

Экология, 2019, № 4, стр. 309-315
Население энхитреид (Annelida, Clitellata, Enchytraeidae) и его зависимость от эдафических условий в рисовых агроэкосистемах России
М. И. Дегтярев a, b, Д. И. Коробушкин a, *, К. Б. Гонгальский a, b, А. С. Зайцев a, c, d
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
b Московский государственный университет им. М.В. Ломоносова
119992 Москва, Ленинские горы, 1, Россия
c Институт молекулярной медицины, Первый Московский государственный медицинский университет
им. И.М. Сеченова
119991 Москва, ул. Трубецкая, 8, Россия
d Институт экологии животных, Университет Юстуса Либиха
35392 Гиссен, Хаинрих-Буфф-Ринг 26, Германия
* E-mail: DKorobushkin@ya.ru
Поступила в редакцию 01.11.2018
После доработки 04.12.2018
Принята к публикации 23.01.2019
Аннотация
Традиционно считается, что сообщества почвенных животных (мезо- и макрофауны) на рисовых чеках фрагментарны и характеризуются крайне низкой численностью. Вместе с тем некоторые группы, например энхитреиды, могут становиться доминантными в условиях затопления, как было показано для тропических рисовых агроэкосистем. Однако в условиях умеренных широт исследования фауны и функциональной структуры энхитреид (Annelida, Clitellata, Enchytraeidae) не проводились. Нами изучено население и соотношение функциональных групп энхитреид в основных рисоводческих регионах России: Краснодарском и Приморском краях и Республике Калмыкия. Пробы были отобраны летом–ранней осенью 2016 г. в четырех типах местообитаний: на залитых рисовых чеках; чеках, занятых незаливными культурами; причековых валиках и ненарушенных травянистых местообитаниях (контроль). Анализ с применением обобщенной линейной модели показал, что фактор “тип местообитания”, а также его взаимодействие с фактором “регион” достоверно влияют на общую численность энхитреид, а отдельно взятый фактор “регион” не оказывает достоверного влияния. Среди местообитаний максимальная численность наблюдалась в контроле и на валиках (3278 ± 1131 и 3255 ± 762 экз/м2 соответственно; указаны среднее по всем регионам ± стандартная ошибка). Недостоверно ниже численность была на чеках, занятых незаливными культурами (1282 ± 850 экз/м2), минимальная – на залитых рисовых чеках (415 ± 323 экз/м2). Сравнение чеков, занятых незаливными культурами и рисом, показывает, что энхитреиды способны относительно быстро восстанавливать свою численность на рисовых чеках после их дренирования. Методом анализа главных компонент показано наличие положительной корреляции между численностью энхитреид, принадлежащих к базофильной экологической группе, и содержанием в почве подвижного фосфора. При этом корреляция численности энхитреид-ацидофилов с этим же показателем была отрицательной. Это свидетельствует о том, что соотношение численности экологических групп энхитреид выступает информативным индикатором концентрации биогенных элементов в почве и зависит от физико-химических характеристик рисовых почв. Можно утверждать, что, несмотря на невысокую численность в почве чеков, энхитреиды могут быть важным компонентом детритных пищевых сетей во время периодов дренирования чеков при интенсивном рисоводстве в умеренных широтах.
Почвенная мезофауна – один из важных, но малоизученных функциональных компонентов почв в рисовых агроэкосистемах. Беспозвоночные-редуценты вносят свой вклад в процессы разложения органики, в том числе и трудноразлагаемых пожнивных остатков риса [1], утилизация огромной массы которых представляет в настоящее время большую проблему [2]. Одним из семейств почвенных беспозвоночных, обитающих на рисовых полях, являются энхитреиды (Annelida, Clitellata, Enchytraeidae), чья роль в качестве эффективных редуцентов была подтверждена ранее для других агроэкосистем [3, 4]. Энхитреиды в почвах в основном исследуются как единая группа, без учета таксономического и экологического разнообразия. Их редко дифференцируют по таксономическим группам из-за относительной сложности определения [5], поэтому работы, посвященные выявлению и исследованию их функционального состава, редки [6], хотя различия в экологической специализации представителей разных родов семейства могут быть значительными [5, 7, 8].
Степень изученности фауны и населения энхитреид в России недостаточна – последние полномасштабные исследования датируются 80-ми годами прошлого столетия [9–11]. Особенно неполно изучены сообщества энхитреид южных регионов страны, при этом исследования в рисовых агроэкосистемах вообще не проводились. В мировой науке, напротив, в последнее время значительно возрос интерес к изучению роли энхитреид [12, 13] и олигохет в целом [14–16] как важных компонентов детритных пищевых сетей рисовых чеков.
Исследование роли энхитреид в формировании почвенного биоразнообразия и функционировании детритных пищевых сетей в рисовых агроэкосистемах крайне актуально, поскольку их разные экологические группы могут иметь различную чувствительность как к затоплению чеков, необходимому для выращивания риса в условиях умеренного климата, так и к изменению физико-химических характеристик почв на них. Более того, в России применяется система двух- или трехгодичного севооборота, при которой заливной рис чередуется с незаливными культурами или паром, что позволяет также исследовать возможность восстановления численности, функционального и таксономического разнообразия животных в условиях сильного антропогенного воздействия [17]. Одновременно могут быть получены данные о динамике и пространственной дифференциации численности и таксономического состава сообществ энхитреид регионов-производителей риса в России.
Цель настоящего исследования – изучение численности и таксономического состава энхитреид в трех основных регионах выращивания риса в России, а также отношения разных экологических групп энхитреид к различным типам местообитаний и пространственной дифференциации эдафических условий между ними в пределах рисовых агроэкосистем.
МАТЕРИАЛ И МЕТОДЫ
Исследования были проведены в трех ведущих рисоводческих регионах России:
1. Краснодарский край, рисовые чеки расположены к западу от г. Краснодара (45°04′ с.ш., 38°43′ в.д.), климат умеренно континентальный, среднегодовая температура составляет 11.5°С, среднегодовое количество осадков 675 мм. Рис в данной местности возделывается с 1930-х годов. Почвы – черноземы глинисто-иллювиальные (Calcic Phaeozems). Здесь и далее климатические параметры даны в соответствии с открытой базой данных Российского гидрометеорологического центра [18], а также с работой Пил с соавт. [19]. Названия типов почв предоставлены местными специалистами и приведены в соответствии с “Полевым определителем почв России” [20], а их международные названия даны согласно “Мировой реферативной базе почвенных ресурсов” [21].
2. Республика Калмыкия (далее “Калмыкия”): рисовые чеки расположены в районе п. Большой Царын (47°54′ с.ш., 45°21′ в.д.), климат семиаридный, среднегодовая температура воздуха 8.9°C, количество осадков 313 мм. Возделывание риса практикуется с 1960-х годов. Почвы – черноземы глинисто-иллювиальные (Haplic Phaeozems).
3. Приморский край (далее “Приморье”): рисовые чеки расположены в окрестностях г. Уссурийска (43°51′ с.ш., 131°56′ в.д.), климат муссонный, средняя температура 3.3°C, количество осадков 591 мм, почвы аллювиальные серогумусовые (Umbric, Histic Fluvisols). Возделывание риса ведется с 1930-х годов.
В Краснодарском крае и Калмыкии материал был отобран в июне, а в Приморье из-за логистических ограничений – в сентябре 2016 г. При этом значимого влияния сезона на численность и параметры населения энхитреид не выявлено (двухфакторный дисперсионный анализ, факторы “местообитание” и “дата отбора проб”, p > 0.05).
Пробные площадки были выбраны случайным образом на отстоящих друг от друга не менее чем на 1 км группах рисовых чеков в пределах четырех типов местообитаний (не менее трех в каждом регионе): залитый водой чек, засеянный рисом (далее “рис”); соседний чек, засеянный незаливной культурой – соя, люцерна, пшеница (далее “незаливная культура”); искусственный причековый валик высотой до 2 м и шириной 3–5 м, покрытый рудеральной травяной растительностью или тростником (далее “валик”); контрольное травянистое местообитание, расположенное в районе рисовых агроэкосистем (далее “контроль”). В Калмыкии контрольные местообитания характеризовались интенсивным выпасом скота, в то время как в других регионах выпаса не было.
Повторность для пробных площадок для каждого типа местообитаний в пределах каждого региона приведена в табл. 1. Число повторностей определяли наличием достаточного количества местообитаний, удовлетворяющих критериям отбора. При этом пробные площадки в типе местообитания “валик” выбирали на валиках, примыкающих либо к залитому, либо к незалитому чеку. Контрольные площадки выбирали на удалении не более 1 км от залитого чека (местообитание “рис”). Из-за особенностей конфигурации в пространстве данных типов местообитаний (в частности, относительной удаленности контрольных местообитаний и необязательного соседнего расположения залитых и незалитых чеков) не удалось реализовать классический блочный план, что наложило некоторые ограничения на используемые методы статистического анализа. Всего обследовано 42 площадки – по 14 в каждом регионе.
Таблица 1.
Численность (экз/м2) родов (выделено полужирным) и видов (не все определены до уровня вида) энхитреид в пределах различных типов местообитаний трех регионов (в скобках указано количество повторностей местообитаний) и разделение таксонов энхитреид на экологические группы по отношению к кислотности и влажности почвы (по: [5–8, 23–25]; ? – нет данных)
| Род/вид | pH-префе-ренция | Гидро-префе-ренция | Краснодар | Калмыкия | Приморье | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| контроль (n = 3) | рис (n = 3) | незаливные культуры (n = 3) |
валик (n = 5) | контроль (n = 4) | рис (n = 3) | незаливные культуры (n = 3) | валик (n = 4) |
контроль (n = 3) | рис (n = 3) | незаливные культуры (n=5) | валик (n=3) |
|||
| Enchytraeus Henle, 1837 | 4303 ± 2131 | 0 | 57 ± 57 | 1053 ± 490 | 807 ± 175 | 962 ± 962 | 3680 ± 2750 | 3142 ± 1559 | 510 ± 294 | 0 | 0 | 0 | ||
| E. buchholzi Vejdovský, 1879 | Нейтро-фил | Ксерофил | 934 ± 934 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| E. dichaetus Schmelz & Collado, 2010 | Базофил | ? | 1189 ± 1079 | 0 | 0 | 34 ± 34 | 0 | 962 ± 833 | 2437 ± 1306 | 1529 ± 763 | 170 ± 147 | 0 | 0 | 0 |
| Fridericia Michaelsen, 1889 | 849 ± 612 | 0 | 0 | 442 ± 219 | 0 | 0 | 0 | 425 ± 202 | 0 | 0 | 0 | 113 ± 113 | ||
| F. galba (Hoffmeister, 1843) | Базофил | Гигро-мезофил | 212 ± 127 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| F. singula Nielsen & Christensen, 1961 | Нейтрофил | ? | 297 ± 297 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| F. tuberosa Rota, 1995 | Нейтро-фил | Гигро-фил | 0 | 0 | 0 | 68 ± 68 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Henlea Michaelsen, 1889 | 0 | 0 | 170 ± 170 | 0 | 0 | 0 | 0 | 297 ± 297 | 1246 ± 1080 | 0 | 170 ± 170 | 962 ± 723 | ||
| H. jutlandica Nielsen & Christensen, 1959 | ? | Гигро-фил | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 297 ± 297 | 0 | 0 | 0 | 0 |
| H. perpusilla Friend, 1911 | Нейтро-фил | Мезо-фил | 0 | 0 | 113 ± 98 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 396 ± 343 |
| Mesenchytraeus sp. Eisen, 1878 | Ацидо-фил | Гигро-фил | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1982 ± 709 | 0 | 0 | 396 ± 150 |
| Enchytraedae gen sp. | 962 ± 623 | 0 | 283 ± 150 | 408 ± 175 | 0 | 283 ± 283 | 226 ± 150 | 2335 ± 1959 | 0 | 0 | 0 | 113 ± 113 | ||
| Общая численность | 6115 ± 3118 | 0 | 510 ± 98 | 1902 ± 625 | 807 ± 175 | 1246 ± 857 | 3907 ± 2894 | 6200 ± 1062 | 3737 ± 1092 | 0 | 170 ± 170 | 1585 ± 736 | ||
Почвенные монолиты (по три на площадке) для последующей экстракции энхитреид в пределах каждого региона были отобраны буром диаметром 5 см до глубины 8 см. Пробы на рисовых чеках были взяты из почвы, полностью покрытой водой. Одновременно с отбором проб измеряли мощность подстилки (при ее наличии). Непосредственно после отбора монолиты были помещены в термоконтейнеры с постоянной температурой +6°С и доставлены в лабораторию.
Экстракция энхитреид из почвы была проведена методом Рёмбке [22] не более чем через 10 дней после отбора проб. Сита с почвенными пробами стояли в емкостях с водой при температуре 15°С в течение двух дней, после чего осевший на дно материал собирали в пробирки. В каждой пробе было подсчитано общее количество энхитреид, далее животные были определены с точностью до рода, по возможности до вида, затем разделены на экологические группы по отношению к кислотности почвы: базофилы, нейтрофилы, ацидофилы [5–8, 23–25]. Виды, для которых отношение к кислотности неизвестно, и особи, не определенные с точностью до вида, сформировали дополнительную группу, которая не была включена в анализ. Разбиение на экологические группы по фактору предпочитаемой влажности почвы признано нецелесообразным, так как в целом повторяет таксономическое деление на роды (см. табл. 1). Все данные о численности энхитреид в данной работе приведены в виде “среднее ± стандартная ошибка”.
Для каждой почвенной пробы в лаборатории стандартными методами [26, 27] были определены: pH почвенного раствора (H2O), водоудерживающая способность почвы (ВУС, %), содержание (мг/100 г) аммонийного азота и фосфора, оценены потери после прокаливания (ППП, %). С помощью прибора PH-061 был определен pH почвенного раствора как pH водной суспензии при отношении почвы к воде 1 : 2.5. Содержание аммонийного азота было измерено согласно ГОСТ Р 53219-2008; содержание подвижного фосфора – фотоколориметрическим методом согласно ГОСТ Р 54650-2011; потери после прокаливания – при прокаливании почвы в фарфоровых тиглях при температуре 450°C в муфельной печи ПМ-8. Перед анализом почва была высушена при температуре 50°C и измельчена при помощи вибрационной мельницы Retsch MM200. Измерения проведены в Центре коллективного пользования “Инструментальные методы в экологии” при ИПЭЭ РАН.
Монолиты, отобранные в пределах каждой пробной площадки (n = 3), представляли собой мнимые повторности и были использованы только для вычисления среднего значения для каждой площадки (всего 42). Анализ остатков (residuals) для общей численности энхитреид и численности различных экологических групп не позволил подтвердить нормальность распределения данных. В связи с этим зависимость этого показателя от факторов “регион” (Краснодарский край, Калмыкия, Приморье) и “тип местообитания” (рис, незаливная культура, валик, контроль), а также их взаимодействия между собой была оценена с помощью обобщенных линейных моделей для данных с отрицательным биномиальным распределением (значения численности округлены до целых чисел), на которое было наиболее похоже распределение общей численности энхитреид (QQ-тест). Для оценки различий средней численности энхитреид между типами местообитаний был использован критерий Тьюки для планов с неравной повторностью. За уровень статистической значимости в анализе принято p < 0.05.
Анализ зависимости численности родов (4 рода) и экологических групп энхитреид от почвенных параметров был проведен методом главных компонент (PCA) для лог-трансформированных данных (ln(x + 1)). В качестве активных переменных выбрана численность родов энхитреид или их функциональных групп соответственно, а почвенные параметры (pH, ВУС, подвижный фосфор, аммонийный азот, ППП) – в качестве дополнительных (пассивных) переменных. Статистическая обработка данных выполнена с использованием программного обеспечения Statistica 13.3 (TIBCO Software Inc., Tulsa, США).
РЕЗУЛЬТАТЫ
В рисовых агроэкосистемах в Краснодарском крае и Калмыкии были обнаружены представители трех родов энхитреид, а в Приморье – четырех (см. табл. 1). Только в Краснодарском крае были найдены Fridericia galba, F. singula, F. tuberosa, исключительно в Калмыкии – Henlea jutlandica; видовая принадлежность особей, собранных в Приморье, требует дальнейшего уточнения.
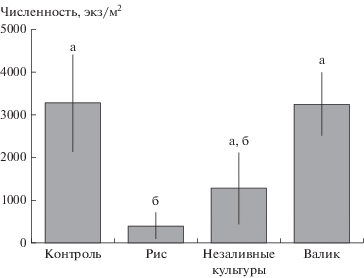
На общую численность энхитреид статистически значимо влияли фактор “тип местообитания”, а также взаимодействие факторов “регион” и “тип местообитания” (табл. 2). В Калмыкии наибольшая численность энхитреид отмечена на валиках и полях незаливных культур, в то время как в Краснодарском крае и Приморье – в контрольных местообитаниях (см. табл. 1). В целом наиболее близкие значения численности с контрольными участками обнаружены в пределах валиков, несколько меньше (недостоверно) численность была на полях незаливных культур и минимальная – на рисовых чеках (рис. 1). Численность энхитреид, отличная от нуля, в пределах чеков, засеянных рисом, обнаружена исключительно в Калмыкии (см. табл. 1). Наибольшая средняя численность энхитреид отмечена на площадках, расположенных в Калмыкии (3142 ± 1648 экз/м2; табл. 1), несколько меньше – в Краснодарском крае (2123 ± 1609 экз/м2), минимальная – в Приморье (1322 ± 955 экз/м2), но средние значения достоверно друг от друга не отличались (см. табл. 2).
Таблица 2.
Результаты статистического анализа зависимости общей численности энхитреид от факторов “регион”, “тип местообитания” и их взаимодействия методом обобщенных моделей для данных с отрицательным биномиальным распределением (н.з. – незначимо)
| Фактор | Статистика Вальда |
Уровень p |
|---|---|---|
| Регион | н.з. | |
| Тип местообитания | 20.83 | >0.0001 |
| Регион × тип местообитания | 53.90 | >0.0001 |
| Свободный член | н.з. |
Рис. 1.
Суммарная численность (экз/м2, среднее ± ошибка) всех собранных энхитреид в различных типах местообитаний. Столбцы, отмеченные разными буквами, статистически значимые различия средних (критерий Тьюки, p < 0.05) между типами местообитаний.
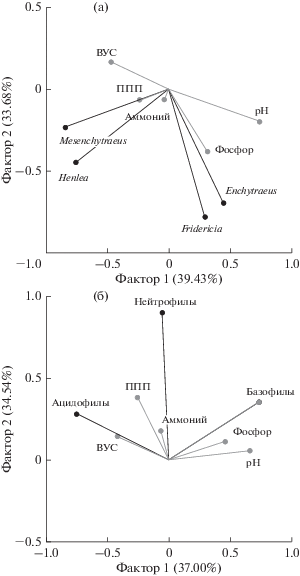
Анализ методом главных компонент (PCA) показал, что численность родов Henlea и Mesenchytraeus положительно коррелировала с величинами ППП, слабо положительно – с ВУС и отрицательно – с величиной pH, в то время как численность родов Fridericia и Enchytraeus имела существенную корреляционную связь с содержанием фосфора в почве и умеренно отрицательную – с ВУС (рис. 2а). Численность энхитреид базофильной экологической группы (E. dichaetus, F. galba) положительно коррелировала со значением pH и содержанием фосфора в почве, а численность энхитреид ацидофильной экологической группы (Mesenchytraeus) – отрицательно с теми же параметрами и положительно – с ВУС (рис. 2б). При этом умеренная положительная корреляционная связь с содержанием аммония в почве была отмечена для численности нейтрофильных энхитреид (E. buchholzi, F. singula, F. tuberosa, H. perpusilla).
Рис. 2.
Результаты анализа методом главных компонент (для лог-трансформированных данных), показывающие отношения между численностью разных родов энхитреид (a) и различных экологических групп энхитреид (б) и некоторыми почвенно-эдафическими характеристиками: черные линии – основные (активные) переменные анализа, серые – вспомогательные (пассивные) переменные; ВУС – водоудерживающая способность почвы (%); pH – pH почвенного раствора (H2O); аммоний – содержание аммонийного азота (мг/100 г); фосфор – содержание подвижного фосфора (мг/100 г); ППП – потери после прокаливания (%).
ОБСУЖДЕНИЕ
Наши результаты свидетельствуют от том, что в отличие от тропиков [1] численность энхитреид на залитых рисовых чеках умеренных широт относительно невысока. Известно, что энхитреиды предпочитают более влажные местообитания [28, 29], но в условиях умеренных широт в России они избегают залитых чеков, демонстрируя высокую численность на оптимально увлажненных валиках с более богатой и густой растительностью, стабилизирующей уровень влажности почвы. Это идет вразрез с данными для аналогичных местообитаний на Филиппинах [1], показывающими, что в условиях тропиков энхитреиды – одна из доминирующих групп в сообществах залитых чеков. После дренирования чеков численность энхитреид под незаливными культурами быстро восстанавливается до уровня, сопоставимого с контролем и/или валиком вне зависимости от региона. Скорее всего, выявленные закономерности объясняются особенностями севооборота риса в России. В наших условиях рисовые чеки находятся под водой незначительное время, но при этом уровень воды нередко превышает 50 см (собственные неопубликованные наблюдения), тогда как в тропиках глубина воды на рисовых чеках колеблется около 5 см [1]. Кроме того, валики в тропиках характеризуются высокой температурой поверхности почвы, которая не оптимальна для данной группы [6]. В условиях умеренных широт условия на валиках более оптимальны.
Относительно низкая (по сравнению с местообитаниями других типов) численность энхитреид на контрольных площадках в Калмыкии в отличие от других регионов может быть связана с повышенным содержанием солей (до 0.5%) и засушливостью климата в целом [30], хотя об отношении к засолению рода Enchytraeus, доминировавшего в этом регионе (см. табл. 1), и представителей других родов, не найденных здесь, известно мало [5]. Также нельзя исключать, что относительно низкая численность энхитреид даже в контроле в Калмыкии может быть связана с небольшой мощностью органогенного горизонта, определяемой типом почв, и интенсивным выпасом скота. Отсутствие достоверного влияния даты отбора проб в разных регионах также свидетельствует о том, что сезонная динамика не оказала значимого влияния на население энхитреид в нашем исследовании, и поэтому этими предварительными данными можно пренебречь.
В отличие от соотношения численности различных родов энхитреид, имеющих четкие предпочтения по влажности почвы (см. табл. 1), которые в значительной степени определялись ВУС, соотношение функциональных групп в большей степени зависит от локальной неоднородности прочих эдафических факторов, включенных в наш анализ. Последнее подтверждается наличием корреляции между численностью энхитреид-базофилов и ацидофилов и pH почв.
В заключение отметим, что, несмотря на низкую численность в почвах рисовых чеков, энхитреиды могут оставаться важным и крайне динамичным компонентом детритных пищевых сетей при интенсивном рисоводстве, достаточно быстро восстанавливающими численность под незаливными культурами. При этом они чутко реагируют не только на стадии севооборота, но и на изменения физико-химических условий почв.
Авторы признательны Р.А. Сайфутдинову и А.Ю. Горбуновой (ИПЭЭ РАН) за помощь в сборе и обработке материала; В.А. Ладатко (ВНИИ риса), С.Н. Чимидову (ФГУП “Харада”), Э.Б. Дедовой (Калмыцкий филиал ВНИИГиМ им. А.Н. Костякова) и Т.В. Суницкой (Приморский НИИСХ) – за содействие в организации полевых работ. Выражаем благодарность доктору К. Экшмитту (университет Юстуса Либиха, г. Гиссен, ФРГ) за консультацию при выборе адекватной модели для статистического анализа данных. Работа выполнена при финансовой поддержке гранта Российского научного фонда № 16-14-00096.
Список литературы
Schmidt A., John K., Arida G. et al. Effects of residue management on decomposition in irrigated rice fields are not related to changes in the decomposer community // PLoS ONE. 2015. V. 10. Article e0134402. https://doi.org/https://doi.org/10.1371/journal.pone.0134402
Smil V. Crop residues: Agriculture’s largest harvest: Crop residues incorporate more than half of the world’s agricultural phytomass // BioScience. 1999. V. 49. P. 299–308. https://doi.org/https://doi.org/10.2307/1313613
Lagerlöf J., Andrén O. Succession and activity of microarthropods and enchytraeids during barley straw decomposition // Pedobiologia. 1985. V. 28. P. 343–357.
Jänsch S., Rӧmbke J., Didden W.A.M. The use of earthworms in ecological soil classification and assessment concepts // Ecotoxicology and Environmental Safety. 2005. V. 62. P. 249–265. https://doi.org/https://doi.org/10.1016/j.ecoenv.2005.03.027
Römbke J., Beck L., Dreher P. et al. Entwicklung von bodenbiologischen Bodengüteklassen für Acker- und Grünlandstandorte / Umweltforschungsplan des Bundesministeriums für Umwelt, Naturschutz und Reaktorsicherheit, Forschungsbericht / UBA-Texte, 2002. 264 S.
Schmelz R.M., Collado R. A guide to European terrestrial and freshwater species of Enchytraeidae (Oligochaeta) // Soil Organisms. 2010. V. 82. P. 1–176
Jänsch S., Römbke J. Ökologische Charakterisierung ausgewählter Enchytraeenarten hinsichtlich relevanter Bodeneigenschaften // UWSF. 2003. V. 15. P. 95–105. https://doi.org/https://doi.org/10.1065/uwsf2002.04.020
Didden W.A.M., Fründ H., Graefe U. Enchytraeids // Fauna in soil ecosystems: recycling processes, nutrient fluxes, and agricultural production. New York: Marcel Dekker Inc., 1997. P. 135–172.
Залесская Н.Т. Фауна энхитреид (Oligochaeta, Enchytraeidae) Московской области // Почвенные беспозвоночные Московской области / Под ред. Гилярова М.С. М.: Наука, 1982. С. 119–133.
Nurminen M. The Enchytraeidae – open field for research in the USSR // Pedobiologia. 1982. V. 23. P. 233.
Бызова Ю.Б., Уваров А.В., Губина В.Г. и др. Почвенные беспозвоночные беломорских островов Кандалакшского заповедника. М.: Наука, 1986. 309 с.
Karnatak A.K., Karnataka D.C., Shukla A., Firake D.M. Impact of pesticides on earthworms and enchytraeids and their bio-diversity in rice-wheat cropping system in Indo-Gangetic plains // Himalayan J. of Environment and Zoology. 2007. V. 21. 29 p.
Schmidt A., John K., Auge H. et al. Compensatory mechanisms of litter decomposition under alternating moisture regimes in tropical rice fields // Applied Soil Ecology. 2016. V. 107. P. 79–90. https://doi.org/https://doi.org/10.1016/j.apsoil.2016.05.014
Simpson I.C., Roger P.A., Oficial R., Grant I.F. Density and composition of aquatic oligochaete populations in different farmers' ricefields // Biol. Fert. Soils. 1993. V. 16. P. 34–40.
Simpson I.C., Roger P.A., Oficial R., Grant I.F. Impacts of agricultural practices on aquatic oligochaete populations in ricefields // Biol. Fert. Soils. 1993. V. 16. P. 27–33. http://dx.doi.org/https://doi.org/10.1007/BF00336511
Bambaradeniya C.N.B., Amarasinghe F.P. Biodiversity Associated with the rice field agroecosystem in Asian countries: A brief review. Colombo: International Water Management Institute, IWMI Working Paper 063, 2003. 24 p. http://dx.doi.org/https://doi.org/10.3910/2009.193
Coyle D.R., Nagendra U.J., Taylor M.K. et al. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action // Soil Biol. Biochem. 2017. V. 110. P. 116–133. https://doi.org/https://doi.org/10.1016/j.soilbio.2017.03.008
Russian Hydrometeorological Research Center public online database. https://meteoinfo.ru.
Peel M.C., Finlayson B.L., McMahon T.A. Updated world map of the Köppen-Geiger climate classification // Hydrol. Earth Syst. Sci. 2007. V. 11. P. 1633–1644. https://doi.org/https://doi.org/10.5194/hess-11-1633-2007
Полевой определитель почв. М.: Почвенный ин-т им. В.В. Докучаева, 2008. 182 с.
IUSS Working Group WRB. World reference base for soil resources 2014. World Soil Resources Reports. 2015. V. 106.
Römbke J. Enchytraeen (Oligochaeta) als Bioindikator // UWSF. 1995. V. 7. P. 246–249.
Rota E. Enchytraeidae (Oligochaeta) of western Anatolia: taxonomy and faunistics // Bollettino di Zoologia. 1994. V. 61. P. 241–260.
Rota E. Italian Enchytraeidae (Oligochaeta) // Bollettino di Zoologia. 1995. V. 62. P. 183–231.
Global Biodiversity Information Facility (GBIF). www.gbif.org.
Методы исследования структуры, функционирования и разнообразия детритных пищевых сетей. Методическое руководство / Под. ред. Покаржевского А.Д., Гонгальского К.Б., Зайцева А.С. М.: ИПЭЭ РАН. 2003. 100 с. [Investigation methods of structure, functioning and diversity of detrital food-webs. Eds. Pokarzhevskii A.D., Gongalsky K.B., Zaitsev A.S. Moscow: IEE RAS, 2003. 100 p.]
Гонгальский К.Б., Зайцев А.С., Коробушкин Д.И. и др. Разнообразие почвенной биоты на гарях южнотаежных лесов (на примере Тверской области) // Почвоведение. 2016. № 3. С. 388–397. [Gongalsky K.B., Zaitsev A.S., Korobushkin D.I. et al. Diversity of the Soil Biota in Burned Areas of Southern Taiga Forests (Tver Oblast) // Eurasian Soil Science. 2016. V. 49. P. 358–366.] https://doi.org/https://doi.org/10.1134/S1064229316030042
O’Connor F.B. The enchytraeids / Methods of study in quantitative soil ecology / Philipson J. (ed.). Edinburgh: Blackwell, 1971. P. 83–106.
Dash M.C. Oligochaeta: Enchytraeidae / Dindal D.L. (ed.). Soil Biology Guide. NY: Wiley and Sons, 1990. P. 311–340.
Национальный атлас почв Российской Федерации. Под ред. Шобы С.А. М.: Астрель, 2011. 632 с.
Дополнительные материалы отсутствуют.