

Экология, 2019, № 3, стр. 236-239
Демографическая структура ценопопуляций Scutellaria baicalensis Georgi в связи с климатическими градиентами и локальными факторами
Д. В. Санданов a, *, С. Росбах b, **
a Институт общей и экспериментальной биологии СО РАН
670047 Улан-Удэ, ул. Сахьяновой, 6, Россия
b University of Regensburg
93040 Regensburg, Germany
* E-mail: denis.sandanov@gmail.com
** E-mail: sergey.rosbakh@ur.de
Поступила в редакцию 23.10.2018
После доработки 05.12.2018
Принята к публикации 17.12.2018
Изучение параметров популяций растений в различных частях ареала позволяет выявить их отклик на пространственные и временны́е изменения экологических условий. В последние годы широко исследуется влияние различных экологических градиентов на демографическую структуру популяций растений [1–3] и продолжительность онтогенеза у растений [4]. Помимо макроклиматических факторов, влияние на демографию популяций могут оказывать и локальные воздействия, в частности выпас [5]. Для некоторых видов на широтном градиенте при низких температурных показателях наблюдается снижение выживаемости растений при повышении репродуктивных параметров. Этот феномен был назван “демографической компенсацией” [6] и связан с тем, что в экстремальных условиях у растений увеличивается продолжительность онтогенеза вследствие ограничения их размножения факторами среды [4].
Большинство современных исследований посвящено анализу различных аспектов структуры популяций, включая соотношение возрастных групп или параметров репродукции, роста и жизнеспособности популяций [7, 8 и др.]. В работе Л.А. Животовского [9] приводятся результаты применения индексов возрастности (Δ) и эффективности (ω) при изучении ценопопуляций растений и отмечается, что данные индексы непосредственно не связаны с экологическими параметрами и определяются возрастной структурой популяции. В последние годы проведен ряд исследований по оценке влияния экологических факторов на онтогенетическую структуру ценопопуляций, в том числе в разных частях ареала [10, 11]. Цель наших исследований – количественная оценка влияния макроклиматических показателей и локальных факторов на онтогенетическую структуру ценопопуляций шлемника байкальского Scutellaria baicalensis Georgi – вида восточноазиатской приуроченности, третичного реликта и ценного лекарственного растения.
Scutellaria baicalensis Georgi – стержнекорневой многоглавый травянистый многолетник с монокарпическими побегами. Для вида характерно только семенное размножение. Выделение онтогенетических состояний проводилось на основе исследований Ю.А. Банаевой [12] с дополнениями Т.Г. Бухашеевой [13]. Демографическую структуру 53 ценопопуляций (ЦП) Scutellaria baicalensis изучали с 2006 по 2017 г. в разных частях ареала вида: Забайкальский край и Дальний Восток России, Восточный аймак Монголии и провинция Внутренняя Монголия Китая. Помимо собственных исследований, в анализ были вовлечены литературные данные по 9 ЦП с Забайкальского края и Дальнего Востока России, приведенные в работе Ю.А. Банаевой [12] (рис. 1). Изучены две группы факторов, оказывающих воздействие на онтогенетическую структуру популяций S. baicalensis: ряд различных макроклиматических переменных и индексов (биоклиматические переменные BIOCLIM [14], индексы ENVIREM [15]) и локальные факторы (увлажненность местообитаний, богатство–засоленность почвы, высота над уровнем моря, проективное покрытие и видовая насыщенность фитоценозов, выпас). При расчете экологических статусов растительных сообществ для каждой ценопопуляции использовали экологические шкалы [16]. Интенсивность выпаса оценивали по 5-балльной шкале пастбищной дигрессии (градации: 1 – влияние выпаса очень слабое или отсутствует, 2 – слабый выпас, 3 – умеренный выпас, 4 – сильный выпас, 5 – сбой). Для изучения демографической структуры ЦП в каждой из них на трансекте закладывали 25–45 пробных площадок размером 1 м2. На основе полученных данных были рассчитаны следующие индексы: возрастности [17], восстановления и замещения [18], старения [19] и эффективности [9]. Взаимозависимость факторов среды и параметров структуры ценопопуляций изучали с помощью ранговых корреляций по Спирмену. Статистическую обработку данных и визуализацию результатов проводили в среде R версия 3.5.0. [20].
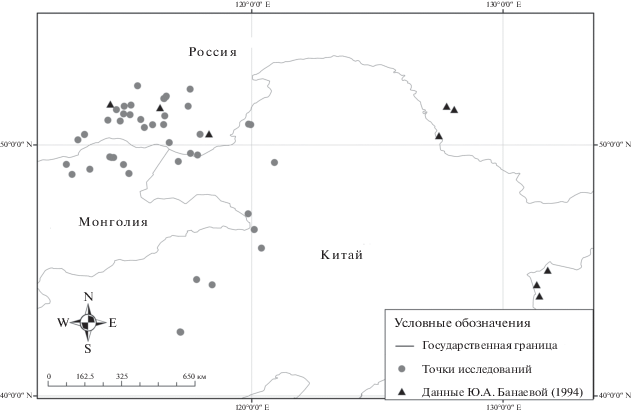
Проведенный анализ показал увеличение доли виргинильных особей S. baicalensis с возрастанием аридности местообитаний и сенильных особей при повышении континентальности климата (рис. 2). Эти тренды являются взаимосвязанными, так как на изучаемой территории с увеличением континентальности климата возрастает и его аридность. Соответственно в более аридных (и более континентальных) местообитаниях на северо-западе ареала растет доля виргинильных и сенильных особей и вследствие этого сокращается численность генеративных растений. Для возрастной структуры ценопопуляций S. baicalensis не менее важное значение имеют и локальные условия. В частности, с ростом абсолютной высоты наблюдается снижение доли ювенильных особей и возрастание доли генеративных растений, а повышение интенсивности выпаса приводит к увеличению доли сенильных особей (см. рис. 2). Для фитоценозов с высоким проективным покрытием и участием S. baicalensis отмечается снижение доли виргинильных особей вида.
Рис. 2.
Корреляционные связи факторов среды и демографических показателей: j – доля ювенильных растений; v – доля виргинильных растений; g – доля генеративных растений; sen – доля сенильных растений; дельта – индекс возрастности (Δ); омега – индекс эффективности (ω); Iв – индекс восстановления; Iст – индекс старения; Iз – индекс замещения; теплообеспеченность – cумма среднемесячных температур для месяцев со средней температурой выше 0°С; аридность – индекс аридности Торнвайта; континентальность – индекс континентальности; высота – высота над уровнем моря; увлажнение – средний экологический статус сообщества по шкале увлажнения; богатство почв – средний экологический статус сообщества по шкале богатства–засоленности почв; ОППтрав – проективное покрытие травяного яруса; число видов – число видов в описании (видовая насыщенность сообщества). Цифрами обозначены коэффициенты корреляции по Спирмену; эти же показатели дублируются графически в виде кругов (размер круга зависит от силы корреляционных связей), цветовые градации соответствуют шкале.
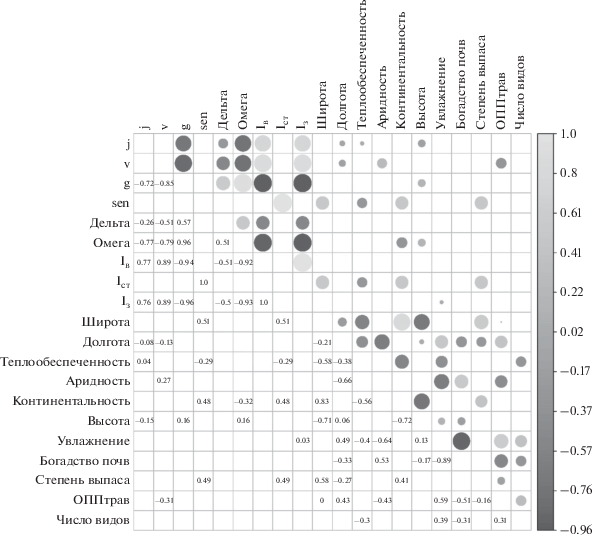
Не обнаружено и наличие “демографической компенсации”, так как в районе исследований в более высоких широтах с понижением показателей температуры также возрастает и аридность, под воздействием которой снижается численность генеративных особей, обладающих высокой биомассой и семенной продуктивностью. Схожие тренды для периферических популяций изучаемого вида в Забайкалье были выявлены нами ранее [21]. Показано [2, 9, 22], что феномен “демографической компенсации” неявно выражен в краевых популяциях вида, а повышение аридности в значительной степени снижает рост популяции.
При увеличении абсолютной высоты наблюдается снижение доли ювенильных особей и возрастание доли генеративных растений. В Баварских Альпах выявлено [4], что на больших абсолютных высотах увеличиваются средний возраст и продолжительность онтогенеза многолетних травянистых растений. Возможно, что при больших диапазонах высот только генеративные особи S. baicalensis могут конкурировать с другими видами. Изучаемый нами вид в основном встречается в среднегорных степных ландшафтах, поэтому для него не характерно широкое распространение на больших высотах.
Высокая пастбищная дигрессия приводит к сокращению численности прегенеративных растений изучаемого вида [23, 24]. Основной угрозой, помимо поедания пастбищными животными, является и непосредственная элиминация молодых растений вследствие вытаптывания. Уплотнение почвы также может в значительной мере сократить семенное возобновление вида. В итоге при сохраняющейся интенсивности выпаса все больше генеративных растений переходит в сенильное возрастное состояние, а отсутствие пополнения молодыми особями приводит к регрессии ценопопуляции. Для изучаемого вида характерно только семенное размножение, поэтому в дальнейшем необходим детальный мониторинг численности и демографической структуры вида.
В результате проведенного анализа обнаружены слабые корреляционные связи между демографическими индексами и различными переменными. В большей степени выявлено влияние факторов среды на возрастные группы или отдельные онтогенетические состояния. Преобладание нормальных дефинитивных ценопопуляций S. baicalensis на всем протяжении ареала способствует тому, что их онтогенетическая структура остается относительно стабильной и соответственно близка к базовому спектру. Поэтому вариабельность рассматриваемых демографических индексов незначительная. Влияние локальных факторов, которые по силе воздействия сильнее, чем макроклиматические, может приводить к существенному изменению демографической структуры ценопопуляций S. baicalensis. Роль и влияние макроклиматических показателей могут быть выявлены при анализе онтогенетической структуры ценопопуляций видов с широким ареалом на выраженных экологических градиентах.
Исследования выполнены в рамках бюджетной темы № АААА-А17-117011810036-3 и поддержаны грантом РФФИ № 16-54-53057.
Список литературы
Vieira V.M.N.C.S., Oppliger L.V., Engelen A.H., Correa J.A. A new method to quantify and compare the multiple components of fitness – a study case with kelp niche partition by divergent microstage adaptations to temperature // PloS One. 2015. https://doi.org/10.1371/journal.pone.0119670
Sheth S.N., Angert A.L. Demographic compensation does not rescue populations at a trailing range edge // PNAS. 2018. V. 115. № 10. P. 2413–2418. https://doi.org/10.1073/pnas.1715899115
Villellas J., Garsia M.B. Life-history trade-offs vary with resource availability across the geographic range of a widespread plant // Plant Biology. 2018. V. 20. P. 483–489. https://doi.org/10.1111/plb.12682
Rosbakh S., Poschlod P. Killing me slowly: Harsh environment extends plant maximum life span // Basic and Applied Ecology. 2018. V. 28. P. 17–26. https://doi.org/10.1016/j.baae.2018.03.003
Lehndal L., Angren J. Herbivory differentially affects plant fitness in three populations of the perennial herb Lythrum salicaria along a latitudinal gradient // PloS One. 2015. https://doi.org/10.1371/journal.pone.0135939
Doak D.F., Morris W.F. Demographic compensation and tipping points in climate-induced range shifts // Nature. 2010. V. 467. P. 959–962. https://doi.org/10.1038/nature09439
Meyer A.H., Schmid B. Experimental demography of rhizome populations establishing clones of Solidago altissima // Journal of Ecology. 1999. V. 87. P. 42–54. https://doi.org/10.1046/j.1365-2745.1999.00317.x
Matias L., Jump A.S. Asymmetric changes of growth and reproductive investment herald altitudinal and latitudinal range shifts of two woody species // Global Change Biology. 2015. V. 21. P. 882–896. https://doi.org/10.1111/gcb.12683
Животовский Л.А. Онтогенетическое состояние, эффективная плотность и классификация популяций // Экология. 2001. № 1. С. 3–7. [Zhivotovsky L.A. Ontogenetic states, effective density, and classification of plant populations // Rus. J. of Ecology. 2001. V. 32. № 1. P. 1–5.] https://doi.org/10.1023/A:1009536128912
Валуйских О.Е., Тетерюк Л.В. Структура и динамика краевых популяций Gymnadenia conopsea (L.) R. Br. (Orchidaceae) на известняках Европейского Северо-Востока России // Экология. 2013. № 6. С. 420–427. [Valuiskikh O.E., Teteryuk L.V. Structure and dynamics of marginal Gymnadenia conopsea (L.) R. Br. (Orchidaceae) populations on limestones in the northeast of European Russia // Rus. J. of Ecology. 2013. V. 44. № 6. P. 461–467.] https://doi.org/0.7868/S0367059713060103
Подгаевская Е.Н., Золотарева Н.В. Особенности произрастания и состояние популяций Stipa pennata L. на северной границе распространения (Свердловская область) // Вест. Томского гос. ун-та. Биология. 2015. № 3 (31). С. 40–53. https://doi.org/10.17223/19988591/31/4
Банаева Ю.А. Шлемник байкальский (Scutellaria baicalensis Georgi) (экология, биология, интродукция): Дис. … канд. биол. наук. Новосибирск, 1994. 177 с.
Бухашеева Т.Г. Эколого-биологические особенности Scutellaria baicalensis Georgi в Забайкалье: Дис. … канд. биол. наук. Улан-Удэ, 2000. 179 с.
Hijmans R.J., Cameron S.E., Parra J.L. et al. Very high resolution interpolated climate surfaces for global land areas // International J. of Climatology. 2005. V. 25. P. 1965–1978. https://doi.org/10.1002/joc.1276
Title P.O., Bemmels J.B. ENVIREM: An expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche mode-ling // Ecography. 2018. V. 41. Issue 2. P. 291–307. https://doi.org/https://doi.org/10.1111/ecog.02880
Королюк А.Ю. Экологические оптимумы растений юга Сибири // Ботанические исследования Сибири и Казахстана. Барнаул; Кемерово, 2006. Вып. 12. С. 3–38.
Уранов А.А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов // Биологические науки. 1975. № 2. С. 7–33.
Жукова Л.А. Популяционная жизнь луговых растений. Йошкар-Ола : РИИК “Ланар”, 1995. 224 с.
Глотов Н.В. Об оценке параметров возрастной структуры популяций растений // Жизнь популяций в гетерогенной среде. Йошкар-Ола: МарГУ, 1998. Ч. 1. С. 146–149.
Team RDC. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing, 2018.
Санданов Д.В. Оценка состояния ценопопуляций восточноазиатских видов растений в различных частях ареала // Растительный мир Азиатской России. 2010. № 2 (6). С. 80–87.
Louthan A.M., Pringle R.M., Goheen J.R. et al. Aridity weakens population-level effects of multiple species interactions on Hibiscus meyeri // PNAS. 2018. V. 115. № 3. P. 543–548. www.pnas.org/cgi/doi/10.1073/pnas.1708436115
Санданов Д.В., Найданов Б.Б., Шишмарев В.М. Влияние региональных и локальных факторов на распространение и структуру популяций Scutellaria baicalensis Georgi // Вестн. Томского гос. ун-та. Биология. 2017. № 38. С. 89–103. https://doi.org/10.17223/19988591/38/5
Бухашеева Т.Г., Санданов Д.В., Асеева Т.А. и др. Возрастная структура ценопопуляций и сырьевая фитомасса Scutellaria baicalensis Georgi (Lamiaceae) // Растительные ресурсы. 2007. Т. 43. Вып. 4. С. 23–32.
Дополнительные материалы отсутствуют.