

Экология, 2019, № 5, стр. 392-400
Особенности стациального распределения мышевидных грызунов в предгорной зоне Республики Дагестан
a Прикаспийский институт биологических ресурсов ДФИЦ РАН
368025 Махачкала, ул. М. Гаджиева, 45, Россия
b Дагестанский государственный университет
367001 Махачкала, ул. М. Гаджиева, 43а, Россия
* E-mail: mmsh78@mail.ru
Поступила в редакцию 11.01.2018
После доработки 17.12.2018
Принята к публикации 29.01.2019
Аннотация
Проанализированы особенности пространственного распределения 4 видов мышевидных грызунов (Sylvaemus fulvipectus, Cricetulus migratorius, Microtus socialis, Dryomys nitedula) в лесных ассоциациях Предгорной зоны Республики Дагестан. Выбраны 16 параметров среды, из которых 10 значимо дискриминировали межвидовые различия. Показано распределение видов грызунов относительно учтенных в описаниях характеристик среды, выявлены видовые предпочтения и требования к среде у каждого вида. Так, желтобрюхая мышь и серый хомячок встречались во всех стациях, тогда как лесная соня и общественная полевка были более избирательны в выборе стаций. Как следствие показатели ширины пространственных ниш у желтобрюхой мыши и серого хомячка были больше, чем у лесной сони и общественной полевки. Результаты попарного сравнения пространственных ниш рассматриваемых видов грызунов выявили низкие значения их перекрывания.
Считается, что пространственная структурированность сообщества определяет наиболее эффективное использование ресурсов среды (пищевых, защитных), обеспечивает максимальную реализацию биотического потенциала популяций видов. Обилие и особенности пространственного размещения видов свидетельствуют о степени благополучия их существования в природной среде [1–4]. Устойчивое пребывание экологически схожих видов на определенной территории возможно благодаря комплексу адаптаций к данным условиям среды, что позволяет этим видам относительно легко переживать неблагоприятные воздействия среды или межвидовую конкуренцию.
В этой связи горные леса могут являться хорошей площадкой для изучения особенностей территориального распределения экологически близких видов и в целом путей формирования структуры данных сообществ. Наличие растительных, почвенных и микроклиматических различий способствует формированию разнообразных стаций и микростаций, которые могут быть заняты экологически близкими видами, что в конечном итоге и позволяет максимально плотно “упаковать” виды.
Основная идея, объясняющая механизмы формирования структуры сообществ – концепция экологической ниши. В концепции Дж. Хачинсона [5] теоретически обоснован поиск методов изучения и отображена реальная экологическая ниша [6]. Традиционно структуру сообществ рассматривали путем сопоставления обобщенных характеристик экологических ниш, используя различные индексы (Симпсона, Шеннона, Пианки и др.) [7, 8]. Недостатком данного подхода считается возможность потери информации о путях разделения ресурсов, субъективность в выборе осей нишевого гиперпространства; использование индексов не позволяет выявить ключевые факторы либо отсечь второстепенные [9]. Позже широкое распространение получили методы многомерной статистики [9–12], которые позволили использовать в качестве осей экологической ниши весь комплекс условий и ресурсов среды. Данный подход не противоречит представлению о многомерной экологической нише, предложенному Хатчинсоном [5]. Наиболее широкое признание получили анализ главных компонент, дисперсионный и дискриминантный анализы, с помощью которых можно выявить ключевые факторы, определяющие структуру сообщества, провести редукцию числа средовых параметров при описании пространственных ниш [9, 10, 12–15].
В результате качество местообитания может выступать ключевой характеристикой, благодаря которой виды по-разному представлены в биоценозах. Более того, есть мнение, что пространственное распределение мелких млекопитающих может определяться средовыми и популяционными факторами локального проявления [16]. В качестве примера отметим работы, в которых показана особенность микросредового распределения мелких млекопитающих, дана оценка различным факторам [2, 6, 10–18].
Цель настоящей работы – оценка стациального распределения совместно обитающих видов мышевидных грызунов и анализ факторов, определяющих положение видов в общем пространстве в лесных ассоциациях Предгорной зоны Республики Дагестан.
МАТЕРИАЛ И МЕТОДИКА
Горная часть Дагестана характеризуется гетерогенной структурой рельефа – даже близко расположенные участки различных склонов существенно различаются по микроклиматическим характеристикам, растительной структуре, продукции и т.д. Широкий спектр условий обитания должен определять и различия в стратегии адаптаций симпатрических видов.
Климат в районе исследования умеренно теплый, средняя температура воздуха зимой +1.5°C, летом +17°C. В ландшафтном отношении изучаемый район – это предгорья (начало переходной зоны между равнинной и горной частями Республики Дагестан). Количество осадков около 350 мм в год. Атмосферные осадки распределяются неравномерно в зависимости от экспозиции склонов, высоты хребтов, близости к Каспийскому морю. Почвенный покров представлен каштановыми карбонатными, коричневыми и бурыми лесными почвами [19].
Растительность в районе исследования включает в себя степные и лесостепные формации. Среди степной растительности главными формациями являются разнотравно-полынно-злаковые и заросли ксерофитных кустарников: держи-дерево (Paliurus spina-christi), крушина Палласа (Rhamnus pallasii) и др. В травостое преобладают ковыль волосатик (Stipa capillata), тимофеевка степная (Phleum phleoides), полынь таврическая (Artemisia taurica) и др. Лесостепной пояс формируется за счет сосново-дубовых, дубовых и можжевеловых редколесий: дуб пушистый (Quercus pubescens) и скальный (Q. petraea), сосна Коха (Pinus kochiana), граб кавказский (Carpinus betulus), можжевельник продолговатый (Juniperus oblonga). Травяной покров разреженный, преимущественно это ксерофитные виды: ячмень заячий (Hordeum leporinum), костер мягкий (Bromus mollis), мятлик луковичный (Poa bulbosa), дубровник белый (Teucrium polium) [20].
Исследования проведены вдоль северного склона Нарат-Тюбинского хребта (площадь северного склона хребта около 50 км2), расположенного в 5 км от г. Махачкала Республики Дагестан (42°54′ с.ш., 47°27′ в.д.). Материал собирали в типичных для района стациях (молодой лес, высокоствольные участки леса и опушка, открытые участки, каменистые осыпи, кустарниковые ассоциации). Основными критериями для выделения стаций были структура растительности (плотность и размеры древесно-кустарниковой растительности) и состояние почвы. Доля выделенных стаций составляла: молодой лес – 23%, высокоствольные участки леса – 34%, опушка леса – 9%, открытые участки – 13%, кустарниковые ассоциации – 16%, каменистые осыпи – 5%. В одних случаях рассматриваемые стации располагались по соседству, в других они не пересекались. Сборы проводили в мае, июле и сентябре 2013–2014 гг. Места проведения исследования располагались на высоте 200–450 м над ур. м.
Животных отлавливали методом ловушко-линий [21]. Для этого использовали давилки-плашки, которые устанавливали в линии (по 100 шт., расстояние между ловушками 10 м, экспозиция в течение 1 сут). Ежедневно линию переставляли на новое место (параллельно предыдущей). Расстояние между линиями составляло не меньше 50 м. В течение одного тура перестановку ловушек осуществляли в течение 3–4 сут. В качестве приманки использовали семена подсолнечника, смоченные в растительном масле. Все значения уловов переводили на 100 ловушко-ночей. Уровни численности оценивали по Г.А. Новикову [22]: низкая – меньше 10 особей на 100 ловушко-суток, средняя – от 0 до 10 особей и высокая – более 30 особей.
В качестве модельных объектов рассматривали желтобрюхую мышь (Sylvaemus fulvipectus Ognev, 1924), серого хомячка (Cricetulus migratorius Pallas, 1773), общественную полевку (Microtus socialis Pallas, 1773) и лесную соню (Dryomys nitedula Pallas, 1779). За период исследования было отработано 2400 ловушко-ночей и отловлено 283 особи, в том числе лесная соня – 54, желтобрюхая мышь – 144, общественная полевка – 59, серый хомячок – 26. Сбор материала проходил таким образом, чтобы учетные линии охватывали типичные стации.
Ключевым моментом в работе было соотнесение характеристик грызунов и среды в районе исследования. Чтобы оценить приуроченность рассматриваемых видов грызунов к выделенным стациям, а не целым биотопам, геоботаническое описание и характеристику почвы проводили строго в точках поимок животных (в каждой точке от 2 до 4 замеров в радиусе до 2 м), используя стандартные методы полевой геоботаники [23]. Надземную фитомассу учитывали методом укосов на площадках размером 1 м2 с разбором укосов по группам (злаки, разнотравье), проективного покрытие растительности – при помощи квадрата-сетки размером 1 м2. Плотность почвы определяли целиндром-буром на глубине до 0.1 м в то же время, что и остальные сборы, влажность почвы – термостатно-весовым методом как разницу между массой взятой из почвы пробы и массой пробы, высушенной в термостате.
Положение пространственных ниш рассмотренных видов грызунов в пространстве условий и ресурсов (учтенных в описаниях параметров) среды исследовали при помощи дискриминантного анализа. Для этого использовали характеристики среды только в местах поимок особей. В результате мы получили характеристику не всего пространства условий и ресурсов в районе исследования, а лишь занятую видами область этого пространства [9, 10].
Выбор параметров среды осуществляли на основе субъективных представлений об их влиянии на распределение рассматриваемых видов грызунов в районе исследования. При этом мы исходили из того, что ключевыми факторами, определяющими состояние популяции вида в среде, является наличие кормов и убежищ. Выделено 16 параметров ландшафтной приуроченности местообитаний рассматриваемых видов грызунов. В их число вошли: площадь покрытия поверхности почвы камнями (%) – ППППК, фитомасса злаков на открытых участках (кг/га) – ФЗО, фитомасса злаков в лесу (кг/га) – ФЗЛ, фитомасса разнотравий на открытых участках (кг/га) – ФРО, фитомасса разнотравий в лесу (кг/га) – ФРЛ, проективное покрытие злаков на открытых участках (%) – ППЗО, проективное покрытие злаков в лесу (%) – ППЗЛ, проективное покрытие разнотравий на открытых участках (%) – ППРО, проективное покрытие разнотравий в лесу (%) – ППРЛ, плотность древостоя (шт./га) – ПД, сомкнутость крон деревьев (%) – СКД, плотность кустарников (шт/га) – ПК, высота кустарников (м) – ВК, сомкнутость крон кустарников (%) – СКК, влажность почвы (%) – ВП и плотность почвы до 30 см глубины (г/см3) – ПП. Все выбранные показатели среды подчинялись закону нормального распределения.
Особенности стациального распределения видов грызунов (пространственная ниша) выражена в виде облака точек в пространстве дискриминантных функций, где каждая точка соответствует определенному описанию среды в местах поимок особей. Следовательно, центроид распределенных в пространстве дискриминантных функций точек считался как центр пространственной ниши каждого рассматриваемого вида грызуна, а ширина ниши представлена как среднее отклонение от точек регистрации видов до центроида данного вида [9, 13]. Распределения по параметрам среды не отличались от нормальных (по Колмогорову–Смирнову), и объем выборки по каждому виду составил: D. nitedula (n = 24), S. fulvipectus (n =29), C. migratorius (n = 25), M. socialis (n = 29).
Величину перекрывания ниш рассчитывали как процент неправильно классифицированных точек в классификационных матрицах апостериорных вероятностей при попарных сравнениях видов сообщества грызунов. Значения перекрывания в диапазоне от 0 до 0.25 считали низкими, от 0.25 до 0.5 – средними, от 0.5 до 0.75 – высокими и от 0.75 и выше – очень высокими. Статистическая обработка данных выполнена в программе “StatSoft STATISTICA for Windows 10” с использованием дискриминантного анализа.
РЕЗУЛЬТАТЫ
Данные о стациальном распределении и обилии рассматриваемых видов грызунов в районе исследования приведены в табл. 1. Особи желтобрюхой мыши и серого хомячка встречались во всех выделенных стациях, а лесная соня и общественная полевка обитали в древесно-кустарниковых зарослях и на открытых участках. Всплески численности характерны для желтобрюхой мыши. На момент проведения исследования ее обилие характеризовалось как низкое [22], а обилие остальных видов грызунов оставалось на уровне стабильно низкое (см. табл. 1).
Таблица 1.
Особенности стациального распределения и показатели относительного обилия видов рассматриваемого сообщества мышевидных грызунов в районе исследования (особей /100 лов.- ночей)
| Стации | D. nitedula (n = 54) |
S. fulvipectus (n = 144) |
M. socialis (n = 59) |
C. migratorius (n = 26) |
|---|---|---|---|---|
| Высокоствольный лес | 1 ± 0.1 | 6.7 ± 0.4 | – | 0.23 ± 0.02 |
| Молодой лес | 5.6 ± 0.32 | 8.4 ± 0.92 | – | 3.32 ± 0.18 |
| Опушка леса | 5.3 ± 0.37 | 8.0 ± 0.44 | 1.31 ± 0.07 | 1.43 ± 0.07 |
| Кустарниковые заросли | 6.7 ± 0.41 | 10.3 ± 0.52 | – | 2.95 ± 0.23 |
| Открытые участки | – | 10.8 ± 0.57 | 10.14 ± 0.62 | 2.17 ± 0.3 |
| Каменистые выступы | – | 6.2 ± 0.36 | 3.72 ± 0.23 | 0.37 ± 0.21 |
| Средняя численность | 4.7 ± 0.55 | 8.41 ± 1.36 | 5.24 ± 0.27 | 2.86 ± 0.57 |
Значимые характеристики среды для каждого вида грызунов были выделены при помощи дискриминантного анализа с последующим построением многомерной модели сообщества. Дискриминантная функция корректно классифицировала 76.8% случаев, что соответствует высоким качеству распознавания и эффективности дикриминантной функции. В ходе дискриминантного анализа из 16 параметров среды было выделено 10 значимо дискриминировавших виды, из них наибольший вклад в межвидовые различия вносили 4 переменные (проективное покрытие разнотравья на открытых участках, проективное покрытие злаков на открытых участках, плотность почвы, проективное покрытие злаков в лесу) (табл. 2).
Таблица 2.
Результаты дискриминантного анализа для параметров среды (Лямбда Уилкса – 0.108, F (48.24) = = 5.53, p < 0.000, df = 196)
| Параметры | ДФ 1 | ДФ 2 | ДФ 3 | Лямбда Уилкса |
Частная лямбда Уилкса |
Критерий Фишера |
Р |
|---|---|---|---|---|---|---|---|
| ПП, г/см3 | 0.423 | –0.092 | –0.426 | 0.123 | 0.880 | 4.126 | 0.012 |
| ВП, % | 0.086 | –0.146 | –0.143 | 0.109 | 0.988 | 0.305 | 0.821 |
| ППППК, % | –0.091 | 0.062 | 0.091 | 0.109 | 0.992 | 0.203 | 0.894 |
| ФЗОУ, кг/га | –0.806 | 1.101 | –3.106 | 0.122 | 0.888 | 3.349 | 0.023 |
| ФЗЛ, кг/га | –0.106 | –0.431 | –0.691 | 0.110 | 0.977 | 0.603 | 0.615 |
| ФРОУ, кг/га | –0.580 | 2.022 | 0.189 | 0.116 | 0.932 | 2.547 | 0.047 |
| ФРЛ, кг/га | 0.413 | 0.437 | 0.411 | 0.119 | 0.908 | 2.709 | 0.050 |
| ППЗОУ, % | 1.844 | –2.122 | 2.748 | 0.131 | 0.828 | 5.589 | 0.001 |
| ППЗЛ, % | 0.421 | 1.475 | –0.552 | 0.123 | 0.879 | 3.641 | 0.016 |
| ППРОУ, % | –0.264 | –2.281 | –0.667 | 0.149 | 0.727 | 9.996 | 0.001 |
| ППРЛ, % | 0.332 | –0.527 | –0.285 | 0.111 | 0.976 | 0.637 | 0.593 |
| ПД, ед./га | –0.290 | –1.224 | –0.332 | 0.113 | 0.951 | 1.366 | 0.258 |
| СКД, % | 0.238 | –0.813 | 0.083 | 0.119 | 0.907 | 2.717 | 0.050 |
| ПК, ед./га | –1.240 | –0.165 | 1.047 | 0.115 | 0.934 | 1.873 | 0.141 |
| ВК, м | –2.028 | –2.792 | –1.289 | 0.134 | 0.806 | 3.126 | 0.018 |
| СКК, % | 1.796 | 3.447 | 0.758 | 0.139 | 0.773 | 3.653 | 0.015 |
| Собственные числа | 2.49 | 1.25 | 0.17 | ||||
| % объясненной дисперсии | 63.7 | 32 | 4.3 | ||||
Примечание. ПП – плотность почвы, ВП – влажность почвы, ППППК – площадь покрытия поверхности почвы камнями, ФЗОУ – фитомасса злаков на открытых участках, ФЗЛ – фитомасса злаков в лесу, ФРОУ – фитомасса разнотравий на открытых участках, ФРЛ – фитомасса разнотравий в лесу, ППЗОУ – проективное покрытие злаков на открытых участках, ППЗЛ – проективное покрытие злаков в лесу, ППРОУ – проективное покрытие разнотравий на открытых участках, ППРЛ – проективное покрытие разнотравий в лесу, ПД – плотность древостоя, СКД – сомкнутость крон деревьев, ПК – плотность кустарников, ВК – высота кустарников и СКК – сомкнутость крон кустарников; выделенные полужирным шрифтом значения – статистически значимы, ДФ – стандартизованный коэффициент дискриминантной канонической функции.
Учитывая сезонную вариабельность ряда параметров среды, была посезонно оценена их роль для каждого вида грызунов отдельно, используя дискриминатный анализ. В табл. 3 показаны значения 8 характеристик травянистого покрова, которые достоверно отличались посезонно для каждого вида рассматриваемого сообщества грызунов.
Таблица 3.
Анализ посезонной вариабельности ряда показателей среды для каждого вида рассматриваемого сообщества грызунов (по результатам дискриминантного анализа)
| Параметры | Вид | |||
|---|---|---|---|---|
| D. nitedula | S. fulvipectus | M. socialis | C. migratorius | |
| ФЗОУ, кг/га | – | p < 0.001 | p < 0.001 | p < 0.05 |
| ФЗЛ, кг/га | p < 0.05 | p < 0.05 | – | |
| ФРОУ, кг/га | – | p < 0.001 | ||
| ФРЛ, кг/га | p > 0.05 | – | ||
| ППЗОУ, % | – | p < 0.001 | ||
| ППЗЛ, % | p > 0.05 | – | ||
| ППРОУ, % | – | p < 0.001 | ||
| ППРЛ, % | p > 0.05 | – | ||
Примечание. ФЗОУ – фитомасса злаков на открытых участках, ФЗЛ – фитомасса злаков в лесу, ФРОУ – фитомасса разнотравий на открытых участках, ФРЛ – фитомасса разнотравий в лесу, ППЗОУ – проективное покрытие злаков на открытых участках, ППЗЛ – проективное покрытие злаков в лесу, ППРОУ – проективное покрытие разнотравий на открытых участках, ППРЛ – проективное покрытие разнотравий в лесу; прочерк – отсутствие вида в отловах.
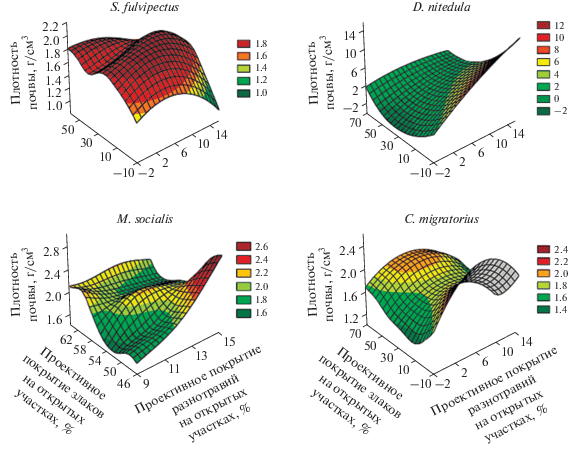
На рис. 1 представлены данные о предпочтении рассматриваемыми видами грызунов значимых средовых переменных. Так, желтобрюхая мышь и серый хомячок встречались в более широком диапазоне значений средовых переменных, а лесная соня и общественная полевка – в более узком диапазоне. Трехмерная картина пространственных ниш исследованных видов грызунов в системе координат из трех максимально дифференцирующих виды переменных среды показана на рис. 2: проекции желтобрюхой мыши и серого хомячка похожи по форме, а лесной сони и общественной полевки имеют специфичную форму.
Рис. 1.
Распределение предпочтений четырех видов грызунов на осях основных переменных, характеризующих среду обитания; среднее – среднеарифметическое значение параметра среды, ст. ош. – стандартная ошибка, ст. откл. – стандартное отклонение.
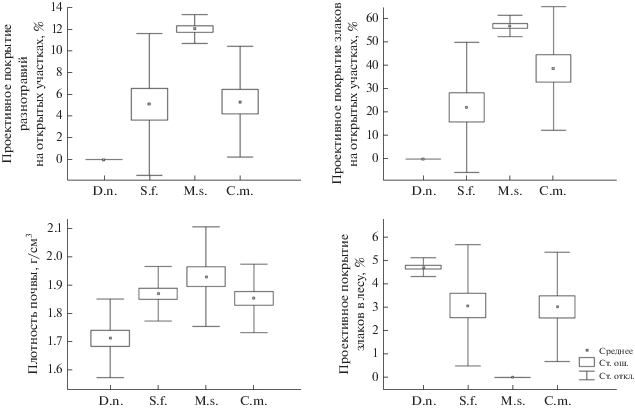
Рис. 2.
Трехмерные проекции пространственных ниш видов в системе координат, оси которой образованы тремя основными разделяющими виды переменными – характеристиками среды обитания.
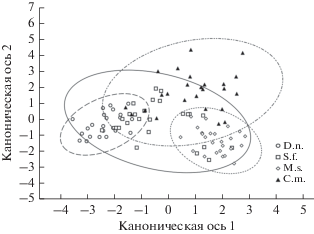
На рис. 3 показано положение рассматриваемых видов грызунов в пространстве, оси которого образованы дискриминантными функциями. Характерно отсутствие обособленных группировок видов. Так, желтобрюхая мышь и серый хомячок имели относительно широкие ниши. Оба вида, встречаясь во всех стациях, с различной степенью перекрывались с остальными видами сообщества, в то время как распределение лесной сони и общественной полевки было более узко локализованным.
Рис. 3.
Диаграмма рассеяния канонических значений видов рассматриваемого сообщества грызунов в пространстве дискриминантных функций.
Используя количество неправильно классифицированных точек в классификационных матрицах апостериорных вероятностей, мы получили оценки перекрываний пространственных ниш видов по рассмотренным параметрам среды. Во всех парах видов грызунов значения перекрывания ниш оказались низкими, а пара лесная соня и общественная полевка вообще не перекрывались (табл. 4).
ОБСУЖДЕНИЕ
В ходе проведенной работы были описаны особенности пространственного распределения видов сообщества грызунов в мозаичных биотопах Предгорной зоны Республики Дагестан. Использование возможностей дискриминантного анализа позволило описать распределение рассмотренных видов грызунов относительно 16 параметров среды, выделив из них значимо влияющие на распределение и показав видовые предпочтения и требования к среде каждого вида. Полученные результаты свидетельствуют о том, что пространственное распределение видов грызунов во многом обусловлено особенностью растительного покрова. Такие параметры, как проективное покрытие травянистой растительности и фитомасса, играли существенную роль при выборе микроместообитаний.
Роль структуры растительности и состояния почвы в характере пространственного распределения грызунов отмечалась во многих работах [9, 11, 15, 24–26]. Так, Б. Красновым с соавт. [27] показано, что пространственная сегрегация 12 пустынных видов грызунов определялась тремя факторами среды: твердость почвы, рельеф и плотность растительности. Куда более значительна роль растительности в лесных биоценозах. В хвойномелколиственных комплексах северной Беларусии были выявлены резко отличающиеся ландшафты, которые формировали микроклимат в этих биотопах, что в свою очередь привело к существенным различиям в видовом богатстве и обилии мелких млекопитающих [28–30]. А. Кокбурн и В. Лидекер [31] показали, что гетерогенность среды обитания для M. californicus в значительной мере объясняется неоднородностью растительного покрова. В целой серии работ освоение пространства показано путем различных стратегий сбора корма и отчасти избегания хищников [10, 15, 30, 32]. Создание видом своей особой структуры размещения в рамках предоставляемых средой пространственных ресурсов является важным адаптивным механизмом, обеспечивающим эффективное использование ресурсов в структуре многовидовых сообществ, неоднородность которых определяется многообразием экологических потребностей различных видов [33].
Для желтобрюхой мыши и серого хомячка характерен эврибиотный тип распределения, а лесная соня и общественная полевка отличались более специализированным типом распределения. Широкая биотопическая и стациальная приуроченность желтобрюхой мыши и серого хомячка описана как на открытых участках [34–36], так и в лесных биотопах [34, 37]. Относительно желтобрюхой мыши есть мнение, что ее распространение на север ограничивают ландшафтно-климатические условия и конкуренция с малой лесной мышью (Sylvaemus uralensis) [34]. Лесная соня, несмотря на обширный ареал, имеет тесную приуроченность к вертикальной ярусности и предпочитает местообитания с высокой степенью сомкнутости крон, хорошо развитыми кустарниковой растительностью и молодым лесом [38–41]. Вертикальная структура является одним из основных путей разделения общего пространства среди древесных видов млекопитающих [42, 43]. Общественная полевка – вид открытых пространств, встречающийся в густых травянистых ассоциациях [44, 45]. В Дагестане и Калмыкии поселения общественной полевки приурочены к злаково-полынным степям [44, 46]. Нами отмечены полевки на участках со слаборазвитой кустарниковой растительностью. В последние десятилетия на территории Северо-Западного Прикаспия на фоне изменения климата, сокращения пастбищной нагрузки в 1990-е годы и замены коренной растительности полупустыни выявлен значительный рост общественной полевки, домовой мыши и серого хомячка – типичных представителей высокотравных сухих степей [45].
Отмеченные примеры демонстрируют ключевую роль флористических и микроклиматических условий в ходе пространственного распределения рассматриваемых видов грызунов. Обитая в пределах общего биотопа, виды могут занять видоспецифические стации или микростации, что позволяет им сосуществовать. Это сформировало у ряда экологов мнение о главенствующей роли пространственной сегрегации в формировании структуры сообществ [3, 4, 11, 14, 16, 18, 30, 47]. На момент проведения исследований степень сегрегации пространственных ниш у видов данного сообщества грызунов показала крайне низкие значения, что может говорить о наличии достаточного резерва для заполнения рассматриваемой среды новыми видами грызунов. Также следует отметить, что проведенный ранее анализ рационов питания рассмотренных видов грызунов выявил [48] вариацию в диапазоне от низких до средних значений перекрывания трофических ниш.
Работа выполнена при финансовой поддержке РФФИ (проект № 13-04-00022).
Список литературы
Шилов И.А. Экология. М.: Высшая школа, 2000. 512 с.
Лукьянова Л.Е., Бобрецов А.В. Локальное распределение численности симпатрических видов лесных полевок в микросредовых условиях дестабилизированных и стабильных местообитаний // Успехи современной биологии. 2008. Т. 128. № 5. С. 517–528.
Rosenzweig M.L. Habitat selection and population interactions: the search of mechanism // Amer. Nat. 1991. № 137. P. 5–28.
Scott D.M., Dunstone N. Environmental determinants of the composition of desert-living rodent communities in the northeast Badia region of Jordan // J. of Zoology. 2000. № 251. P. 481–494.
Hutchinson G.E. Homage to Santa Rosalia or why are there so many kinds of animals? // American Naturalist. 1959. V. XCIII. № 870. P. 145–159.
Пузаченко Ю.Г., Кузнецов Г.В. Экологическая дифференциация грызунов сезонно-влажных тропических лесов северного Вьетнама // Зоол. журн. 1998. Т. 77. № 1. С. 117–132.
Hurlbert S.H. The measurement of niche overlap and some relatives // Ecology. 1978. № 59. P. 67–77.
Krebs C.J. Ecological methodology. Menlo Park, CA. Addison-Wesley Educational Publishers, Inc. 2nd ed. 1999. 620 p.
Роговин К.А., Шенброт Г.И. Структурные аспекты организации сообществ наземных позвоночных на примере пустынных грызунов Монголии // Успехи современной биологии. 1993. Т. 113. № 2. С. 198–222.
Shenbrot G.I., Krasnov B. Temporal dynamics in spatial organization of rodent community in the Negev Highlands (Israel) // J. of Zoology. 2004. № 263. P. 207–218.
Stevens R.D., Tello J.S. Micro- and macrohabitat associations in Mojave desert rodent communities // J. of Mammalogy. 2009. V. 90 № 2. P. 388–403.
Kubiak B.B., Galiano D., Ochotorena de Freitas T.R. Sharing the space: distribution, habitat segregation and delimitation of a new sympatric area of subterranean rodents // PLoS ONE. 2015. V. 10. № 4. P. 1–10.
Шенброт Г.И. Экологические ниши, межвидовая конкуренция и структура сообщества наземных сообществ // Итоги науки и техники. Зоология позвоночных. М.: ВИНИТИ, 1987. Т. 14. С. 5–70.
Лукьянова Л.Е. Сопряженность симпатрических видов мелких млекопитающих в контрастных условиях среды // Экология. 2013. № 1. С. 65–72.
Kalcounis-Ruppell M.C., Millar J.S. Partitioning of space, food, and time by syntopic Peromyscus boylii and P. californicus // J. of Mammalogy. 2002. V. 83. № 2. P. 614–625.
Буяльска Г., Лукьянов О.А., Мешковска Д. Детерминанты локального пространственного распределения численности островной популяции рыжей полевки // Экология. 1995. № 1. С. 35–46.
Лукьянова Л.Е., Бобрецов А.В. Выбор рыжей полевкой (Clethrionomys glareolus Schreber, 1780) микроместообитаний в стабильных и дестабилизироанных условиях среды // Вестник Томского гос. ун-та. Биология. 2014. № 4(28). С. 88–107.
Zhong W., Wang G., Zhou Q. et al. Spatial niche partitioning of coexisting small mammals in sand dunes // Italian J. of Zoology. 2016. V. 83. № 2. P. 248–254.
Акаев Б.А. Рельеф. Физическая география Дагестана. М.: Школа, 1996. С. 112–150.
Львов П.Л., Абачев К.Ю. Растительность Предгорного Дагестана. Физическая география Дагестана. Ростов-на-Дону: РДПИ, 1984. С. 101–121.
Кучерук В.В. Новое в методике количественного учета вредных грызунов и землероек в СССР // Вопросы организации и методы учета ресурсов фауны позвоночных. М.: Изд-во АН СССР, 1963. С. 159–183.
Новиков Г.А. Полевые исследования экологии наземных позвоночных животных. М.: Сов. наука, 1949. 502 с.
Воронов А.Г. Геоботаника. М.: Высшая школа, 1973. 2 изд. 383 с.
Shenbrot G.I., Rogovin K. A., Heske E.J. Comparison of niche-packing and community organization in Asia and North America // Aust. J. Zool. 1994. № 42. P. 479–499.
Kelt D.A., Brown J.H., Rogovin K., Shenbrot G. Patterns in the structure of Asian and North American desert small mammal communities // J. of Biogeography. 1999. № 26. P. 825–842.
Abu Baker M., Patterson B.D. Patterns in the local assembly of Egyptian rodent faunas: Co-occurrence and nestedness // J. of Arid Environments. 2011. № 75. P. 14–19.
Krasnov B., Shenbrot G., Khokhlova I., Ivanitskaya E. Spatial patterns of rodent communities in the Ramon erosion cirque, Negev Highlands, Israel // J. of Arid Environments. 1996. № 32. P. 319–327.
Виноградов В.В. Сравнительная характеристика сообществ мелких млекопитающих горных лесов юга Средней Сибири // Вестник Томского гос. ун-та. Биология. 2010. № 3(11). С. 47–59.
Абрамов С.А., Виноградов В.В. Экологическая дифференциация мышевидных грызунов лесного пояса гор юга Средней Сибири // Поволжский экологич. журн. 2012. № 4. С. 363–375.
Борякова Е.Е., Лямина Н.С. Пространственная структура сообществ мелких млекопитающих и ее связь с фитоценозом // Вестник ОГУ. 2013. № 6. С. 138–142.
Cockburn A., Lidicker W.Z. Microhabitat heterogeneity and population ecology of a herbivorous rodent, Microtus californicus // Oecologia. 1983. V. 59. P. 167–177.
Роговин К.А. Динамика пространственной организации сообществ мелких млекопитающих (на примере грызунов Заалтайской Гоби Монголии) // Журн. общ. биол. 1998. Т. 59. № 5. С. 477–497.
Проскурина Н.С. Еще один подход к сопряженному анализу хорологической структуры растительного покрова и населения мышевидных грызунов // Экология. 1986. № 6. С. 14–20.
Стахеев В.В., Богданов А.С. К характеристике желтобрюхой мыши (Sylvaemus fulvipectus Ognev) Северного Кавказа // Вестник Южного научного центра РАН. 2007. Т. 3. № 3. С. 101–104.
Кузнецов В.А., Савинецкая Л.Е., Чабовский А.В. Динамика пастбищных экосистем юга Калмыкии: влияние изменения климата и антропогенной нагрузки // Успехи современной биологии. 2011. Т. 131. № 5. С. 460–465.
Neronov V.M., Khlyap L.A., Warshavsky A.A. Diversity of the rodent communities in the Turan Desert region // Integrative Zoology. 2006. № 1. P. 153–161.
Громов И.М., Ербаева M.A. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб.: ЗИН РАН, 1995. 522 с.
Айрапетьянц А.Э. Сони. Л.: ЛГУ, 1983. 187 с.
Россолимо О.Л., Потапова Е.Г., Павлинов И.Я. и др. Сони мировой фауны. М.: МГУ, 2001. 229 с.
Krystufek B., Vohralik V. Distribution of the Forest dormouse Dryomys nitedula (Pallas, 1779) (Rodentia, Myoxidae) in Europe // Mammalian Review. 1994. № 24. P. 161–177.
Magomedov M.Sh. Assessment of the habitat quality of the forest dormouse (Dryomys nitedula) in Daghestan, Russia: role of foods and vegetation structure // Folia Zool. 2015. V. 64. № 4. P. 356–360.
Emmons L.H. Ecology and resource partitioning among nine species of African rainforest squirrels // Ecological Monograph. 1980. V. 50. № 1. P. 31–54.
Sushma H.S., Singh M. Resource partitioning and interspecific interactions among sympatric rain forest arboreal mammals of the Western Ghats, India // Behavioral Ecology. 2006. № 3. P. 479–490.
Шилова С.А., Касаткин М.В. Сравнительный анализ популяционной структуры общественной полевки (Microtus socialis Pall., 1773, Cricetidae, Rodentia) в различных частях ареала // Экология. 2000. № 4. С. 287–294.
Букреева О.М., Лиджи-Гаряева Г.В. Массовая миграция и гибель общественных полевок (Microtus socialis Pallas, 1773) в Северо-Западном Прикаспии // Аридные экосистемы, 2018. Т. 24. № 2(75). С. 78–83.
Щипанов Н.А., Касаткин М.В. Общественная полевка (Microtus socialis Pall.) в измененном ландшафте южного Дагестана: популяционный аспект выживания // Зоол. журн. 1996. Т. 75. Вып. 9. С. 1412–1426.
Bouchon-Navaro Y. Partitioning of food and space resources be chaetodontid fishes on coral reefs // J. of Experimental Marine Ecology and Biology. 1986. № 103. P. 21–40.
Магомедов М.Ш. Анализ питания мышевидных грызунов в предгорной зоне Республики Дагестан (микрогистологический кутикулярно-копрологический метод) // Зоол. журн. 2017. Т. 96. № 5. С. 581–588.
Дополнительные материалы отсутствуют.