

Экология, 2019, № 3, стр. 172-181
Гипотеза о лофотенском плейстоценовом рефугиуме Pinus sylvestris L.
С. Н. Санников a, Н. С. Санникова a, *, И. В. Петрова a, Е. В. Егоров a
a Ботанический сад УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202а, Россия
* E-mail: sannikovanelly@mail.ru
Поступила в редакцию 04.05.2018
После доработки 07.06.2018
Принята к публикации 02.07.2018
Аннотация
Проведен анализ дат появления репрезентативной доли пыльцы Pinus sylvesris L. в палиноспектрах (≥20%), отражающей распространение ее авангардных массивов из Центральной и Восточной Европы вдоль вероятных альтернативных путей иммиграции популяций в регион Лофотена. Выявлено, что даже при максимально возможной скорости расселения (300–500 км/1000 лет) они могли бы достичь этого региона не ранее 6500–7000 лет BP, т.е. на 2500–3000 лет позднее фактической палинологической даты. Таким образом, вероятность происхождения лофотенских популяций P. sylvestris от адвентивных, иммигрировавших в Скандинавию из смежных регионов Европы, исключается. Обобщение результатов изучения хронопалинологии и скорости голоценовой реколонизации P. sylvestris, данных по палео-гляциологии, -маринологии и палеоботанике, а также современной геногеографии и экологии сосны обыкновенной подтвердило гипотезу о выживании в регионе Лофотенских островов в течение последней гляциальной фазы и расселении в голоцене автохтонных рефугиальных популяций этого вида.
Одним из дискуссионных вопросов филогеографии популяций Pinus sylvestris L. и других видов является вероятность сохранения в ледниковые фазы плейстоцена их локальных рефугиумов в некоторых перигляциальных местообитаниях [1–9]. Стации долговременного выживания, воспроизведения и непрерывного существования небольших групп деревьев (микропопуляций) сосны обыкновенной и других растений в условиях фонового ледникового ландшафта могут сохраняться в наиболее прогреваемых микроэкотопах, защищенных от холодных ветров горными хребтами и отепляемых крупными водоемами. Таковы, например, омываемые морскими течениями юго-западное побережье Ирландии [2, 10] или южные склоны интенсивно инсолируемых песчаных террас рек юго-восточной Прибалтики [3, 5]. В плейстоцене теплообеспеченность некоторых “экологических ниш” в подобных регионах могла быть намного благоприятнее для выживания и возобновления автохтонных популяций растений, чем в окружающем ландшафте.
Особый интерес в этом плане представляет собой регион Лофотенских островов (с островами Вестеролен), расположенных за полярным кругом (около 68–69° с.ш.) на северо-западном побережье Скандинавии, омываемых и отепляемых Гольфстримом. По палинологическим данным [3, 11, 12], значительное распространение популяций Р. sylvestris выявлено в этом регионе уже около 9500 лет BP (т.е. до 1950 г. – условной даты современности). Однако вряд ли это произошло вследствие невероятно быстрого (свыше 1000 км/1000 лет!) расселения сосны сюда из Альп, через пролив Каттегат, который появился не ранее 10 700 лет BP [13]. Максимально возможная скорость анемохорного распространения ее популяций в горах Скандинавии (в сочетании с более быстрой гидрохорией по течению рек) составляет 250–300 км/1000 лет [14]. В течение менее чем 1000 лет (с 10 500 до 9500 лет BP) вряд ли они могли бы успеть расселиться с крайнего юга Скандинавии вдоль побережий Северного моря далее района г. Бергена, отстоящего от Лофотенских островов еще на 1000 км. По-видимому, исключена также и вероятность быстрой реколонизации популяций сосны в этот регион с северо-запада Русской равнины, в обход ледника через Лапландию, где они появились лишь около 8000 лет BP [1, 3]. Между тем палеоботанические спектры озерных и болотных отложений на побережье Лофотенских островов свидетельствуют о появлении Р. sylvestris там уже 9500–13 000 лет ВР [3, 11, 12], а ее макрофоссилий – даже 19 200–22 000 лет ВР [6].
Некоторые авторы сомневаются в вероятности выживания хвойных и нахождения их автохтонных макрофоссилий в зоне тундры [15]. В то же время исследования по палеогляциологии [13] и палеомаринологии [16] выявили вероятность постоянного отепляющего воздействия течений палео-Гольфстрима на зону шельфа Лофотенских островов в течение “последнего ледникового максимума” (LGM). Недавно появились и данные по геногеографии Picea abies [6] и Pinus sylvestris [9], отражающие генетическую специфику лофотенских популяций этих видов.
К настоящему времени накоплены разносторонние факты, позволяющие предположить, что в течение последней гляциальной фазы плейстоцена в Скандинавии существовал и в голоцене послужил источником расселения популяций P. sylvestris L. один из крайне северных рефугиумов этого вида. Однако все эти данные еще во многом противоречивы, дискуссионны и слабо аргументированы на количественном уровне.
Задача настоящей работы – анализ и обобщение данных по хронологии и темпам голоценовой реколонизации в регион Лофотенских островов Скандинавии популяций P. sylvestris из окружающих перигляциальных зон, а также по палеоэкогеографии, генетике и экологии популяций сосны этого региона с целью верификации гипотезы об их автохтонном рефугиальном происхождении и расселении в голоцене.
ОБЪЕКТЫ И МЕТОДЫ
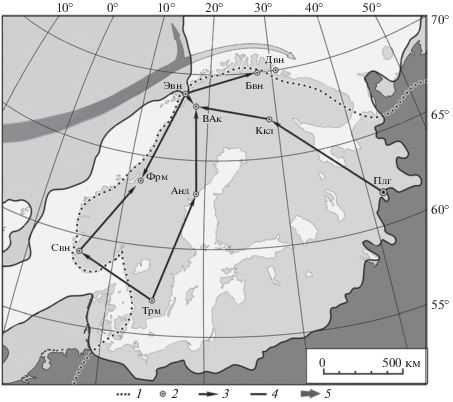
Объектами сравнительного хроно-палинологического изучения времени и скорости реколонизации популяций P. sylvestris в голоцене в направлении Лофотенских островов служили три их группы, расположенные вдоль наиболее вероятных путей расселения вслед за отступающим скандинавским ледником из его перигляциальной зоны (рис. 1): 1) с юга Скандинавии (Труммен) на северо-запад вдоль побережий Северного моря, а затем на северо-северо-восток по побережью Норвежского моря (Труммен–Сандвикватн–Форрамурене–Вуолеп-Аллакасьяурэ); 2) с юга Скандинавии на северо-северо-восток по западному побережью Балтики до Ботнического залива, а далее на север через Скандинавские горы (Труммен–Андунсьо–Вуолеп-Аллакасьяурэ); 3) с северо-запада Русской равнины на запад-северо-запад через Фенноскандию (Паденга–Кааккурилампи–Вуолеп-Аллакасьяурэ).
Рис. 1.
Картосхема расселения в голоцене популяций P. sylvestris из Центральной и Северо-Восточной Европы по альтернативным путям в регион Лофотенских островов Скандинавии. 1 – границы ареала P. sylvestris, 2 – локализация палиноспектров и даты появления доли пыльцы в них ≥20%, 3 – направления расселения популяций, 4 – границы максимального покровного оледенения в последней гляциальной фазе (22 000 лет ВР), 5 – схема палео-Гольфстрима. Шифры палиноспектров: Анд – Андунсьо, Бвн – Бруватнет, ВАк – Вуолеп-Аллакасьяурэ, Двн – Домсватнет, Ккл – Куаккурилампи, Пдг – Паденга, Свн – Сандвикватн, Трм – Труммен, Фрм – Форрамуренэ, Эвн – Эндлеватн.
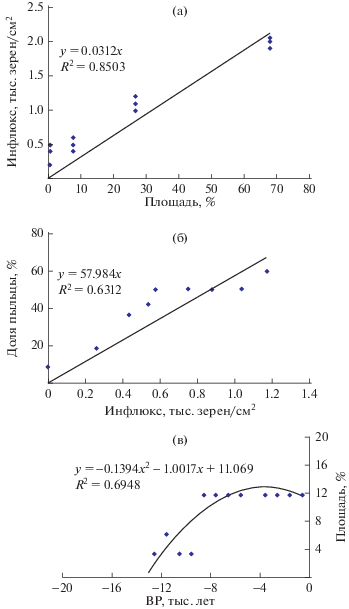
Дата иммиграции авангардных популяций P. sylvestris в тот или иной пункт ареала определена (с точностью 0.5 тыс. лет) на основе данных палинологии по радиодатированному времени появления достаточно репрезентативной и возрастающей доли ее пыльцы (≥20% общего количества) в палиноспектре древесных растений. Этот условный норматив, соответствующий примерно 10% площади лесов с доминированием сосны в радиусе 100 км вокруг пункта палиноанализа, принят нами на основании регрессионного анализа результатов исследовандий Х. Хиверинена [1] абсолютного содержания пыльцы сосны (зерен в 1 см3) и ее доли (%) в общем палиноспектре древесных растений в редколесьях на северной границе P. sylvestris в Лапландии. Установлено, что инфлюкс ее пыльцы почти пропорционален (R2 = = 0.85) площади ее лесов в окружающем ландшафте (рис. 2а). Определены также достаточно тесные связи (R2 = 0.63) доли пыльцы (%) с ее инфлюксом в палиноспектре (рис. 2б) и площади сосновых лесов с долей пыльцы (рис. 2в).
Рис. 2.
Анализ связей между инфлюксом и долей пыльцы в палиноспектрах P. sylvestris на северной границе ее ареала в Лапландии (по данным исследований H. Hyvärinen [1]) и реконструкция хронологической динамики площади редколесий P. sylvestris в голоцене на Лофотенских островах (Эндлеватн): а – связь инфлюкса пыльцы с относительной площадью редколесий сосны (%) в радиусе 100 км от палиноспектра в период 1000–3000 ВР; б – связь доли пыльцы в дендроспектре с ее инфлюксом; в – реконструированная динамика площади редколесий сосны (%) на Лофотенских островах в голоцене.
Скорость распространения популяций сосны вычислена по палинологическим картам [3, 4, 10] как отношение расстояния между смежными палинопрофилями на трансекте к разности дат появления репрезентативной доли пыльцы в них. При этом учтены результаты исследований, проведенных нами в тех же регионах ранее [14]. Расстояния между пунктами палиноанализа, особенности рельефа суши, моря и направления течения рек определены по географическим координатам на базе программы Google Earth Pro. Сравнительный анализ параметров генетической дифференциации современных популяций P. sylvestris Лофотенской и смежных филогеографических групп Скандинавии проведен по данным европейских исследований структуры и полиморфизма их митохондриальной ДНК [9].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Рассмотрим хроногеографические особенности реколонизации популяций Р. sylvestris по альтернативным направлениям к региону Лофотенских островов из перигляциальной зоны Скандинавского ледника.
Юг Скандинавии–побережье Атлантики–Лофотенские острова (Труммен–Сандвикватн– Форрамуренэ–Вуолеп-Аллакасъярэ). Распространение популяций сосны с крайнего юга Скандинавии в этом направлении по побережьям вначале Северного, а затем Норвежского морей могло начаться не ранее 10 700 лет BP. В это время возник перешеек суши через пролив Каттегат [13], а индекс доли пыльцы сосны в палиноспектре Труммен достиг 30% (рис. 3а).
Рис. 3.
Хронопалинологический анализ динамики голоценовой реколонизации попопуляций P. sylvestris в регион Лофотенских островов Скандинавии по альтернативным путям из Центральной и Северо-Восточной Европы: а – Труммен–Сандвикватн–Форрамуренэ–Вуолеп-Аллакасьяурэ–Эндлеватн; б – Труммен–Андунсьо–Вуолеп-Аллакасьяурэ; в – Паденга–Куаккурилампи–Вуолеп-Аллакасьяурэ; 1 – доля пыльцы P. sylvestris в ее общем дендроспектре (%), 2 – первая репрезентативная (≥20%) доля пыльцы сосны, 3 – вероятные направления и даты расселения популяций при его максимальной скорости (в направлении от Эвн к ВАк и Фрм – по данным палиноспектров).
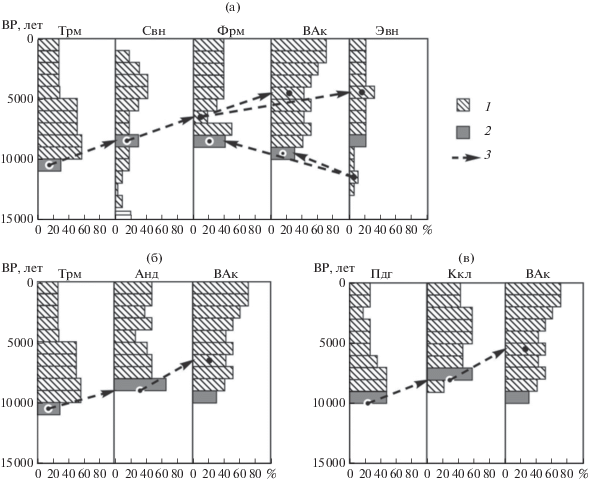
Быстрое (до 8–10 км/сут [17, 18]) “анемогидрохорное” распространение семян сосны по поверхности литоральных морских вод (с летальной для них соленостью) было исключено. Максимальная скорость их анемохорного расселения по побережьям и тем более через горы Скандинавии, вероятно, не превышала 250–300 км/1000 лет [8, 14]. Следовательно, даже для расселения сосны с юга Скандинавии (Труммен) до г. Ставамгер на юго-западе Норвегии (палиноспекр Сандвикватн) на расстояние около 600 км потребовалось бы как минимум 2000–2300 лет. Поэтому она достаточно репрезентативно появилась там не ранее 8500 лет BP [19].
Между тем довольно значимая доля пыльцы сосны (18%) была зафиксирована здесь уже около 10 500 лет BP (см. рис. 3а), а начиная с 14 000 лет BP отмечены ее первые заметные отложения (3–9%). Такое неожиданно раннее появление и быстрый рост доли пыльцы сосны на юго-западе Норвегии, синхронные с таковыми на юге Швеции, могут быть связаны с ее дальним заносом [20] или приближением массивов сосны, возможно, расселявшейся по перешейкам суши, возникавшим в Северном море в регионе Ютландии [9, 21].
В палиноспектре Форрамуренэ, расположенном на норвежском побережье, на 470 км далее к северу от пункта Сандвикватн, первое и сразу весьма значительное отложение пыльцы P. sylvestris (41%) наблюдается около 8500 лет BP ([22]; см. рис. 3а). По-видимому, это свидетельствует о его вторичном, “иммиграционном”, типе. Однако вероятность происхождения популяций сосны здесь в результате расселения с юго-запада региона г. Ставамгера исключается. Даже при максимальной скорости распространения ее популяций по побережью 300 км/1000 лет они могли бы появиться в регионе Форрамуренэ не ранее чем 7000 лет BP, т.е. на 1500 лет позднее даты их фактического появления и начала быстро возрастающего вклада их пыльцы в дендроспектр (до 50% в период 7000–8000 лет ВР).
Парадоксально раннее (9500 лет BP), вполне репрезентативное (36%) появление и экспоненциальный рост вклада пыльцы Р. sylvestris (до 51% около 7500 лет BP) установлены в дендропалинологическом спектре в горном регионе напротив Лофотенских островов (Вуолеп-Аллакасьяурэ, 68°10′, 18°10′ в.д. [11]; см. рис. 1 и 3а). Он находится на 450 км севернее палиноспектра Форрамуренэ, на высоте 995 м над ур. м., в 35 км к юго-востоку от г. Нарвика, расположенного на берегу фьорда. Кроме пыльцы сосны, в отложениях торфяника небольшого болота в углублении горного плато содержится примерно такое же количество пыльцы берез секций Nana и Albae. Вероятность раннего массового (36% пыльцы в дендроспектре) появления сосны здесь в результате расселения ее популяций с юга, из региона Форрамуренэ, исключается, так как даже при максимально возможной скорости распространения ее популяций (300 км/1000 лет) для этого потребовалось бы не менее 1500 лет. При этом они достигли бы Лофотенских островов лишь около 7000 лет BP, что на 2500 лет позднее палинологической даты появления сосны в окружающем ландшафте. На этом биогеографически достаточно достоверном основании можно сделать вывод о независимом от реколонизации с юга Скандинавии происхождении первичных популяций P. sylvestris в регионе Лофотенских островов.
Особо следует подчеркнуть, что в лофотенском палиноспектре достаточно репрезентативная доля пыльцы сосны (36%) установлена около 9500 лет ВР на высоте 995 м над ур. м. в горах, которые до 12 000–11 500 лет BP еще были покрыты льдами [13]. Однако в нижней части склонов Лофотенских островов – островов Вестеролен, омываемых и отепляемых водами Гольфстрима, небольшие пережившие гляциальную фазу группы деревьев (микропопуляции) сосны могли оставить и более ранние отложения пыльцы. На Южном Урале сосна на высоте 1120 м над ур. м. появилась примерно на 2000 лет позднее, чем на высоте 700 м [23].
В крайне низкопродуктивных перигляциальных редколесьях Р. sylvestris (Vб бонитета), которые сохранялись в лесотундре [1, 24, 25], плотность ее пыльцевого потока на порядки ниже, чем в бореальной зоне [26], а его следы в палиноспектрах голоцена почти отсутствуют. Кумулятивная пыльцевая продуктивность предтундровых редколесий Р. sylvestris или Larix sukaczewii, как и семенная, возрастает почти пропорционально повышению термообеспеченности климата и вегетативной продуктивности древостоя [26–29], а также площади их массивов. Тесная достоверная связь (R2 = 0.85; р = 0.03) инфлюкса пыльцы Р. sylvestris и в меньшей мере ее доли в палиноспектре c относительной площадью окружающих ее массивов выявлена нами по данным исследований Х. Хиверинена [1] в Лапландии (см. рис. 2а). На ее основе возможна достаточно надежная вероятностная реконструкция площади сосновых лесов и ее динамики по параметрам инфлюкса или даже доли пыльцы в седиментах (см. рис. 2в).
Можно предположить, что в период с 13 000 до 8000 лет BP в связи с повышением индекса изотопа кислорода, а также, вероятно, и температуры воздуха примерно на 25% [3] инфлюкс пыльцы вокруг распространяющихся массивов сосны постепенно возрастал. Именно этим, по-видимому, обусловлены отсутствие или крайне слабые следы пыльцы сосны в нижних слоях палиноспектров (ранее 10 000 лет BP) и последующий быстрый подъем ее доли не только в лофотенских, но и в других палиноспектрах болот и озер перигляциальных зон [3, 4, 10].
Эту гипотезу подтверждают данные стратиграфии пыльцевых отложений другого, более детально радиодатированного палиноспектра, проанализированного на одном из островов Вестеролен, расположенных на 100 км севернее Лофотенских (Эндлетватн, 69°14′ с.ш., 16°05′ в.д.; [12]; см. рис. 3а). Кроме вполне репрезентативного слоя пыльцы P. sylvestris (21%), относящегося к периоду 8000–9000 лет BP, в составе древесного палиноспектра здесь зафиксированы ее более ранние достаточно достоверные отложения (5–10%), начиная уже с 12 500 BP. Их источником в это время, когда почти вся Скандинавия и смежные регионы Европы в радиусе не менее 1500 км от Лофотенских островов были покрыты льдами [13], могли быть лишь местные автохтонные микропопуляции. Располагаясь на берегу островов, на высоте всего несколько десятков метров выше берега шельфа, они постоянно отеплялись водами одной из ветвей Гольфстрима [16].
На серии новейших палеогляциологических карт [13] хорошо видно, что в отличие от других частей береговой линии Атлантики граница открытого моря именно в регионе Лофотенских островов, и особенно островов Вестеролен, даже в фазы максимального оледенения (например, 22 000 лет BP) достигала их шельфа, отепляя его (см. рис. 1). В микропериоды же потепления климата в каждом тысячелетии она отступала здесь на сушу на расстояние до 10–15 км [6].
Вряд ли можно согласиться с гипотезой [15] о средних температурах воздуха летом на островах Вестеролен (Андойа) в последнем гляциале на уровне лишь около +1…+3°С. Она была принята таковой по аналогии с современной температурой в арктической тундре Земли Франца Иосифа лишь на основании доминирования в палеоседиментах Лофотена фоссилий видов тундровой растительности и фауны. Однако климат архипелага ЗФИ, расположенного вне зоны влияния Гольфстрима, не является аналогом климата палео-Лофотена. Наиболее подходящим здесь для реконструкции климата последнего гляциаля может быть современный климат юго-западного Шпицбергена (омываемого, как и Лофотенские острова, Гольфстримом), где температура воздуха в июле в арктической тундре (Свеагрува) летом в 80-е годы ХХ в. составляла +5…+6°С.
Вполне вероятно, что в западинных заболоченных местообитаниях на берегах озер региона Андойа в последнюю гляциофазу действительно доминировал мезоклимат тундры с температурой лета около +5°С, т.е. ниже экологического минимума для сосны. Но это не означает, что в обширном регионе Лофотенских островов – островов Вестеролен, в зоне их шельфа не было более теплых микростаций. Выживание растений определяет не некая средняя по региону, а реальная микроэкотопическая среда, которая в горном ландшафте Лофотена крайне вариабельна. Прежде всего высотный градиент летней температуры воздуха, равный около 0.8°С на каждые 100 м альтитуды, возрастает по мере приближения к морю. В защищенных же от арктических ветров ущельях фьордов, особенно на инсолируемых южных склонах с глубже протаивающими песчаными почвами, температура воздуха может быть как минимум на 2°С выше, чем в окружающих фоновых. По нашим наблюдениям, в лесотундре Западной Сибири температура суглинистой почвы (на глубине 50 см) на моренном плато в начале сентября была –0.7°С, а на южном склоне песчаной террасы на 3°С выше (+2.3°С). В горной тундре Урала температура воздуха на южном склоне на 3°С выше, чем на северном.
Наиболее высокая локальная теплообеспеченность могла создаваться в зонах узких (шириной менее 0.5 км), интенсивно турбулирующих проливов между западными островами Вестеролен и Лофотенскими, омываемыми водами глубокого и теплого (до 4–6°С зимой!) Норвежского течения. В их зонах, особенно на южных инсолируемых крутосклонах узких фьордов с песчаными почвами, обледенение территории на участках шириной до 2 км могло вообще отсутствовать [15], а температура воздуха летом достигать более +8…+10°С. В общем в периоды повышения летней температуры воздуха на 1.5–2.0°С более средней многолетней, обычные в каждом тысячелетии [29], она могла локально достигать уровня, достаточного для вызревания семян сосны – около +10…+11°С [20, 24]. Таким образом, вероятно, обеспечивались не только выживание, но и апериодическое возобновление микропопуляций сосны в течение всей гляциальной фазы.
Реконструкция хронодинамики редколесий сосны по данным палиноспектра [12] на островах Вестеролен (Андойа, Эндлеватн) на основании выявленных нами связей (см. рис. 2) показывает быстрый рост их относительной площади – с 1.2 до 12.7% – по мере потепления климата в период с 12 500 до 4500 лет ВР. Это отражает закономерный процесс возобновления и расселения автохтонных рефугиальных популяций сосны в лофотенском регионе с самого начала голоцена – еще задолго до инвазии ее адвентивных поселений.
Юг Скандинавии–побережье Балтики–Лофотенские острова (Труммен–Андунсьо–Вуолеп-Аллакасьяурэ). Судя по дате появления репрезентативной доли пыльцы сосны в Труммене (наиболее вероятно около 10 500 лет BP) [3, 30] и дате (около 9000 лет BP) ее массового появления (63%) на расстоянии 760 км (Анундсьо, 63°30′ с.ш., 18°12′ в.д.) на западном берегу Ботнического залива (рис. 3б), скорость распространения сосны на данном этапе составляла 507 км/1000 лет. Эта величина почти в 4 раза больше максимальной вероятной скорости расселения популяций этого вида (120–140 км/1000 лет) в оптимальных условиях климата, субстрата и семеношения P. sylvestris в центре ее ареала [31].
Быстрое распространение сосны вдоль побережья Балтики можно объяснить на основании гипотезы гидрохории семян [31] P. sylvestris по поверхности пресного Иольдиева моря, существовавшего в период с 10 700 по 8000 лет BP. Экспериментально показано, что при скорости ветра 5 м/с семена P. sylvestris под влиянием парусности их крылаток “анемогидрохорно” распространяются по поверхности озера со скоростью 8–10 км в сутки [17]. За трое суток попутного ветра они могут уплыть на 27–30 км. При наступлении генеративной стадии деревьев сосны в 30–35 лет скорость расселения ее популяций вдоль берегов пресных водоемов составит около 1000 км/1000 лет. В действительности происходит “эстафетное” распространение семян хвойных, при котором их гидрохория чередуется с анемохорией.
Дальнейший путь реколонизации популяций сосны от берегов Ботнического залива (Андунсьо) к региону Лофотенских островов пролегал на север через горные хребты и гольцовые тундры Скандинавии в направлении, противоположном течению рек. Следовательно, быстрая гидрохория семян вдоль них здесь была исключена. Ранее нами было определено [14], что в подобных условиях средняя скорость продвижения популяций сосны в горах Скандинавии, даже при частичном участии гидрохории вдоль рек, не превышает 250–300 км/1000 лет, а при ее отсутствии, как и у ели европейской [18], примерно вдвое меньше – 130–150 км/1000 лет. Поэтому расстояние в 515 км до лофотенского региона, через горный водораздел с мерзлотными почвами, популяции сосны с прибалтийского побережья могли преодолеть не менее чем за 2500–3000 лет, а достигнуть регионов Лофотена не ранее 6000–6500 лет BP. Это на 3000–3500 лет позднее даты репрезентативного появления ее пыльцы во вторичном горном палиноспектре Вуолеп-Аллакасьяурэ (см. рис. 3б).
Северо-запад Русской равнины–Фенноскандия–Лофотенские острова (Паденга–Кааккурилампи–Вуолеп-Аллакасьяурэ). По данным палинологического спектра [32], авангардная перигляциальная популяция Паденга на северо-западе Русской равнины (61°45′ с.ш., 42°45′ в.д.) появилась около 10 000 лет BP (рис. 3в). Позднее с повышением температуры воздуха популяции P. sylvestris сравнительно быстро распространялись вслед за отступающим ледником. Это было возможно как анемохорно, так и гидрохорно – по течению рек Северная Двина, Онега и других, текущих в северо-западном направлении, а в Фенноскандии – и по поверхности ледниковых озер “Суоми”, вытянутых в том же направлении.
Судя по пыльцевому спектру Кааккурилампи (67°03′ с.ш., 28°56′ в.д., [33]), репрезентативное обилие пыльцы (57%) здесь появилось около 8000 лет BP (см. рис. 3в). На расстояние 885 км от Паденги до Кааккурилампи сосна продвинулась за 2000 лет с довольно высокой скоростью – около 440 км/1000 лет.
Далее по мере отступления ледника (позднее 9000 лет ВР, [13]) популяции сосны расселялись к Лофотенским островам через север Скандинавского хребта. Хотя, вероятно, этому препятствовали встречное течение рек и вплоть до 9000 лет ВР северная кромка отступающего ледника с мерзлотной перигляциальной гольцово-тундровой зоной [13]. На заключительном этапе иммиграции в регион Лофотена длиной 470 км скорость распространения вряд ли превышала 150–200 км [14]. Поэтому сосна могла достичь гор Лофотена (Вуолеп-Аллакасьяурэ) не ранее чем через 2500–3000 лет, т.е. около 5000–5500 лет BP. Это, как минимум, на 4000–4500 лет позднее установленной даты (9500 лет BP) массового (30%) появления ее пыльцы в лофотенском палиноспектре ([11]; см. рис. 2в).
Таким образом, ни по одному из трех рассмотренных выше гипотетичных направлений иммиграции популяций Р. sylvestris в регион Лофотенских островов они не могли достичь его, даже при максимально возможных вероятных темпах их расселения, ранее дат 4500–6500 лет BP, т.е. на 3000–5000 лет позднее даты репрезентативного массового появления в палиноспектрах и начала расселения сосны в окружающем ландшафте (9500 лет BP). Следовательно, в регионе Лофотенских островов Скандинавии в течение по крайней мере последней гляциальной фазы плейстоцена непрерывно выживали и воспроизводились ее автохтонные рефугиальные микропопуляции.
Генетическая специфика лофотенских популяций Р. sylvestris. Хронопалинологические свидетельства выживания и самодостаточного независимого постгляциального возобновления и расселения популяций сосны в Лофотенском регионе подтверждают новейшие данные географии митохондриальной ДНК [9]. Проведенный нами анализ параметров состава гаплотипов и полиморфизма лофотенской филогеногеографической группы популяций P. sylvestris выявил их достоверные отличия (р ≤ 0.004–0.011) от смежных юго-западно-скандинавской и западно-прибалтийской (“шведской”) групп (табл. 1). Все популяции лофотенской группы однородны (моногаплотипны) по митотипу а во всем ареале их приоритетного расселения. Как показано выше, вероятно, он сформировался задолго до внедрения адвентивных популяций сосны в Скандинавию (в период с 9000 до 6000 лет ВР) – на расстоянии до 500 км от центра региона по побережьям Атлантики (на юг и север) и до 150 км на восток – до водораздела скандинавского хребта. В то же время в спектрах обеих соседствующих групп популяций Скандинавии содержится от 2.7 до 14.0% гаплотипа b, характерного для адвентивных популяций, распространившихся с юга и юго-востока периферии Скандинавского ледника (см. табл. 1). На юго-западе Норвегии гаплотип b изредка встречается лишь до 64.5° с.ш. – вплоть до зоны полного доминирования лофотенского моногаплотипа a, расселившегося сюда на 2000 лет ранее (см. рис. 3). Таким образом, лофотенская группа популяций P. sylvestris, приоритетным центром происхождения которой могли быть лишь автохтонные популяции, генетически резко дифференцирована от остальных скандинавских, иммигрировавших из Центральной и Восточной Европы [4, 9, 14, 34].
Таблица 1.
Дифференциация средних параметров митохондриальной ДНК между филогеографическими группами популяций P. sylvestris в Скандинавии (по данным [9])
| Филогеографи-ческие группы популяций | Параметры митохондриальной ДНК | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| гаплотип а | гаплотип b | Hmt | ||||||||||
| Mx ± m | Δ | t | p | Mx ± m | Δ | t | p | Mx ± m | Δ | t | p | |
| Юго-Западная Норвежская | 14.6 ± 0.56 | 0.4 ± 0.56 | 0.052 ± 0.072 | |||||||||
| 0.4 | 1.71 | 0.105 | 0.4 | –1.71 | 0.105 | 0.052 | –1.43 | 0.171 | ||||
| Лофотенская | 15.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 | |||||||||
| 2.1 | –3.06 | 0.004 | 2.1 | 2.88 | 0.006 | 0.216 | 3.37 | 0.002 | ||||
| Шведская | 12.9 ± 1.82 | 2.1 ± 1.82 | 0.216 ± 0.175 | |||||||||
Можно предположить, что в настоящее время в итоге длительной репродуктивной изоляции, экотопического отбора, а также дрейфа генов (в экстремальных условиях дефицита тепла и численности особей) и других факторов микро-эволюции сформировалась особая географическая раса вида Pinus sylvestris L., var. lofotenica. Она может значительно отличаться от других популяций P. sylvestris экологической нишей, в том числе меньшим (возможно, на 1.5–2.0°С) минимумом летней температуры воздуха для вызревания семян. Установлено, что нормы реагирования генетических рас видов на факторы среды широко вариабельны: например, оптимальная влажность субстрата для прорастания семян P. sylvestris лесной зоны Зауралья составляет 20%, а семян степных популяций Тургая – лишь 12%; рост корней сосны северотаежных популяций начинается при +2.5…+3.0°С, а лесостепных – при +5°С [17].
В зонах контактов и гибридизации лофотенской группы популяций сосны с инвазионными европейскими возможны достоверные градиенты, а на Скандинавском водоразделе даже вполне отчетливые границы их генофонда, аналогичные таковым на хребте Большого Кавказа [35]. Однако для верификации этих гипотез необходимы более детальные междисциплинарные – палеогеографические, популяционно-генетические, экологические и фенотипические – исследования.
ЗАКЛЮЧЕНИЕ
В итоге хронопалинологического анализа голоценовой реколонизации популяций Pinus sylvesris L. в регион Лофотенских островов Скандинавии из ее перигляциальной зоны установлено, что даже при максимальной скорости распространения (300 км/1000 лет) они не могли достичь этого региона ранее 6500–7000 лет BP. Это, как минимум, на 2500–3000 лет позднее фактической даты (9500 лет ВР) репрезентативного появления пыльцы сосны (≥20%) в местном горном палиноспектре, а тем более на низких берегах проливов и фьордов. Таким образом, вероятность происхождения сосны здесь от адвентивных популяций, иммигрировавших из регионов Центральной и Восточной Европы, и их приоритетного вклада в формирование ее местного генофонда исключается.
Появление и постепенный рост доли пыльцы P. sylvestris в озерных седиментах Лофотенских островов–островов Вестеролен, начиная уже с раннего голоцена (12 500 лет ВР), когда вся остальная Скандинавия в радиусе 1500 км от них была покрыта льдами, а также находки макрофоссилий Pinus sp. давностью 22 000–17 700 лет ВР с достаточной вероятностью свидетельствуют о постоянном существовании здесь микропопуляций сосны в течение последней гляциальной фазы и их прогрессирующем расселении в голоцене по мере потепления климата.
Анализ новейших данных по геногеографии лофотенской группы современных популяций P. sylvestris свидетельствует об ее отчетливой генетической специфичности и расселении в голоцене в радиусе до 500 км. Можно предположить, что в итоге длительной изоляции и микроэволюции в условиях Крайнего Севера ареала здесь сформировалась особая географическая раса Pinus sylvestris L., var. lofotenica (nova nom. prov.), менее требовательная к дефициту тепла для выживания и возобновления популяций.
Известные в настоящее время палеоданные гляциологии, маринологии, климата и ботаники по региону Лофотенских островов, во многом обусловленные непрерывным отепляющим влиянием палео-Гольфстрима, в целом также подтверждают или, по крайней мере, не опровергают гипотезу о существовании и постгляциальном расселении автохтонной лофотенской рефугиальной группы популяции Pinus sylvestris L., хотя, безусловно, нуждаются в разносторонней верификации.
Работа выполнена в рамках государственного задания Ботанического сада УрО РАН (номер государственной регистрации: АААА-А17-117072810011-1) при поддержке РФФИ (проект № 16-04-00948).
Авторы подтверждают отсутствие конфликта интересов.
Список литературы
Hyvärinen H. Absolute and relative pollen diagrams from northernmost Fennoscandia // Fennia. 1975. V. 142. 23 p.
Bennett K.D., Tzedaki P.C., Willis K.J. Quarernary refugia of north European trees // J. of Biogeography. 1991. V. 18. P. 103–115.
Lang G. Quartäre Vegetationsgeschichte Europas: Methoden und Ergebnisse. Jena, Stuttgard, New York: Gustav Fischer Verlag, 1994. 462 S.
Cheddadi R., Vendramin G.G., Litt T. et al. Imprints of glacial refugia in the modern genetic diversity of Pinus sylvestris // Global Ecol. Biogeogr. 2006. V. 15. P. 271–282.
Pihäjärvi T., Salmela M.J., Savolainen O. Colonization routes of Pinus sylvestris inferred from distribution of mitochondrial DNA variation // Tree genetics and genomes. 2008. V. 4. P. 247–254.
Parducci L., Jø´rgensen T., Tollefsrud M.M. et al. Glacial survival of boreal trees in Northern Scandinavia // Science. 2012. V. 335. P. 1083–1086.
Buchovska J., Danusevičius D., Baniulis D. et al. The location of the north glacial refugium of Scots pine based on mitochondrial DNA markers // Baltic Forestry. 2013. V. 19. P. 2–12.
Санников С.Н., Петрова И.В., Егоров Е.В., Санникова Н.С. Выявление системы плейстоценовых рефугиумов Pinus sylvertris L. в южной маргинальной зоне ареала // Экология. 2014. № 3. С. 174–181. [Sannikov S.N., Petrova I.V., Egorov E.V., Sannikova N.S. A system of pleistocene refugia for Pinus sylvestris L. in the southern marginal part of the species range // Rus. J. of Ecology. 2014. V. 45. No. 3. P. 167–173.]
Dering M., Kosiński P., Wyka T.P. et al. Tertiary remnants and Holocene colonizers: genetic structure and phylogegraphy of Scots pine reveal higher genetic diversity in young boreal than in relict Mediterranian populations and a dual colonization of Fennoscandia // Diversity and distributions. 2017. V. 23. P. 540–555.
Huntley B., Birks H.J. An atlas of past and pollen maps for Europe: 0–13000 years ago. Cambridge: Cambridge Univ. Press, 1983. 667 p.
Küttel M. Vuolep Allakacjaure – eine pollenanalytische Studie zur Vegetationgseschichte der Tundra in Nordschwedewn // Festschrift Max. Welten. Diss. Bot. Vaduz. 1984. S. 191–212.
Vorren K.-D., Alm T. An attempt at synthesizing the Holocene biostratigraphy of a “type area” in northern Norway by means of recommended methods for zonation and comparison of biostratigraphical data // Ecol. Medit. 1985. V. 11. P. 53–64.
Hughes L.C.A., Gyllencreutz R., Lohne Ø´.S. et al. The last Eurasian ice sheets – a chronological database and time-slice reconstruction. DATED-1 // Boreas. 2016. V. 45. P. 1–45.
Санников СН., Санникова Н.С. Пути и темпы реколонизации Pinus sylvestris L. и видов Picea в Скандинавии в голоцене // Журн. общ. биол. 2015. Т. 76. № 6. С. 475–481.
Vorren T.O., Vorren K.-D., Aasheim O. et al. Palaeoenvironment in northern Norway between 22.2 and 14.5 cal. ka BP // Boreas. 2013. V. 42. P. 876–895.
Ezat M.M., Rasmussen T.L., Groeneveld J. Persistent intermedia water warming during cold stadials in the sourtheastern Nordic seas during the past 65 k.y. // Geology. 2014. V. 42. P. 663–666.
Санников С.Н., Санникова Н.С., Петрова И.В. Очерки по теории лесной популяционной биологии. Екатеринбург: УрО РАН, 2012. 272 с.
Hafsten U. The immigration and spread of spruce forest in Norway, traced by biostratigraphical studies and radiocarbon dating. A preliminary report // Norsk. geogr. Tidskr. 1985. V. 39. P. 99–108.
Paus A. Late Weicheselian vegetation, climate, floral migration at Sandvicvatn, North Rogaland, southwestern Norway // Boreas. 1988. V. 17. P. 113–139.
Санников С.Н., Петрова И.В. Дифференциация популяций сосны обыкновенной. Екатеринбург: УрО РАН, 2003. 247 с.
Coles B.J. Doggerland: the cultural dynamics of a coast-line // Geolog. Soc. Special Publications. 2000. V. 175. P. 393–401.
Hafsten U., Solem T. Age, origin and paleo-eclogical evidence of blanket bogs in Nord-Trondelag, Norway // Borras. 1976. V. 5. P. 119–141.
Панова Н.К. История лесной и болотной растительности центральной горной провинции Южного Урала в голоцене (по данным споро-пыльцевого анализа торфа): Автореф. дис. … канд. биол. наук. Свердловск, 1987. 23 с.
Renvall A. Die periodischen Erscheinungen der Reproduktion der Kiefer an der polaren Waldgrenze // Acta Forest. Fenn. 1912. V. 1. P. 1–154.
Гричук В.П. История флоры и растительности Русской равнины в плейстоцене. М.: Наука, 1989. 183 с.
Санников С.Н., Петрова И.В., Санникова Н.С. О более надежном методе определения времени поселения Pinus sylvestris (Pinaceae) в местном ландшафте // Бот. журнал. 2009.Т. 94. № 1. С. 128–134.
Sarvas R. Investigations on the flowering and seed crop of Pinus sylvestris // Comm. Inst. Forest. Fenn. 1962. V. 53. № 4. P. 1–198.
Петрова И.В., Санников С.Н. Изоляция и дифференциация популяций сосны обыкновенной. Екатеринбург: УрО РАН, 1996. 159 с.
Хантемиров Р.М. Динамика древесной растительности и изменения климата на севере Западной Сибири в голоцене: Автореф. дис. … докт. биол. наук. Екатеринбург, 2009. 43 с.
Digerfeldt G. The post-glacial development of Lake Trummen. Regional vegetation history, water level changes and paleolimnology // Folia Limnol. Scand. 1972. V. 16. P. 1–104.
Санников С.Н., Санникова Н.С. Гипотеза гидрохорного расселения популяций хвойных древесных растений // Экология. 2007. № 2. С. 83–87.
Никифорова Л.Д. Изменение природной среды в голоцене на Северо-Востоке. Европейской части СССР. Автореф. дис. … канд. геогр. наук. М.: МГУ, 1980. 17 с.
Sorsa P. Pollenanalytische Untersuchungen zur spätquartären Vegetations- und Klimaentwicklung im östlichen Nordfinnland // Ann. Bot. Fenn. 1965. V. 2. S. 301–413.
Sannikov S.N., Paule L., Yegorov E.V. Identification of probable Pleistocene refugia of Pinus sylvestris L. in Europe // Forest Genetics. 2013. V. 13(4). P. 201–206.
Петрова И.В., Санников С.Н., Темботова Ф.А. и др. Геногеография популяций Pinus sylvestris L. Большого Кавказа и Крыма // Экология. 2017. № 6. С. 431–439.
Дополнительные материалы отсутствуют.