

Экология, 2019, № 4, стр. 286-296
Соотношение хронографической и географической изменчивости морфофункциональных признаков рыжей полевки (Clethrionomys glareolus Shreb.) на юге ареала
А. Г. Васильев a, *, В. Н. Большаков a, И. А. Васильева a, Ю. В. Городилова a, М. В. Чибиряк a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: vag@ipae.uran.ru
Поступила в редакцию 10.10.2018
После доработки 12.11.2018
Принята к публикации 30.11.2018
Аннотация
Параллельный анализ хронографической изменчивости нижней челюсти рыжей полевки (Clethrionomys glareolus Shreb.) в трех географически удаленных в долготном направлении популяциях, населяющих пойменные леса рек Сакмары и Самары на юге ареала вида в Оренбургской области, выявил в каждой из них существенные морфогенетические и морфофункциональные изменения за 33-летний период. Методами геометрической морфометрии обнаружены изменения формы нижней челюсти в аллохронных выборках полевок (1983 и 2015 гг.). Морфофункциональные перестройки оценили по мандибулярным индексам, характеризующим различия в строении нижней челюсти, вызванные сменой функциональных нагрузок при смене диеты. Во всех аллохронных популяциях произошли значимые изменения как формы мандибул, так и величин индексов. Хронографическая изменчивость формы нижней челюсти для всех популяций имеет сходное направление в морфопространстве, по размаху превышая географическую изменчивость и отражая существенную историческую перестройку морфогенеза. Обсуждается вероятная связь быстрых морфогенетических и морфофункциональных перестроек мандибул полевок с трансформацией локальных трофических условий, обусловленных разной направленностью растительных сукцессий, связанных в свою очередь с локальными трендами климатических изменений.
Проблема соотношения хронографической и географической изменчивости при оценке исторических процессов в популяциях животных издавна привлекает внимание экологов и эволюционистов [1–10]. Обычно принято считать, что размах географических различий между популяциями выражен больше, чем хронографических изменений в каждой из них. При этом, однако, оценка популяционных изменений во времени часто охватывает не более трех последовательных лет сравнения. Анализ хронографической изменчивости на больших исторических отрезках времени, как правило, не проводится. Оценка соотношения географической и хронографической форм изменчивости на основе параллельных сборов синтопных аллохронных выборок за исторически длительный отрезок времени представляет особый эволюционно-экологический интерес, поскольку прямой или опосредованный морфогенетический отклик на влияние климатических изменений за исторические времена продолжительностью более двух–трех десятилетий более вероятно обнаружить, чем за два–три смежных года. Климатические тренды вызывают ускорение сукцессионных процессов, влияют на их направленность [11], что в свою очередь в виде смены диеты оказывает воздействие на морфофункциональные признаки консументов первого порядка [12]. Так, смена состава растительности может, например, повлиять на строение функциональных структур грызунов – форму и размеры нижней челюсти, щечных зубов, прогнатизм резцов, а также пропорции частей осевого черепа [13–15]. В последние годы появляются примеры направленных изменений конфигурации нижней челюсти грызунов при питании разной по механическому составу пищей [12, 14, 16]. Параллельный долговременный хроно-географический анализ изменчивости позволяет на основе методов геометрической морфометрии [15, 17–19] оценить отдаленные морфогенетические и морфофункциональные перестройки модельных морфоструктур.
Цель данной работы заключалась в сопоставлении размаха и направления хронографической и географической форм изменчивости морфофункциональных признаков в одних и тех же удаленных популяциях рыжей полевки на южной границе ареала за исторический период, включающий более 30 лет (1983–2015 гг.). Особое внимание уделено эволюционно-экологическому анализу быстрых морфогенетических и морфофункциональных исторических перестроек нижней челюсти двух внутрипопуляционных структурно-функциональных групп: самцов и самок возрастной группы взрослых сеголеток.
МАТЕРИАЛ И МЕТОДЫ
В качестве материала использовали коллекции Зоологического музея ИЭРиЖ УрО РАН, собранные авторами в 1983 и 2015 гг. в трех локалитетах Оренбургской области: 1 – с. Тоцкое (52°31′54″ с.ш., 52°47′16″ в.д.), 2 – с. Платовка (51°56′09″ с.ш., 54°00′45″ в.д.) и 3 – г. Кувандык (51°28′50″ с.ш., 57°28′25″ в.д.). Сбор материала в XX и XXI вв. проведен во второй половине лета в сходных по условиям биотопах – пойменных лесах рек Самара (тоцкая и платовская популяции) и Сакмара (сакмарская популяция). Локалитеты размещены в пространстве в линию с запада на восток: крайними являются тоцкая и сакмарская популяции, платовская занимает промежуточное положение, но располагается географически ближе к тоцкой. Отловы осуществлены с помощью линий деревянных ловушек-плашек в течение 5 сут экспозиции. Сравнивали синтопные выборки взрослых сеголеток рыжей полевки 1983 и 2015 гг. Общий объем изученного материала по рыжей полевке в 1983 г. составил 97 экз., а в 2015 г. – 103 экз. (тоцкая – 35 и 39, платовская – 35 и 23, сакмарская – 27 и 41 соответственно). Исследование провели на выборках взрослых сеголеток обоих полов.
В качестве объекта исследования использовали нижнюю челюсть как важный в экологическом отношении орган, непосредственно связанный с кормодобыванием, функцией питания и ценотической ролью вида. Изменчивость формы нижней челюсти изучали методами геометрической морфометрии [15, 17–19] на сканированных изображениях правых нижнечелюстных ветвей. Оцифровку фотографий лингвальной стороны мандибулы провели при разрешении 1200 dpi с помощью программ tpsUtil и tpsDig2 [20, 21]. Для характеристики изменчивости формы нижней челюсти использовали конфигурацию 16 гомологичных меток-ландмарок (landmarks) (рис. 1). Мандибулярные размеры в выборках полевок оценили по величине площади нижнечелюстной ветви (SM), вычисленной в пределах ее контура, замкнутого по меткам-ландмаркам, с помощью программы tpsUtil [20], поскольку обычно используемый для косвенной оценки размеров центроидный размер (CS – centroid size) зависит не только от величины объекта, но и от степени “изрезанности” его контура [18]. Суперимпозиция конфигураций ландмарок выполнена методом генерализованного Прокрустова анализа – GPA [22] с использованием наименьших квадратов и вычислением прокрустовых координат (Procrustes coordinates), характеризующих изменчивость формы (shape). Методы геометрической морфометрии позволяют изучать изменчивость формы оцифрованных объектов, исключив влияние изменчивости их размеров, и допускают возможность морфогенетической интерпретации различий [15, 18, 19].
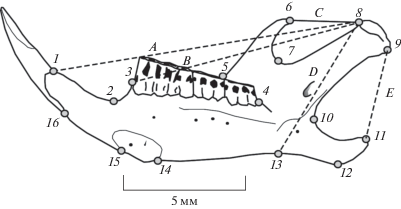
Рис. 1.
Схема размещения на лингвальной стороне нижнечелюстной ветви рыжей полевки 16 меток-ландмарок (1–16) и промеров (A–E), необходимых для вычисления мандибулярных функциональных индексов: TI, TM, MI, MM, AM (формулы для их расчета см. в тексте).
Межгрупповые различия формы нижней челюсти оценивали с помощью канонического анализа прокрустовых координат. Для оценки значимости межгрупповых различий по факторам год (Y) и локалитет (L) по совокупности значений всех канонических переменных применили непараметрический многомерный двухфакторный анализ (Two-way NPMANOVA), основанный на перестановочном (permutation) тестировании для 10 000 повторных реплик [23].
Для оценки функциональных особенностей конфигураций нижней челюсти использовали пять промеров: A – сочленовно-резцовый, B – сочленовно-молярный, C – темпорально-сочленовный, D – сочленовно-массетерный и E – сочленовно-угловой (см. рис. 1). По данным промерам вычислили функциональные мандибулярные индексы [см. 12]: TI – темпорально-резцовый, TM – темпорально-молярный, MI – массетерно-резцовый, MM – массетерно-молярный и AM – ангулярно-массетерный. Расчет индексов провели по формулам [12]: TI = C/A; TM = C/B; MI = D/A; MM = D/B и AM = E/D. Индексы TI и TM характеризуют интенсивность продольных режуще-жевательных движений, а индексы MI и MM – интенсивность движений, связанных с грызением и дроблением пищевых объектов. Предложенный нами индекс AM косвенно характеризует интенсивность поперечных жевательных движений, связанных с перетиранием корма [24]. При множественном сравнении использовали однофакторный (One-way ANOVA) метод дисперсионного анализа. Межвыборочные парные различия оценили с помощью апостериорного теста по Q-критерию Тьюки.
Уровни внутригруппового морфофункционального разнообразия оценили методом анализа паттерна дистанций между ординатами средних значений индексов в пределах годового полигона изменчивости, для чего вычислили средние дистанции между ординатами (MMFD – measure of morphofunctional disparity) с учетом их стандартных ошибок [25]. Значимость различий MMFD между выборками оценили по критерию t для неравных дисперсий. Основные расчеты и статистический анализ материала выполнены с помощью программ TPS [20, 21], PAST 2.17с [26] и MorphoJ 1.6d [19].
РЕЗУЛЬТАТЫ
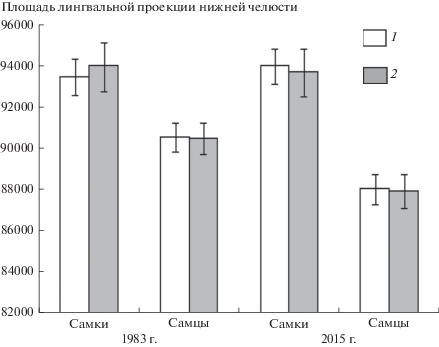
Сравнение площади лингвальной проекции нижнечелюстных ветвей (SM) между выборками самцов и самок выявило сходные половые различия в 1983 и 2015 гг. (рис. 2). Сравнение проводили как по реальным размерам выборок, так и по случайно выровненным по минимальным размерам (после процедуры рарефакции). Результаты в обоих случаях оказались почти идентичными. Статистическую значимость различий средней площади нижнечелюстных ветвей между выборками оценили с помощью апостериорного теста по Q-критерию Тьюки. В оба года наблюдений средние размеры нижней челюсти самок (и соответственно их габитуальные размеры) были значимо больше, чем у самцов (1983 г. – Q = 3.66, p = = 0.041; 2015 г. – Q = 7.47, p ⪡ 0.0001). Другими словами, на обоих временных этапах в популяциях наблюдается проявление полового диморфизма по размерам мандибул (1983 г. – 3.2%; 2015 г. – 6.6%). На основе однофакторного дисперсионного анализа (One-Way ANOVA) выявлены значимые различия (F = 12.21; d.f.1 = 3; d.f.2 = 196; p ⪡ 0.0001) по величине SM. Тест F Уэлча (Welch F-test) подтвердил этот результат (F = 12.16; d.f. = 108.5; p ⪡ 0.0001). Показатель размера эффекта Коэна (Cohen’s effect size) ƒ2 = 0.17 превысил в данном случае установленный средний уровень [27, 28], т.е. выявленные половые различия по размерам существенны.
Рис. 2.
Сравнение площади лингвальной проекции мандибулы самцов и самок рыжей полевки с учетом стандартных ошибок (±SE) в объединенных выборках 1983 и 2015 гг. по реальным объемам наблюдений (1) и минимальным случайным выборкам после процедуры рарефакции (2) (rarefaction).
При сравнении площади мандибул в объединенных по полу выборках трех синтопных географически удаленных популяций в разные годы также обнаружены значимые различия (One-Way ANOVA: F = 3.28; d.f.1 = 5; d.f.2 = 194; p = 0.007), но размер эффекта Коэна в данном случае крайне мал (ƒ2 = 0.06) и близок к наименьшему установленному уровню. Значимые различия обнаружены только в 2015 г. при сравнении географически наиболее удаленных тоцкой и сакмарской выборок (Q = 4.16; p = 0.038). Географические различия по размерам мандибул в пределах каждого временного этапа не выявлены и при дальнейшем сравнении формы ими можно пренебречь.
В результате канонического анализа прокрустовых координат, характеризующих изменчивость формы, установлено, что конфигурация нижней челюсти синтопных популяций в 1983 и 2015 гг. существенно различается (рис. 3, табл. 1). Межгрупповые различия вдоль всех канонических переменных статистически значимы. Вдоль первых трех осей проявилось 81.6% дисперсии, что позволяет достаточно полно характеризовать межгрупповую изменчивость формы мандибул. На рис. 3, где вдоль каждой канонической оси представлены центроиды выборок с отложенными величинами стандартных ошибок (±SE), видно, что межгрупповые различия между выборками разных временных этапов (хронографическая изменчивость) выражены вдоль первой канонической оси – CV1 (49.01% общей дисперсии). Полигон, включающий выборки 2015 г., смещен на графике вправо по отношению к полигону, объединяющему выборки 1983 г. По конфигурациям контурных изображений (outlines) мандибул вдоль первой оси при сопоставлении их с консенсусной конфигурацией, расположенной в центре рисунка, можно заключить, что от первого временного этапа (1983 г.) к современности (2015 г.) наблюдается усиление грацильности нижней челюсти, сопровождающееся удлинением углового отростка, редукцией головки сочленовного отростка с ее смещением кзади и укорочением венечного отростка.
Рис. 3.
Результаты канонического анализа прокрустовых координат, характеризующих межгрупповую хронографическую и географическую изменчивость формы нижней челюсти рыжей полевки трех аллохронных (83 – 1983 г.; 15 – 2015 г.) синтопных популяций (T – тоцкая, P – платовская, K – сакмарская). Центроиды выборок, приведенные с учетом стандартных ошибок, по каждому году объединены в полигоны изменчивости. Контурные изображения мандибул (outlines) соответствуют крайним значениям соответствующих канонических осей. В центре – консенсусная конфигурация. Стрелки указывают направления хронографической и географической изменчивости в общем морфопространстве.
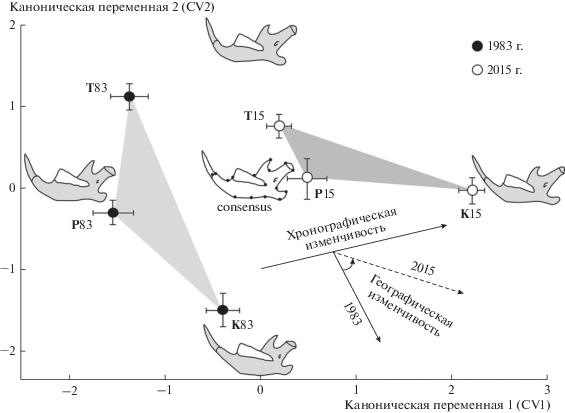
Таблица 1.
Результаты канонического анализа прокрустовых координат нижней челюсти и значения центроидов трех синтопных популяций рыжей полевки с учетом стандартных ошибок (±SE) в 1983 и 2015 гг.
| Названия выборок и оценка значимости межгрупповых различий | Канонические переменные | ||
|---|---|---|---|
| CV1 | CV2 | CV3 | |
| Тоцкая. 1983 г. | –1.373 ± 0.194 | 1.113 ± 0.159 | 0.690 ± 0.190 |
| Платовская. 1983 г. | –1.541 ± 0.213 | –0.297 ± 0.146 | –1.084 ± 0.146 |
| Сакмарская. 1983 г. | –0.396 ± 0.172 | –1.487 ± 0.203 | 0.598 ± 0.164 |
| Тоцкая. 2015 г. | 0.191 ± 0.134 | 0.754 ± 0.143 | –0.107 ± 0.147 |
| Платовская. 2015 г. | 0.482 ± 0.206 | 0.115 ± 0.248 | 0.754 ± 0.271 |
| Сакмарская. 2015 г. | 2.204 ± 0.136 | –0.030 ± 0.158 | –0.374 ± 0.162 |
| Собственные числа | 1.731 | 0.709 | 0.441 |
| Λ-критерий Уилкса | 0.0847 | 0.2314 | 0.3955 |
| Число степеней свободы | 140 | 108 | 78 |
| Доля дисперсии, % | 49.01 | 20.09 | 12.48 |
| Уровень значимости, p | <0.0001 | <0.0001 | <0.0001 |
Вдоль второй канонической оси CV2 (20.09% дисперсии) проявилась географическая изменчивость популяционных выборок в направлении с запада на восток. Наиболее четко географическая изменчивость формы нижней челюсти выражена вдоль данной оси между выборками 1983 г. (от наиболее западной тоцкой до крайней восточной сакмарской) и проявляется в относительном укорочении углового отростка, удлинении венечного отростка и смещении головки сочленовного отростка кпереди при общем увеличении высоты тела мандибулы и относительной длины нижнего зубного ряда. В выборках 2015 г. направление географической изменчивости формы нижней челюсти в общем морфопространстве изменилось и в значительной степени стало сходным с направлением хронографической изменчивости.
С помощью многомерного непараметрического двухфакторного дисперсионного анализа PERMANOVA по комплексу всех пяти канонических переменных оценили значимость влияния факторов год (Y), локалитет (L) и их взаимодействия (Y × L) на межгрупповую изменчивость формы нижней челюсти в аллохронных синтопных популяциях рыжей полевки (табл. 2). Влияние обоих факторов и их взаимодействия оказалось высоко значимым (p < 0.0001). Аналогичный двухфакторный PERMANOVA с учетом факторов год (Y) и пол (S) дал сходный результат, однако уровень значимости влияния фактора пол был ниже (p = 0.024). При этом взаимодействие факторов год (Y) и пол (S) статистически недостоверно (p = 0.122).
Таблица 2.
Результаты многомерного непараметрического двухфакторного дисперсионного анализа PERMANOVA канонических переменных, характеризующих изменчивость формы нижней челюсти в трех аллохронных (1983 и 2015 гг.) географически удаленных синтопных популяциях рыжей полевки в Оренбургской области
| Источник изменчивости (фактор) | Сумма квадратов | d.f. | Средний квадрат | F | Уровень значимости (p) |
|---|---|---|---|---|---|
| Год (Y) | 261.40 | 1 | 261.400 | 49.762 | 0.0001 |
| Локалитет (L) | 264.01 | 2 | 132.000 | 25.129 | 0.0001 |
| Взаимодействие (Y × L) | 121.99 | 2 | 60.994 | 11.611 | 0.0001 |
| Остаточная (Residual) | 1019.10 | 194 | 5.253 | ||
| Общая (Total) | 1666.50 | 199 |
Изменения конфигурации мандибул неизбежно связаны с модификацией их функциональной нагрузки и отражены в величинах мандибулярных индексов. Предварительно оценили средние значения индексов в сравниваемых популяциях на объединенных по полу выборках (табл. 3). Попарное сопоставление значений индексов между годами и в пределах каждого года между локалитетами выявило несколько значимых различий. По темпорально-резцовому индексу (TI) во всех трех популяциях наблюдаются значимые межгодовые (хронографические) изменения, однако если в тоцкой популяции от 1983 к 2015 г. происходит уменьшение индекса, т.е. ослабление жевательной функции, то в платовской и сакмарской, напротив, наблюдается увеличение индекса и, следовательно, усиление данной функции. В сакмарской популяции также значимо возрастает ангулярно-массетерный (AM) индекс, характеризующий интенсивность поперечных перетирающих жевательных движений мандибулы. В платовской популяции от 1983 к 2015 г. значимо возрастают массетерно-резцовый (MI) и массетерно-молярный (MM) индексы, т.е. усиливается степень грызущих и дробящих пищу движений нижней челюсти на фоне общего усиления ксероморфности растительности в локалитете. Судя по величинам индексов, в разных популяциях как в пределах 1983 г., так и в 2015 г. межпопуляционные различия также хорошо выражены. Интересно, что индексы, характеризующие жевательную функцию (TI – темпорально-резцовый и TM – темпорально-молярный), в географически наиболее удаленных популяциях – тоцкой и сакмарской – в разные годы изменяются противоположно: в 1983 г. в тоцкой популяции индексы значимо выше, чем в сакмарской, а в 2015 г., наоборот, существенно ниже. Другими словами, у зверьков этих популяций наблюдаются встречные, противоположно направленные процессы функциональных изменений в строении и инструментальном использовании мандибул.
Таблица 3.
Мандибулярные индексы (±стандартные ошибки, SE), характеризующие морфофункциональные свойства нижней челюсти трех синтопных аллохронных популяций рыжей полевки на двух отдаленных временных этапах (1983 и 2015 гг.) в Оренбургской области (данные объединены по полу)
| Популяция, год | N, экз. | Морфофункциональные мандибулярные индексы (±SE) | ||||
|---|---|---|---|---|---|---|
| TI | TM | MI | MM | AM | ||
| 1983 г.: | ||||||
| Тоцкая | 27 | 0.233 ± 0.004 | 0.299 ± 0.006 | 0.549 ± 0.004 | 0.704 ± 0.005 | 0.591 ± 0.005 |
| Платовская | 35 | 0.207 ± 0.004 | 0.267 ± 0.005 | 0.530 ± 0.003 | 0.684 ± 0.004 | 0.590 ± 0.005 |
| Сакмарская | 35 | 0.213 ± 0.004 | 0.274 ±0.006 | 0.538 ± 0.003 | 0.691 ± 0.004 | 0.583 ± 0.005 |
| 2015 г.: | ||||||
| Тоцкая | 41 | 0.219 ± 0.003 | 0.281 ± 0.003 | 0.543 ± 0.002 | 0.696 ± 0.003 | 0.592 ± 0.005 |
| Платовская | 23 | 0.222 ± 0.006 | 0.284 ± 0.007 | 0.545 ± 0.003 | 0.694 ± 0.004 | 0.587 ± 0.004 |
| Сакмарская | 39 | 0.232 ± 0.003 | 0.296 ± 0.004 | 0.537 ± 0.003 | 0.686 ± 0.003 | 0.603 ± 0.005 |
Многомерный двухфакторный непараметрический анализ (NPMANOVA) значений мандибулярных индексов (табл. 4) выявил значимое влияние факторов принадлежности к году (Y), локалитету (L) и их взаимодействия (Y × L). В этом случае наибольшая межгрупповая дисперсия обнаружена между популяциями, а наименьшая связана с межгодовыми различиями индексов. Промежуточный уровень величины дисперсии наблюдался при взаимодействии факторов, т.е. в разные годы в разных популяциях могут происходить существенные, иногда противоположные, изменения значений мандибулярных индексов.
Таблица 4.
Результаты многомерного непараметрического двухфакторного дисперсионного анализа PERMANOVA значений мандибулярных индексов, характеризующих функциональные особенности нижней челюсти трех аллохронных (1983 и 2015 гг.) географически удаленных синтопных популяций рыжей полевки в Оренбургской области
| Источник изменчивости (фактор) | Сумма квадратов | d.f. | Средний квадрат | F | Уровень значимости (p) |
|---|---|---|---|---|---|
| Год (Y) | 0.00980 | 1 | 0.00980 | 3.354 | 0.0191 |
| Локалитет (L) | 0.02108 | 2 | 0.01054 | 3.608 | 0.0022 |
| Взаимодействие (Y × L) | 0.01723 | 2 | 0.00862 | 2.950 | 0.0001 |
| Остаточная (Residual) | 0.56662 | 194 | 0.00292 | ||
| Общая (Total) | 0.61473 | 199 |
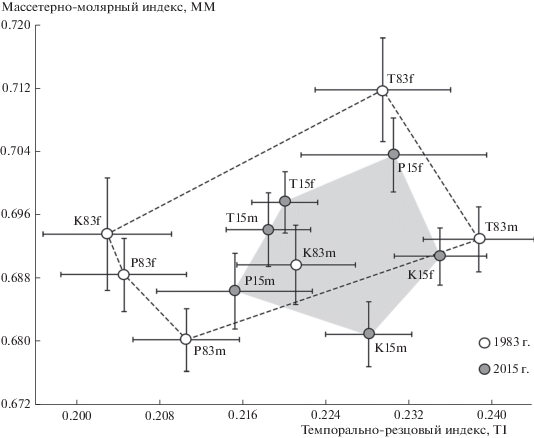
Сравнение значений индексов провели также с учетом пола зверьков. На рис. 4 по индексам TM и MI оконтурены полигоны межгрупповой изменчивости, соответствующие двум временным этапам, объединяющие центроиды выборок самцов и самок с учетом стандартных ошибок для каждого года. Площадь полигона изменчивости для выборок 2015 г. оказалась существенно меньше по сравнению с полигоном 1983 г., отражая относительно большее морфофункциональное сходство географически удаленных популяций рыжей полевки в начале XXI в. Такое же уменьшение площади полигона изменчивости для выборок 2015 г. наблюдается в общем морфопространстве и по индексам TI и MM (рис. 5).
Рис. 4.
Сравнение средних значений морфофункциональных мандибулярных индексов MI и TM с учетом стандартных ошибок (±SE) в выборках самцов (m) и самок (f) тоцкой (T), платовской (P) и сакмарской (K) популяций рыжей полевки разных лет (83 – 1983 г. и 15 – 2015 г.).
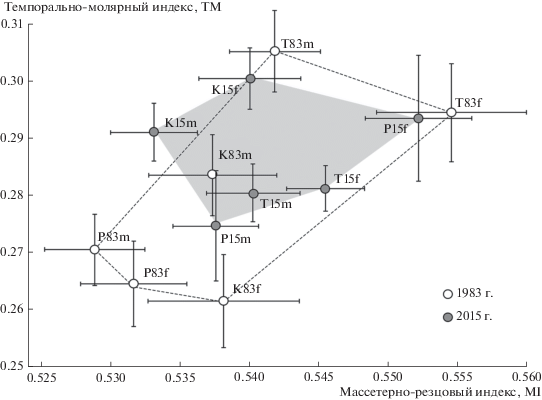
Рис. 5.
Сравнение средних значений морфофункциональных мандибулярных индексов TI и MM с учетом стандартных ошибок (±SE) в выборках самцов (m) и самок (f) тоцкой (T), платовской (P) и сакмарской (K) популяций рыжей полевки разных лет (83 – 1983 г. и 15 – 2015 г.).
Уровни внутригруппового морфофункционального разнообразия (MMFD) для каждого года оценили методом анализа паттерна евклидовых дистанций между ординатами выборок самцов и самок по средним значениям всех пяти функциональных мандибулярных индексов. Среднее значение MMFD для выборок 1983 г. составило 0.0322 ± ± 0.0017, а для 2015 г. – 0.0274 ± 0.0012. Полученные данные подтверждают, что в целом морфофункциональное разнообразие самцов и самок в трех модельных популяциях в 2015 г. значимо уменьшилось (для неравных дисперсий критерий t = 2.25; d.f. = 130; p = 0.0264) по сравнению с предыдущим временным этапом. Дисперсия межвыборочных дистанций в 1983 г. была также достоверно выше, чем в 2015 г. (F = 1.77; p = 0.0225).
ОБСУЖДЕНИЕ
Следует отметить, что в 1983 и 2015 гг. среднегодовые показатели как температуры, так и осадков были близки. Для этих лет также наблюдались значимые коэффициенты корреляции Пирсона между рядами среднемесячных температур (r = 0.99; p < 0.0001) и количества осадков (r = = 0.98; p < 0.0001). Поэтому наблюдаемые в эти годы эффекты не являются прямой реакцией на конкретные годовые условия, а во многом отражают исторически возникшие различия. По многолетним данным Оренбургской метеостанции, взятым начиная с 1966 г., можно заключить, что между первыми 25 годами (до 1990 г.) и последующими 25 (до 2015 г.) наблюдаются значимые различия по среднегодовым показателям температуры: 4.67 ± 0.22 и 5.59 ± 0.21 соответственно (t = 3.00; p = 0.004). По среднегодовому количеству осадков различия при этом не выявлены (t = 0.33; p = 0.741).
Таким образом, за рассмотренный исторический период времени произошло повышение среднегодовой температуры, но уровень осадков в регионе в начале XXI в. остался приблизительно на том же уровне, что и в конце XX в. Несомненно, что повышение температуры и сохранение того же режима влажности неизбежно должно было привести к изменению растительности. Наиболее заметные изменения следует отметить в пойменном лесу р. Сакмары вблизи г. Кувандык (место обитания сакмарской популяции рыжей полевки), который за последние 10–15 лет превратился в труднопроходимую урему [29]. Изменения коснулись также пойменного леса р. Самары вблизи с. Платовка (платовская популяция), где наблюдались антропогенная денудация лесного полога и травяного покрова, деградация ягодников (ежевика, дикая вишня) и общее усиление ксерофитности растительности. В районе пос. Тоцкое (тоцкая популяция) пойменный лес за последние 10–15 лет подвергался постоянным неконтролируемым вырубкам и рекреативному воздействию, что привело к почти полному исчезновению насаждений дуба, деградации ягодников и изменению состава ряда связанных с дубом травянистых ассоциаций на левобережье р. Самары. В то же время в пойменном лесу возросла доля инвазивного вида – клена американского, а также вяза шершавого. Изменение климатических условий и состава растительности должно было в свою очередь повлиять на трофические и другие экологические условия пойменных популяций рыжей полевки в данных локалитетах Оренбургской области, а также опосредованно и на изменчивость формы и функциональных свойств нижней челюсти грызунов.
Половой диморфизм по размерам мандибул у рыжей полевки в среднем составил 5.01%. Средние размеры (площадь) нижней челюсти у самцов значимо меньше, чем у самок, в оба периода времени, но в разные годы и разных популяциях их соотношение может варьировать. Поэтому можно предполагать, что и размеры мандибул, и габитуальные размеры животных адаптивно пластичны и имеют функциональное значение. В платовской популяции в оба года наблюдений половой диморфизм по размерам мандибул вообще не проявился, а в сакмарской значимые половые различия обнаружены только в 2015 г. Интересно, что при сравнении площади мандибул в разных популяциях и в разные годы у самок статистически значимые различия не проявились. При аналогичном сопоставлении аллохронных аллотопных выборок значимое своеобразие проявилось только у самцов из платовской популяции 1983 г., которые в этом году оказались такими же крупными, как и самки, в отличие от самцов тоцкой и сакмарской популяций. В 2015 г. нижние челюсти самцов платовской популяции стали, однако, сравнительно мелкими, как и в других популяциях, т.е. половой диморфизм по размерам мандибул в этом случае все же проявился. Таким образом, тот факт, что половой диморфизм по размерам может быть в разной степени выражен в разных популяциях, причем то проявляется, то нивелируется в них в разные годы, косвенно подтверждает функциональную природу различий между полами по размерам.
Оценка регрессии между значениями логарифмов центроидного размера (CS) и первой главной компоненты формы (PCS1) выявила линейную зависимость (p < 0.001), но доля объясненной дисперсии при этом оказалась невелика и составила 14%. В этом случае можно говорить об очень слабой, хотя и статистически значимой аллометрической зависимости формы нижней челюсти от ее размера, которой можно пренебречь. Отсутствие существенных межпопуляционных различий по площади мандибул в соответствующие годы сравнения также позволяет считать, что слабые аллометрические эффекты при сходных средних размерах объектов не влияют на межпопуляционные различия, выявленные по форме мандибул.
Результаты канонического анализа формы нижней челюсти позволяют заключить, что с 1983 по 2015 г. произошли существенные морфогенетические изменения, которые обусловлены не аллометрическими эффектами, а накопленными мелкими многочисленными нелинейными изменениями конфигурации нижней челюсти. Хронографическая изменчивость, выраженная вдоль первой канонической оси, по величине межгрупповой дисперсии более чем в два раза превышает географическую изменчивость (для выборок в 1983 г.), проявившуюся вдоль второй оси. В исходный 1983 г. географическая изменчивость формы мандибул от западной тоцкой до наиболее восточной сакмарской популяции в общем морфопространстве направлена почти под прямым углом (77°) к тренду хронографической изменчивости. Косинус этого угла, выраженного в радианах, в данном случае приблизительно соответствует коэффициенту корреляции Пирсона r = 0.22, т.е. наличию слабой связи. В 2015 г. направления географической и хронографической изменчивости в значительной степени согласуются. В этом случае угол между направлением хронографической и географической изменчивости становится острым (30°30′) и соответствует коэффициенту r = = 0.86, т.е. проявлению сильной связи между ними.
Сходство географической изменчивости нижней челюсти на рубеже XXI в. с хронографической косвенно указывает на то, что градиент условий обитания вида в пространстве во многом согласуется с его изменениями в историческом времени, отражая направленное изменение климата и соответственно его влияние на пойменную растительность и фауну беспозвоночных животных, которыми питаются рыжие полевки. Как уже отмечалось выше, изменение трофических условий должно было затронуть и морфофункциональные свойства мандибул полевок.
Ранее в экспериментальной работе на лабораторных мышах Андерсон с соавт. [12] показали, что длительное питание животных кормом разной консистенции – твердой или мягкой – приводит к статистически достоверным изменениям значений мандибулярных индексов. Нами также установлено, что длительная интродукция южных и северных популяций ондатры (Ondatra zibethicus L.) сопровождалась изменением значений индексов во времени и пространстве, связанных с изменением диеты животных [30]. В другом исследовании при использовании мандибулярных индексов нами [14] выявлены морфофункциональные различия окрасочных морф обыкновенной слепушонки (Ellobius talpinus Pall.) и специфика питания мономорфных и полиморфных по окраске популяций. Поэтому в настоящей работе мандибулярные индексы рассматриваются как морфофункциональные показатели – аналог индексов, использованных в рамках метода морфофизиологических индикаторов [31], характеризующие особенности диеты полевок и способ инструментальной обработки объектов питания.
Сравнение величин морфофункциональных мандибулярных индексов позволяет заключить, что в каждой изученной популяции за более чем 30-летний период наблюдается специфическое изменение этих характеристик у самцов и самок (см. рис. 4, 5): у одних групп происходит усиление грызущей функции, а у других наблюдается усиление конфигурации, связанной с функцией жевания. В сравниваемых аллохронных синтопных популяциях произошли разнонаправленные изменения мандибулярных индексов, главным образом касающиеся функции жевания (индексы TI и TM). Наибольшие функциональные различия по индексам TI и TM выражены между аллохронными выборками самок сакмарской популяции: жевательная функция у самок усиливается от 1983 к 2015 г. Для самцов это не характерно, причем в 2015 г. половые различия по этим индексам нивелируются. Если в платовской популяции в 1983 г. половые различия по индексам TI и TM, с одной стороны, и индексам MI и MM, с другой, не выражены, то в 2015 г. у самок достоверно возрастают значения обоих групп индексов. Другими словами, у самцов значения индексов остались прежними, а у самок произошли существенные изменения, связанные с усилением функций грызения и жевания пищевых объектов. В тоцкой популяции в 1983 г. наблюдались половые различия по обеим группам индексов, причем если у самок были усилены функции грызения и жевания, то у самцов – только функция жевания. В 1983 г. эту популяцию отлавливали в разреженной пойменной дубраве, поэтому, судя по величинам индексов, можно полагать, что самцы и самки специализировались в питании разными объектами: в диете крупных по размерам самок преобладали желуди, а более мелкие самцы, вероятно, предпочитали питание злаками, ягодами (ежевика) и почвенными беспозвоночными. В 2015 г. экологические условия в тоцкой популяции изменились, и половые различия по обеим группам индексов нивелировались. Таким образом, самцы и самки разных популяций в разные годы способны отчасти переключать морфогенез нижней челюсти и реализовать набор конфигураций мандибул, специализированных для питания разными по твердости и инструментальным требованиям к их механической обработке кормовыми объектами.
Начиная с 1998 г. в Оренбургской области наблюдалось существенное уменьшение амплитуды межгодовых колебаний показателя среднегодовой температуры при ее большей средней величине по сравнению с предыдущими 30 годами. Такая многолетняя ситуация должна была способствовать стабилизации разнообразия растительности и состава почвенных беспозвоночных, в том числе и той их части, которой питается рыжая полевка. Возможно, именно это повлияло на меньший разброс величин мандибулярных индексов и уменьшение показателя морфофункционального разнообразия MMFD на втором временном этапе сравнения выборок в 2015 г.
ЗАКЛЮЧЕНИЕ
Проведенное исследование показало, что в исторический период времени за три десятилетия (1983–2015 гг.) на южной границе ареала вида на фоне климатических изменений в направлении повышения среднегодовой температуры, сопряженных с трансформацией растительности, произошло однонаправленное изменение морфогенеза нижней челюсти в географически удаленных популяциях рыжей полевки. Данный исторический процесс направленной хронографической изменчивости сопровождался в общем морфопространстве сходной модификацией направления и географической изменчивости локальных популяций. Изменения формы нижней челюсти одновременно сопряжены с перестройкой ее функций при инструментальной обработке кормовых объектов, причем они могут быть различны у самцов и самок. Зверьки разных полов выступают в популяции фактически как представители разных функциональных морф, обладающих разными инструментальными возможностями при обработке кормовых объектов, что четко проявляется по значениям морфофункциональных мандибулярных индексов самцов и самок (см. рис. 4, 5). При благоприятных условиях, которые, в частности, наблюдались в 2015 г., половые морфофункциональные различия нивелируются, и животные разных полов выполняют в популяции и локальном сообществе грызунов сходные трофические функции, питаясь одним и тем же составом кормов. В иных условиях, как, например, в 1983 г., морфофункциональные половые различия, напротив, усиливаются, расширяя функциональные возможности внутрипопуляционных структурных групп (к таковым могут относиться самцы и самки, возрастные когорты, функциональные физиологические группы – ФФГ [см. 32], биотипы [см. 33] и др.). Действительно, в популяции животные разного пола могут становиться специализированными морфами, которые, с одной стороны, обеспечивают более эффективное использование кормовых ресурсов, а с другой, позволяют снизить уровень трофической конкуренции.
Таким образом, морфофункциональные мандибулярные индексы фактически дополняют метод морфофизиологических индикаторов, предложенный ранее С.С. Шварцем с соавт. [31], обеспечивая соответствующую функциональную интерпретацию различий на внутри- и межпопуляционном уровнях сравнения. Проведенные исследования выявили связь быстрых морфогенетических и морфофункциональных перестроек мандибул полевок с изменением локальных трофических условий, обусловленных разной направленностью растительных сукцессий, связанных в свою очередь с локальными трендами климатических изменений. Подобные быстрые направленные изменения морфогенеза полевок могут быть интерпретированы как пример адаптивных микроэволюционных процессов, происходящих в историческом, а не геологическом масштабе времени.
Работа выполнена в рамках государственного задания № 1201356803 ИЭРиЖ УрО РАН при частичной поддержке Комплексной программы УрО РАН (№ 18-4-4-28). Авторы благодарят к.б.н. Е.Ю. Захарову, к.б.н. А.О. Шкурихина, к.б.н. Т.С. Ослину и Ю.М. Чибиряка за помощь в сборе материала.
Соавторы сердечно поздравляют академика В.Н. Большакова с 85-летним юбилеем.
Список литературы
Яблоков А.В. Изменчивость млекопитающих. М.: Наука, 1966. 364 с.
Яблоков А.В. Популяционная биология: Учеб. пос. для биол. спец. вузов. М.: Высш. шк., 1987. 303 с.
Майр Э. Зоологический вид и эволюция. М.: Мир, 1968. 597 с.
Шварц С.С. Эволюционная экология животных: экологические механизмы эволюционного процесса. Свердловск: УФАН СССР, 1969. 199 с.
Шварц С.С. Экологические закономерности эволюции. М.: Наука, 1980. 278 с.
Большаков В.Н. Пути приспособления мелких млекопитающих к горным условиям. М.: Наука, 1972. 200 с.
Паавер К.Л. Вопросы синтетического подхода к биоморфологии. Таллин: Валгус, 1976. 256 с.
Васильев А.Г. Изоляция расстоянием и дифференциация популяций // Журн. общ. биол. 1984. Т. 45. № 2. С. 164–176.
Павлинов И.Я. Морфологическое разнообразие: общие представления и основные характеристики // Зоологические исследования. М.: Изд-во МГУ, 2008. С. 343–388.
Adams D.C., Rohlf F.J., Slice D.E. A field comes of age: geometric morphometrics in the 21st century // Hystrix, the Italian J. of Mammalogy. 2013. V. 24. № 1. P. 7–14.
Parmesan C. Ecological and evolutionary responses to recent climate change // The Annual Review of Ecology, Evolution, and Systematics. 2006. V. 37. P. 637–669.
Anderson Ph.S.L., Renaud S., Rayfield E.J. Adaptive plasticity in the mouse mandible // BMC Evolutionary Biology. 2014. V. 14. P. 85–93.
Васильев А.Г., Фалеев В.И., Галактионов Ю.К. и др. Реализация морфологического разнообразия в природных популяциях млекопитающих / Отв. ред. Равкин Ю.С. 2-е изд., испр. Новосибирск: Изд-во СО РАН, 2004. 230 с.
Васильев А.Г., Большаков В.Н., Евдокимов Н.Г., Синева Н.В. Морфоразнообразие моно‑ и полиморфных популяций обыкновенной слепушонки: реализуется ли “принцип компенсации” Ю.И. Чернова внутри популяции? // Докл. РАН. 2016. Т. 468. № 1. С. 118–121. [Vasil’ev A.G., Bolshakov V.N., Evdokimov N.G., Sineva N.V. Morphological diversity of mole vole mono‑ and polymorphic populations: does Chernov’s “compensation principle” work within a population? // Doklady Biological Sciences. 2016. V. 468. № 1. P. 118–121.]
Васильев А.Г., Васильева И.А., Шкурихин А.О. Геометрическая морфометрия: от теории к практике. М.: Тов-во научн. изд. КМК, 2018. 471 с.
Васильев А.Г., Васильева И.А., Городилова Ю.В., Добринский Н.Л. Принцип компенсации Ю.И. Чернова и влияние полноты состава сообщества грызунов на изменчивость популяции рыжей полевки (Clethrionomys glareolus) на Среднем Урале // Экология. 2017. № 2. С. 116–125. [Vasil’ev A.G., Vasil’eva I.A., Gorodilova Yu.V., Dobrinskii N.L. Chernov’s compensation principle and the effect of rodent community completeness on the variability of Bank Vole (Clethrionomys glareolus) population in the Middle Urals // Rus. J. of Ecology. 2017. V. 48. № 2. P. 161–169.]
Rohlf F.J., Slice D. Extension of the Procrustes method for the optimal superimposition of landmarks // Syst. Zool. 1990. V. 39. № 1. P. 40–59.
Zelditch M.L., Swiderski D.L., Sheets H.D., Fink W.L. Geometric Morphometrics for Biologists: A Primer. New York: Elsevier Acad. Press, 2004. 437 p.
Klingenberg C.P. MorphoJ: an integrated software package for geometric morphometrics // Mol. Ecol. Resour. 2011. V. 11. P. 353–357.
Rohlf F.J. tpsUtil. Version 1.64. Department of Ecology and Evolution, State University of New York, Stony Brook (program). 2015a. (http://life.bio.sunysb.edu.morph/).
Rohlf F.J. tpsDig. Version 2.22. Department of Ecology and Evolution, State University of New York, Stony Brook (program). 2015b. (http://life.bio.sunysb.edu.morph/).
Rohlf F.J. Shape statistics: Procrustes superimpositions and tangent spaces // J. of Classification. 1999. V. 16. P. 197–223.
Anderson M.J. A new method for non-parametric multivariate analysis of variance // Australian Ecology. 2001. V. 26. P. 32–46.
Воронцов Н.Н. Низшие хомякообразные (Cricetidae) мировой фауны. Ч. I. Морфология и экология. Фауна СССР. Млекопитающие. Т. III, вып. 6. Л.: Наука, 1982. 451 с.
Васильев А.Г., Васильева И.А., Городилова Ю.В., Чибиряк М.В. Соотношение морфологического и таксономического разнообразия сообществ грызунов в зоне влияния Восточно-Уральского радиоактивного следа на Южном Урале // Экология. 2010. № 2. С. 119–125. [Vasil’ev A.G., Vasil’eva I.A., Gorodilova Y.V., Chibiryak M.V. Relationship between morphological disparity and taxonomic diversity in rodent communities in the zone of influence from the Eastern Ural radioactive trace in the Southern Urals // Russ. J. of Ecology. 2010. V. 41. № 2. P. 153–158.]
Hammer Ø., Harper D.A.T., Ryan P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis // Palaeontologia Electronica. 2001. V. 4, № 1. 9 p.
Hedges L.V., Olkin I. Statistical methods for Meta-Analysis. New York: Acad. Press, 1985. 369 p.
Cohen J. A power primer // Psychological Bulletin. 1992. V. 112. № 1. P. 155–159.
Васильев А.Г., Большаков В.Н., Васильева И.А. и др. Фауна насекомоядных млекопитающих и грызунов Губерлинского мелкосопочника (Оренбургская область) // Фауна Урала и Сибири. 2017. № 1. С. 223–244.
Васильев А.Г., Большаков В.Н., Васильева И.А., Синева Н.В. Последствия интродукции ондатры в Западной Сибири: морфофункциональный аспект // Российский журн. биологич. инвазий. 2016. № 4. С. 2–13. [Vasil’ev A.G., Bol’shakov V.N., Vasil’eva I.A., Sineva N.V. Aftereffects of Muskrat Introduction in Western Siberia: Morphological and Functional Aspects // Rus. J. of Biological Invasions. 2017. V. 8. № 1. P. 1–9.]
Шварц С.С., Смирнов В.С., Добринский Л.Н. Метод морфофизиологических индикаторов в экологии наземных позвоночных. Свердловск, 1968. 387 с.
Оленев Г.В. Альтернативные типы онтогенеза цикломорфных грызунов и их роль в популяционной динамике (экологический анализ) // Экология. 2002. № 5. С. 341–350.
Васильев А.Г. Эпигенетические основы фенетики: на пути к популяционной мерономии. Екатеринбург: Изд-во “Академкнига”, 2005. 640 с.
Дополнительные материалы отсутствуют.