-генетические дополнения к вопросу о роли фактора солености в микроэволюции пресноводных рыб // Экология. 1993. № 1. С. 82–86. [Y.V. Slynko, A.N. Miro-novskii. Population-genetic additions to the question of the role of the salinity factor in microevolution of fresh-water fish // Rus. J. of Ecology. 1993. V. 24(1). P. 66–69.]
Мироновский А.Н., Слынько Ю.В., Касьянов А.Н., Горошкова Т.В. Сравнительный анализ изменчивости экологических форм алтайского османа Oreoleuciscus potanini (Cyprinidae) в озерах Айраг, Хяргас и Ногон (Западная Монголия) // Экология. 2018. Вып. 3. С. 214–219. https://doi.org/10.7868/S036705971803006X. [A.N. Mironovsky, Y.V. Slynko, A.N. Kasyanov, T.V. Goroshkova. Comparative Analysis of Variation in Ecological Forms of Altai Osman, Oreoleuciscus potanini (Cyprinidae), in Lakes Ayrag, Khyargas, and Nogon (Western Mongolia) // Rus. J. of Ecology. 2018. V. 49(2). P. 160–165. https://doi.org/10.1134/S1067413618020091.]
Дулмаа А. Биология озер Монголии. Улан-Батор, 2015. С. 330.
Баасанжав Г., Дгебуадзе Ю.Ю., Демин А.Н. и др. Экология и хозяйственное значение рыб МНР. М.: Наука, 1985. С. 200.
Болдбаатар S., Звонов Б.М., Букреев С.А. Современное состояние орнитофауны Монголии и изменение среды ее обитания // Экологические последствия биосферных процессов в экотонной зоне Южной Сибири и Центральной Азии. Улан-Батор, 2010. С. 228–230.
Хлебович В.В. Критическая соленость биологических процессов. Л.: Наука, 1974. С. 236.
Мартемьянов В.И. Влияние солености на пресноводных рыб // Зоол. журн. 1989. Т. 68. Вып. 5. С. 72–81.
Дополнительные материалы отсутствуют.