

Экология, 2019, № 5, стр. 341-355
Динамика древостоев и их продуктивности на верхнем пределе произрастания в Хибинах на фоне современных изменений климата
П. А. Моисеев a, *, А. А. Галимова a, М. О. Бубнов a, Н. М. Дэви a, В. В. Фомин a, b
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
b Уральский государственный лесотехнический университет
620100 Екатеринбург, Сибирский тракт, 37, Россия
* E-mail: moiseev@ipae.uran.ru
Поступила в редакцию 12.09.2018
После доработки 25.02.2019
Принята к публикации 01.03.2019
Аннотация
В пределах экотона верхней границы древесной растительности на юго-восточном макросклоне горного массива Хибины (Кольский п-ов) были изучены пространственная и возрастная структуры, а также особенности накопления фитомассы елово-березовых древостоев. В результате анализа выявлено, что в последние столетия произошло многократное увеличение сомкнутости и продуктивности древостоев, а верхняя граница редколесий и сомкнутых лесов значительно продвинулась выше в горы. Все это происходило на фоне повышения в этом районе в ХХ в. ранних летних температур и увеличения продолжительности вегетационного периода. Полученные данные помогут смоделировать реакцию горных экосистем региона на будущие изменения климата.
На современном этапе исследований широко обсуждается углерододепонирующая роль наземных экосистем, и особенно лесных сообществ, в связи с увеличением концентрации углекислого газа (CO2) в последнем столетии, который является, по мнению многих ученых, основным парниковым газом, способствующим повышению температуры атмосферы Земли. Протяженность границы леса в горных регионах мира составляет десятки тысяч километров и во многих регионах в связи с изменениями климата произошло их смещение вверх по склонам на сотни метров [1, 2], что увеличило лесопокрытую площадь на тысячи квадратных километров. Несмотря на это количественные исследования накопления фитомассы и особенностей ее динамики в древостоях на верхнем пределе их произрастания проводились крайне редко: на Урале [3], в Скалистых горах [4] и Тибете [5]. В частности, на Южном Урале нами ранее [3] было обнаружено, что с 1910 г. по 2013 г. на фоне повышения здесь ранних летних (на 0.6°С) и зимних (1.8°С) температур и увеличения количества твердых осадков надземная фитомасса березово-еловых древостоев на склонах массива Иремель увеличилась на современной верхней границе сомкнутых лесов в среднем с 0.3 до 95.8 т/га, на границе редколесий – до 45.8 т/га и на границе групп деревьев (редин) – до 4.3 т/га.
В последние десятки лет также выявлены значительные изменения в составе, структуре и высотном положении древостоев на верхнем пределе их произрастания в Фенноскандии [6–9]. На склонах гор Кольского полуострова такие исследования проводились лишь эпизодически, но в результате также было отмечено смещение границ редин и редколесий в долине р. Тульйок (Хибины) на 27–29 м высоты с 1958 г. [10]. Это подтверждают разовые наблюдения сотрудников МГУ и Полярного геофизического института в долинах р. Юмьегорр (западный макросклон Хибин), которые на основе сравнения изображений растительности на исторической фотографии В. Рамзая (1890-е годы) сделали вывод о смещении верхней границы березового криволесья на 100–120 м высоты [11, 12]. Поэтому нами на склонах Хибин в экотоне верхней границы древесной растительности были проведены количественные исследования возраста, морфометрических параметров, роста и продуктивности как отдельных деревьев, так и целых древостоев для создания основы для разработки модели депонирования углерода в региональных лесных сообществах в ходе их климатогенных изменений.
ОБЪЕКТЫ И МЕТОДИКА ИССЛЕДОВАНИЙ
Район исследований – горный массив Хибины (наивысшая точка г. Юдычвумчорр, 1200.6 м над ур.м.) – расположен в полосе наиболее высоких центральных возвышенностей Кольского полуострова (рис. 1), для которого характерны значительные высоты (до 900–1200 м над ур. м.), их большие амплитуды (до 800 м), сложная геологическая структура, влажный климат (до 1200 мм в год), ярко выраженная высотная ландшафтная поясность [11]. Экотон верхней границы древесной растительности (ЭВГДР), под которым, согласно П.Л. Горчаковскому и С.Г. Шиятову [13], принято считать переходный пояс в горах между верхними границами распространения сомкнутых лесов и отдельных деревьев в тундре, в Хибинах приурочен к высотам от 300 до 600 м над ур. м. Произрастающие в его пределах древостои в основном сформированы березой извилистой (Betula pubescens ssp. tortuosa (Ledeb.) Nyman) и сосной обыкновенной (Pinus sylvesris L.) с примесью ели сибирской (Picea obovata Ledeb.).
Рис. 1.
Местоположение района исследований в пределах Российской Федерации (1), Кольского полуострова (2) и горного массива Хибины (3).
Значения координат левого верхнего и правого нижнего углов района интенсивных исследований на юго-восточном склоне Хибин с условным названием “Коашва” составляют 67.629° с.ш., 33.979° в.д. и 67.549° с.ш., 34.135° в.д. В ходе проведенных исследований в географической информационной системе ARC/INFO (ESRI Inc., США) с использованием алгоритма TOPOGRID была построена цифровая модель рельефа (ЦМР) данного района на основе геоинформационных слоев изолиний, отметки высот, водных потоков и озер (рис. 2). Данные слои были созданы в результате цифрования по сканированным изображениям листов карт “Госгисцентра” масштаба 1 : 25 000, полученных из общедоступного веб-ресурса loadmap.net. С использованием топографических карт 1956 и 1986 гг. и современных спутниковых снимков высокого пространственного разрешения, а также данных полевых обследований были созданы три линейных векторных геоинформационных слоя, которые характеризуют местоположение верхней границы леса в прошлом (1956 и 1986 гг.) и настоящем (2017 г.).
Рис. 2.
Верхняя граница редколесий сомкнутостью 35–40% на юго-восточном склоне г. Китчепах (Хибины) в 1956, 1986 и 2017 гг.
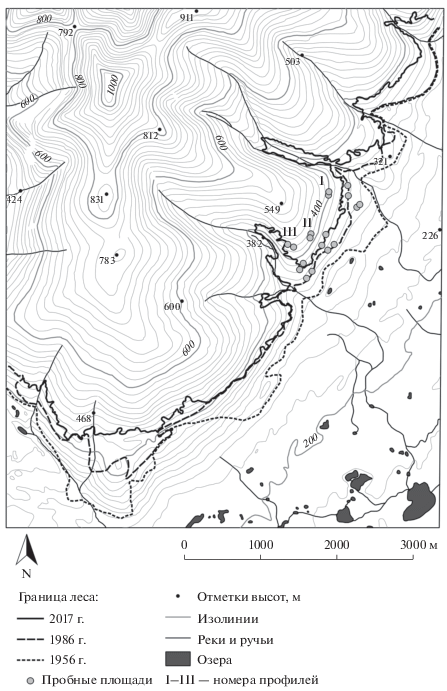
Величины вертикального и горизонтального сдвигов оценивали с использованием ранее разработанной авторами методики [14, 15]. Для получения значений высотного положения верхней границы леса предварительно растеризованная линия границы накладывается на цифровую модель рельефа. Каждой ячейке границы соответствует ячейка ЦМР размером 30 × 30 м с определенным значением высоты, а именно: 3302 ячейки – в 1956 г., 3322 – в 1986 г., 5260 – в 2017 г. Таким образом, высотное положение границы было охарактеризовано набором статистик распределения высот ячеек (табл. 1). Для оценки изменения величины высотного положения границы (высотного сдвига) использовали разность значений статистики, например медианы или среднего значения, высотного текущего положения верхней границы леса и ее положения в прошлом. Скорость высотного сдвига рассчитывали как отношение величины сдвига к временнóму интервалу.
Таблица 1.
Характеристики высотного положения и сдвига верхней границы редколесий сомкнутостью 35–40% на юго-восточном макросклоне Хибин (Кольский п-ов) за период с 1956 по 2017 г.
| Характеристики | Годы | Статистики | |||||
|---|---|---|---|---|---|---|---|
| среднее значение | стандартное отклонение | ошибка среднего | минимум | медиана | максимум | ||
| Высотное положение границы, м | 1956 | 304.5 | 39.2 | 0.7 | 249.1 | 293.1 | 397.9 |
| 1986 | 360.7 | 42.5 | 0.8 | 276.5 | 351.2 | 472.6 | |
| 2017 | 392.4 | 38.7 | 0.5 | 315.8 | 390 | 478.2 | |
| Высотный сдвиг, м/скорость сдвига, м/год | 1956–1986 | 56.2/1.9 | – | – | 27.4/0.9 | 58/1.9 | 74.7/2.5 |
| 1986–2017 | 31.7/1.0 | – | – | 5.6/0.2 | 38.8/1.3 | 39.4/1.3 | |
| 1956–2017 | 87.9/1.4 | – | – | 66.7/1.1 | 80.3/1.3 | 96.9/1.6 | |
| Горизонтальный сдвиг, м/скорость сдвига, м/год | 1956–1986 | 165.7/5.5 | 118.3 | 2.1 | – | 155/5.2 | 478.9/16 |
| 1986–2017 | 240.2/7.7 | 236.9 | 3.3 | – | 160.1/5.2 | 1036.2/33.4 | |
| 1956–2017 | 421.4/6.9 | 278.4 | 3.8 | – | 326.1/5.3 | 1413.2/23.2 | |
Для оценки величины и скорости горизонтального сдвига от линии границы в начале исследуемого периода рассчитывали растр, ячейки которого содержат значения евклидова расстояния. На него накладывается растеризованная линия верхней границы леса в конце исследуемого периода. Величина горизонтального сдвига была рассчитана с использованием статистик распределения его значений, а скорость – делением значений сдвига на величину временнóго интервала (см. табл. 1).
В июле 2017 г. для изучения структуры древостоев на трех высотных профилях на разных по локальной экспозиции участках (Ю, ЮВ, В) юго-восточного склона г. Китчепахк были заложены 18 пробных площадок размером 20 × 20 м – по 2 в нижней (320–340 м над ур. м.), средней (350–380 м над ур. м.) и верхней (410–450 м над ур. м.) частях экотона ВГДР (рис. 2, 3). На каждой пробной площади для отдельного дерева (одноствольной и многоствольной форм роста) определяли вид, местоположение, высоту ствола градуированным 6-метровым шестом и диаметр (у основания и на высоте груди) ствола через измерение периметра при помощи измерительной ленты, а также диаметр проекции кроны по двум направлениям и жизненное состояние.
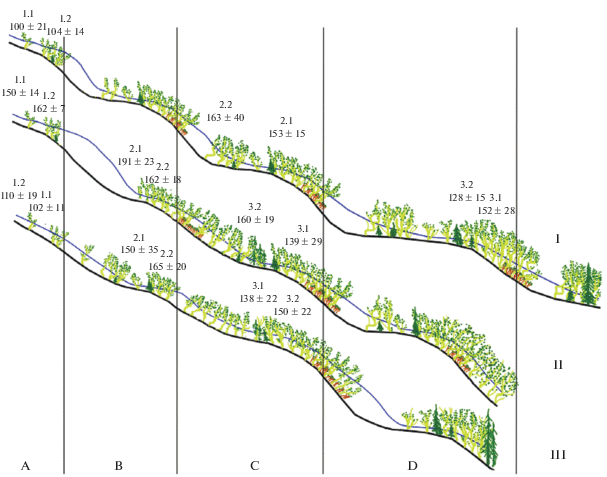
Рис. 3.
Схема распределения древостоев и снежных масс по высотному градиенту на исследованных профилях на юго-восточном склоне г. Китчепахк (Хибины): I–III – номера профилей; вертикальными линиями и буквами обозначены нагорные террасы (A–D); арабскими цифрами отмечены названия и места закладки пробных площадок и высота снега на них (см) в марте 2018 г. (средняя ± стандартное отклонение).
Для определения календарного времени появления живых деревьев (одноствольной формы роста) или стволов (многоствольной формы роста) диаметром >3–4 см брали буровые образцы древесины на высоте до 40 см, а у усохших выпиливали диск. У каждого второго молодого дерева высотой >0.2 м, но диаметром у основания ствола <3–4 см на уровне корневой шейки брали поперечный диск. В лабораторных условиях каждый поперечный диск и буровой образец (предварительно закрепленный в деревянном держателе) были зачищены режущим инструментом, а затем на зачищенную поверхность втирали зубной порошок для увеличения контрастности годичных колец. После этого у всех образцов древесины измеряли ширину годичных колец на установке LINTAB-III, а при помощи программного обеспечения TSAP–3.0 окончательно датировали год формирования самого ближнего к центру годичного кольца.
Если взятые керны не достигали центра ствола, то для определения времени формирования центрального кольца сначала определяли радиус дуги, образуемой самым ранним (ближним к центру) годичным кольцом, сравнивая его с линиями окружностей разного размера, нанесенных на прозрачную пленку. Затем подсчитывали количество годичных колец на самом раннем участке керна, равном вычисленному радиусу, и прибавляли к количеству выявленных и сдатированных годичных колец на образце [3]. Расчет поправки при помощи возрастной кривой нами не применялся, так как проведенные нами тестовые сравнения количества колец на 77 березовых кернах, достигших центра на отрезке от 0 до 2.5 мм (средний радиус дуги последнего полного кольца у не достигших центра 133 кернов) и от 2.6 до 5 мм не показали достоверных различий в количестве колец (6.3 ± 3.1 и 6.2 ± 2.9 мм соответственно).
Поскольку возраст подроста выше 0.2 см и диаметром <3–4 см был определен по спилам, взятым на уровне корневой шейки, то он был достоверным в наибольшей степени. Используя возраст таких деревьев и высоту их стволиков, было рассчитано уравнение регрессии зависимости между этими показателями, с помощью которого вычисляли поправки для определения более точного возраста каждого изученного нами дерева диаметром >3–4 см.
В соответствии с рядом распределения стволов елей и берез по диаметру на каждом высотном уровне в 2017 г. в непосредственной близости от заложенных пробных площадей формировали систематическую выборку модельных деревьев для определения таксационных показателей и фитомассы древостоев. Модельные деревья (10–30 шт.) отбирали средними по диаметру, высоте и размерам кроны для ступени толщины в пределах всего диапазона варьирования диаметров стволов в древостое.
Фитомасса была определена у 11 одноствольных елей и 53 стволов многоствольных особей березы с подразделением на следующие фракции: древесина и кора ствола, древесина и кора ветвей, хвоя или листья, отмершие ветви. Фитомассу стволов в коре определяли непосредственным взвешиванием в полевых условиях с точностью 50 г, для чего ствол распиливали на метровые секции. Содержание сухого вещества как в древесине, так и в коре определяли по дискам, выпиленным с торцов распиленных секций. Древесину и кору с дисков взвешивали на месте с точностью до 0.1 г, а затем отправляли в лабораторию для дальнейшего высушивания и измерения их массы в абсолютно сухом состоянии.
При определении фитомассы кроны и ее структурных частей все ветви со ствола были обрублены и разделены на три секции по расположению в кроне (верхняя, средняя и нижняя части). Общую массу каждой секции кроны измеряли непосредственным взвешиванием на месте в полевых условиях, после чего делили крону на охвоенную (олиствленную) и неохвоенную (неолиствленную) части. Из охвоенной (олиствленной) части отбирали навеску до 20–30% по весу от общей. Хвою (листья) в навеске отделяли от ветвей и взвешивали. Неохвоенные (неолиствленные) ветви взвешивали отдельно и из них брали навеску (5–10% от веса) для определения доли древесины и коры в ветвях. Для измерения абсолютно сухого вещества в хвое (листьях) из каждой секции кроны брали навеску весом 20 г для последующего высушивания в лабораторных условиях. Все навески высушивали в сушильном шкафу (ШСП-0.25–100) при температуре 106°С до абсолютно сухого состояния и взвешивали. Полученные соотношения между массой навесок в сыром (в полевых условиях) и абсолютно сухом состоянии использовали для вычисления фитомассы различных фракций модельных деревьев в абсолютно сухом состоянии. Для определения сухого вещества во всех фракциях взято 848 спилов и навесок и с использованием полученных весовых пропорций произведены расчеты фитомассы деревьев в абсолютно сухом состоянии.
Используя буровые образцы древесины (керны), для которых стандартными дендрохронологическими методами была измерена ширина каждого годичного кольца и определен календарный год их образования, рассчитали диаметры стволов, которые были у деревьев в каждый год их жизни. При расчетах считалось, что прирост ствола по отдельному радиусу (в месте изъятия керна) синхронен с приростом по осредненному диаметру ствола, рассчитанному через периметр его поверхности при помощи измерительной ленты. Расстояние по радиусу вычисляли путем сложения размеров всех годичных колец от сердцевины до годичного кольца, образовавшегося к определенному календарному году, включая последний [16–19].
Расчеты проводили по формуле Din year N = = (Lin year N/Lin 2017 year) × D measured in 2017 year, где Din year N – расчетные диаметры дерева в определенный календарный год N (например, 1935); Lin year N и Lin 2017 year – расстояние по керну от сердцевины до годичного кольца, образовавшегося в определенный календарный год N и конечный год (2017 г.) соответственно; D measured in 2017 year – фактический диаметр ствола в год измерения (2017 г.).
На основе фактических (2017 г.) и рассчитанных (для предшествующих лет роста деревьев) данных по диаметрам стволов отдельных деревьев на обследованных пробных площадках и выявленных тесных связей между надземной фитомассой деревьев и диаметрами их стволов были вычислены запасы надземной фитомассы на единицу площади и оценена скорость ее накопления для различных типов древостоя за последние 165 лет. Высоты деревьев были рассчитаны с использованием выявленных нами аллометрических зависимостей между высотой (y) и диаметром стволов (x), имеющих вид y = 13.43х0.67 и у = 17.46х0.78 для березы и ели соответственно.
С июля 2017 г. по июль 2018 г. для измерений температуры воздуха и поверхностей крон и стволов деревьев на высоте 2 м от поверхности земли в кроне (с защитой от прямых солнечных лучей и без) на каждой площадке было размещен автономный термодатчик (DS1921 Thermochron iButton™). В нижней части восточного профиля (Профиль I) и средней части южного профиля (Профиль III) были установлены по одной метеостанции, которые в этот же период производили измерения температуры и влажности воздуха, количества жидких осадков и ФАР (фотосинтетически активная радиация). В верхней, средней и нижней частях ЭВГДР рядом с площадками по линии сверху вниз через 5 м в почве на глубине 10 см были размещены термодатчики (DS1921 Thermochron iButton™). На основе полученных данных были вычислены средние температуры отдельных суток и месяцев, а также выявлены переходы через пороговые величины и даты схода снега.
В марте 2018 г. для оценки снегонакопления на 18 ранее изученных пробных площадях измеряли глубину снежного покрова: в верхней части ЭВГДР – при помощи 1.5-метрового шеста (по 30 на пробную площадь), а в средней и нижней – посредством покраски стволов всех деревьев на уровне снега и последующим измерением высоты меток в летнее время.
РЕЗУЛЬТАТЫ
Динамика верхней границы редколесий в последние 60 лет. Анализ современной высотной позиции верхней границы редколесий сомкнутостью 35–40% на участках разных экспозиций выявил для этого района Хибин увеличение ее средних величин начиная с северо-восточных (337 ± ± 28 м над ур. м.) к восточным (361 ± 13 м над ур. м.), юго-восточным (407 ± 35 м над ур. м.), южным (427 ± 17 м над ур. м.) и юго-западным (460 ± 36 м над ур. м.) склонам. Сравнение описанными выше методами современной высотной позиции верхней границы редколесий сомкнутостью 35–40% и отмеченной на топографических картах 1956 и 1986 гг. создания показало, что за последние 60 лет в этом районе произошло существенное продвижение древесной растительности выше в горы: в среднем на 88 м – по вертикали и 421 м – по горизонтали (см. табл. 1, рис. 2). При этом наиболее значительные сдвиги границ были выявлены на склонах ЮВ, Ю и ЮЗ экспозиций (в среднем 107, 105, 99 м высоты соответственно), средние – на В (в среднем 90 м) и минимальные – на СВ (в среднем 70 м).
Современная структура древостоев на исследованных профилях. При сравнении характеристик древостоев в пределах ЭВГДР (табл. 2) отмечается уменьшение в 1.2–1.8 раза средних значений морфометрических параметров деревьев по мере увеличения высоты над уровнем моря. При этом более заметно снижаются количество живых стволов березы высотой более 20 см и доля отмерших и поврежденных стволов в общей выборке. Доля стволов многоствольных берез в составе древостоев превышает в среднем 60%. Заметное участие ели в составе древостоев (20–60%) отмечается на отдельных площадках лишь в нижней части профилей (см. табл. 2), а на остальных уровнях ее доля не превышает нескольких процентов. Помимо ели и березы, в составе древостоев присутствует сосна обыкновенная (Pinis sylvestris L.), но ее доля в целом не более 0.1%.
Таблица 2.
Характеристика древостоев на обследованных площадках исследовательских профилей на юго-восточном склоне г. Китчепахк (Хибины) в 2017 г. (среднее значение ± стандартное отклонение)
| Высота над ур. м., м |
Сумма проекций крон, м2/га | Количество деревьев, шт./га | Диаметр ствола на высоте 1.3 м, см |
Высота ствола, м |
Возраст, лет | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| береза | ель | ||||||||||
| береза | ель | <1.5 м | >1.5 м | <0 м | в среднем | max | в среднем | max | в среднем | max | |
| Высотный профиль I | |||||||||||
| 407 | 372 | 0 | 325 | 300 | 0 | 4.7 ± 1.8 | 8.3 | 1.6 ± 0.6 | 2.6 | 51 ± 19 | 93 |
| 411 | 629 | 0 | 100 | 225 | 0 | 4.9 ± 3.7 | 11.8 | 1.8 ± 1.2 | 4.4 | 56 ± 18 | 94 |
| 345 | 6809 | 0 | 100 | 1100 | 0 | 7.5 ± 3.2 | 17.8 | 2.7 ± 0.8 | 3.9 | 88 ± 23 | 133 |
| 356 | 6094 | 0 | 350 | 1875 | 0 | 5.7 ± 0.7 | 16.9 | 2.2 ± 1.0 | 4.8 | 52 ± 19 | 119 |
| 322 | 6492 | 91 | 725 | 1500 | 75 | 6.4 ± 4.9 | 21.7 | 2.1 ± 1.1 | 5.5 | 55 ± 27 | 129 |
| 324 | 4519 | 833 | 575 | 3300 | 650 | 4.6 ± 2.7 | 17.5 | 1.9 ± 0.9 | 5.0 | 44 ± 12 | 101 |
| 9.6 ± 5.7 | 28.7 | 2.5 ± 1.8 | 6.8 | 49 ± 21 | 95 | ||||||
| Высотный профиль II | |||||||||||
| 442 | 515 | 73 | 250 | 250 | 100 | 4.9 ± 3.3 | 11.5 | 1.5 ± 0.7 | 2.6 | 49 ± 14 | 67 |
| 425 | 407 | 233 | 25 | 250 | 50 | 5.7 ± 1.9 | 9.1 | 1.8 ± 0.5 | 2.6 | 61 ± 18 | 94 |
| 381 | 4046 | 0 | 75 | 675 | 0 | 6.8 ± 3.2 | 14.3 | 2.1 ± 0.9 | 4.7 | 79 ± 29 | 122 |
| 360 | 5526 | 0 | 25 | 825 | 0 | 10.3 ± 4.2 | 20.9 | 2.4 ± 0.9 | 4.4 | 85 ± 36 | 194 |
| 333 | 6918 | 1 | 25 | 825 | 25 | 7.3 ± 3.2 | 15.3 | 2.5 ± 1.1 | 6.6 | 79 ± 27 | 121 |
| 335 | 2125 | 3187 | 25 | 400 | 850 | 8.6 ± 4.2 | 16.9 | 3.0 ± 1.2 | 5.2 | 85 ± 20 | 114 |
| 8.4 ± 4.9 | 34.5 | 2.8 ± 1.9 | 7.6 | 93 ± 41 | 201 | ||||||
| Высотный профиль III | |||||||||||
| 448 | 2589 | 79 | 125 | 475 | 25 | 4.5 ± 2.7 | 13.7 | 1.2 ± 0.5 | 2.3 | 62 ± 23 | 109 |
| 445 | 1638 | 0 | 0 | 300 | 0 | 8.8 ± 4.5 | 18.8 | 1.8 ± 0.5 | 2.5 | 92 ± 31 | 173 |
| 371 | 6718 | 0 | 100 | 1150 | 0 | 6.6 ± 2.9 | 12.7 | 2.4 ± 0.8 | 3.8 | 81 ± 25 | 135 |
| 379 | 6749 | 0 | 100 | 850 | 0 | 7.0 ± 3.6 | 15 | 2.3 ± 0.7 | 3.6 | 72 ± 30 | 124 |
| 339 | 5237 | 897 | 50 | 750 | 225 | 9.9 ± 5.5 | 29.6 | 3.0 ± 1.3 | 5.7 | 88 ± 28 | 146 |
| 8.1 ± 3.3 | 25.5 | 3.3 ± 2.0 | 7.0 | 97 ± 37 | 158 | ||||||
| 347 | 7101 | 411 | 0 | 1025 | 100 | 8.2 ± 4.6 | 22.3 | 2.7 ± 1.1 | 5.0 | 93 ± 36 | 161 |
Как видно из рис. 3, сомкнутость древостоев изменяется с высотой на исследованных профилях крайне неравномерно, что связано с явно выраженной террасированностью склонов, характерной для всего горного массива Хибин. Высота ступеней и ширина террас крайне вариабельны и изменяются от нескольких до десятков, а местами и сотни метров. Следует отметить, что в верхних частях ступеней нередко встречаются заросли искривленных осин (Populis tremula L.), имеющих высоту не более 2 м.
История формирования древостоев. Анализ данных о распределении деревьев березы по периодам появления на площадках Профиля I (рис. 4) показал, что в нижней части ЭВГДР (322–324 м над ур. м.) появление наиболее старых, ныне растущих деревьев датируется последним десятилетием XIX в., однако их количество очень мало (около 0.1%). Практически все деревья появились здесь в течение ХХ в. Наиболее активным семенное возобновление как березы, так и ели было между 1960 и 2000 гг. В период с 1905 г. по 1960 г. увеличению густоты древостоев на этом уровне способствовало появление новых вертикальных стволов у многоствольных деревьев. В средней части профиля (345–356 м над ур. м.) возраст самых старых живых деревьев также не превышает 135 лет. Основная масса деревьев появилась здесь, как и в нижней части, во второй половине XX в. Практически в течение всего периода заселения значительный вклад в увеличение густоты древостоев на этом уровне способствовало появление большого количества новых вертикальных побегов у многоствольных деревьев (более половины от общего количества). В верхней части профиля (407–411 м над ур. м.) деревья берез старше 95 лет не были обнаружены, а основная часть стволов появилась в период после 1970 г.
Рис. 4.
Возрастная структура древостоев на различных высотах исследованных профилей на юго-восточном склоне г. Китчепахк (Хибины) (количество стволов, появившихся в отдельные 5-летние интервалы (например, периоду между 1901 и 1905 гг. соответствует обозначение 1905)): 1– ель; 2 – основные (наиболее старые) стволы березы; 3 – дополнительные стволы березы.
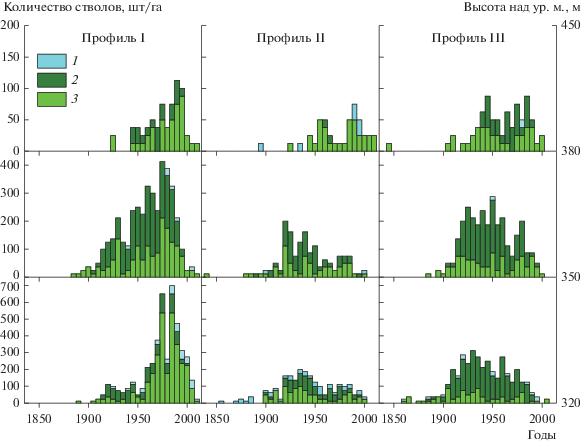
Анализ данных о распределении деревьев по периодам появления на площадках Профиля II показывает, что в нижней части этого профиля (333–335 м над ур. м.) возраст самых старых живых берез не превышает 125 лет, елей – 200 лет. Большинство деревьев берез появилось здесь в ХХ в., а наиболее активным возобновление было между 1900 и 1950 гг., затем оно снизилось в среднем в два раза. Увеличению густоты древостоев на этом уровне также способствовало появление значительного количества новых вертикальных побегов у многоствольных деревьев (≈40% от общего количества). На выровненных участках террас в составе древостоя ель более заметна – ее покрытие достигает местами 60%. Возобновление ели шло здесь параллельно с появлением берез в течение всего ХХ в. В средней части профиля (360–381 м над ур. м.) самые первые деревья берез из ныне растущих появились еще в первой половине XIХ в. Как и в нижней части, активное возобновление происходило между 1920 и 1950 гг., а также в 1960–1980 гг. В верхней части этого профиля (425–442 м над ур. м.) первые ныне растущие здесь березы стали появляться лишь в 1920-х годах, а активное ее заселение происходило после 1950 г. Ель стала появляться значительно раньше – в 40-х годах XIX в.
Анализ данных о распределении деревьев по периодам появления на площадках Профиля III показывает, что в его нижней части (339–347 м над ур. м.) возраст самых старых живых как берез, так елей около 160 лет. Наиболее активным возобновление березы было между 1905 и 1940 гг., а в последующий период оно заметно снизилось. На всем протяжении периода заселения этого уровня увеличению густоты древостоев способствовало появление новых вертикальных побегов у многоствольных деревьев (более 70% от общего количества). В средней части профиля (371–379 м над ур. м.) самые первые деревья из ныне растущих появились в конце XIX в., а периоды активного заселения были приурочены к 1905–1960 и 1970–1990 гг. Около 70% стволов, которые появились в период до 1960 г., принадлежат куртинам многоствольных деревьев. Позже увеличение густоты древостоев шло за счет появления как одноствольных, так и дополнительных стволов многоствольных деревьев в равной степени. В верхней части профиля (445–448 м над ур. м.) первые ныне растущие здесь березы стали появляться еще в 1840-х годах, но периоды активного заселения были приурочены к более позднему периоду, а именно к 1920–1960 и 1970–1990 гг.
Фитомасса модельных деревьев. В фракционной структуре фитомассы обоих изученных видов преобладает стволовая фитомасса, но ее вклад у ели и березы различается в 1.5–2 раза. Так, у ели на фитомассу ствола приходится в среднем 41% от общей надземной фитомассы деревьев, в то время как у березы этот показатель равен 68%. Доля ветвей в сложении надземной фитомассы у обоих видов не отличается и составляет в среднем 25–26%. Наибольшие различия заметны при сравнении фитомассы фотосинтезирующих органов: у березы фитомасса листьев составляет всего 5% от общей надземной фитомассы деревьев при 34%, приходящихся на долю фитомассы хвои ели.
На основе полученных результатов для каждого вида были разработаны уравнения связи массы различных фракций с единым морфометрическим показателем – диаметром у основания ствола. Анализ пригодности различных регрессионных уравнений показал, что наиболее точно данную связь описывает степенная функция вида y = = axb (рис. 5). Статистические показатели разработанных уравнений приведены в табл. 3. Связь между представленными показателями характеризуется как высокая и чрезвычайно высокая, что позволило применить данные уравнения для расчета фитомассы отдельных деревьев, произрастающих на пробных площадях, с дальнейшим переходом на уровень древостоев.
Рис. 5.
Зависимость надземной фитомассы деревьев ели и березы от их диаметра у основания ствола: 1 – ель, 2 – береза.
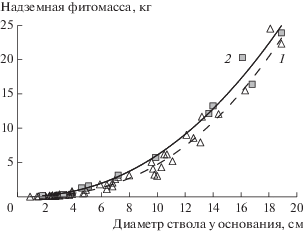
Таблица 3.
Характеристики уравнений зависимости фракций фитомассы (кг) стволов березы и деревьев ели от диаметра у основания ствола (см) на верхней границе редколесий на г. Китчепахк ( Хибины)
| Фитомасса | Вид | a | b | R2 | n |
|---|---|---|---|---|---|
| Общая надземная | Береза | 14.337 | 2.4939 | 0.9809 | 53 |
| Ель | 10.148 | 2.224 | 0.9881 | 11 | |
| Ствола | Береза | 13.533 | 2.6095 | 0.9737 | 53 |
| Ель | 7.9381 | 2.4946 | 0.9714 | 11 | |
| Ветвей | Береза | 1.0838 | 2.1377 | 0.8454 | 53 |
| Ель | 3.4208 | 2.3668 | 0.9877 | 11 | |
| Листьев или хвои | Береза | 0.0878 | 1.8612 | 0.8443 | 53 |
| Ель | 1.3212 | 1.8592 | 0.9633 | 11 |
Реконструкция таксационных характеристик и фитомассы древостоев. Анализ данных показал, что на всех высотных уровнях наблюдается постепенное увеличение средних диаметров и высот деревьев, однако скорость наблюдаемых изменений крайне низкая (табл. 4). На верхнем высотном уровне морфометрические характеристики деревьев за 137 лет возросли в среднем в 1.9–2 раза. В средней и нижней частях высотного профиля данные изменения более выражены – параметры деревьев увеличились от 3 до 7 раз. Также на среднем и нижнем высотных уровнях наблюдаются более высокие темпы роста густоты древостоев.
Таблица 4.
Реконструированные и фактические характеристики древостоев на разных типах верхних границ на юго-восточном склоне г. Китчепахк (Хибины)
| Характеристики | Верхняя граница | Годы | |||||||
|---|---|---|---|---|---|---|---|---|---|
| реконструированные | факти-ческие | ||||||||
| 1880 | 1910 | 1940 | 1955 | 1970 | 1985 | 2000 | 2017 | ||
| Средний диаметр ствола у основания, м | Редина | 0.02 | 0.03 | 0.01 | 0.01 | 0.02 | 0.02 | 0.03 | 0.04 |
| Редколесье | 0.03 | 0.02 | 0.01 | 0.02 | 0.03 | 0.04 | 0.05 | 0.07 | |
| Сомкнутый лес | 0.01 | 0.02 | 0.02 | 0.04 | 0.05 | 0.05 | 0.05 | 0.07 | |
| Средняя высота ствола, м | Редина | 0.85 | 1.14 | 0.75 | 0.86 | 0.96 | 1.065 | 1.17 | 1.66 |
| Редколесье | 1.29 | 0.97 | 0.76 | 0.99 | 1.21 | 1.475 | 1.74 | 2.18 | |
| Сомкнутый лес | 0.58 | 0.84 | 1.03 | 1.36 | 1.68 | 1.79 | 1.90 | 2.31 | |
| Количество стволов, шт./га | Редина | 4 | 4 | 37 | 148 | 258 | 450 | 642 | 667 |
| Редколесье | 8 | 50 | 612 | 1219 | 1825 | 2356 | 2887 | 2887 | |
| Сомкнутый лес | 29 | 112 | 875 | 917 | 958 | 2298 | 3637 | 3775 | |
| Надземная фитомасса, т/га | Редина | 0.0002 | 0.001 | 0.004 | 0.017 | 0.104 | 0.168 | 0.326 | 0.823 |
| Редколесье | 0.002 | 0.010 | 0.064 | 0.286 | 0.795 | 1.834 | 3.836 | 7.412 | |
| Сомкнутый лес | 0.001 | 0.015 | 0.247 | 1.062 | 2.879 | 6.048 | 10.67 | 17.82 | |
| Увеличение фитомассы в предшествующий период, т/га в год | Редина | 0.00001 | 0.00001 | 0.00002 | 0.0001 | 0.006 | 0.004 | 0.011 | 0.031 |
| Редколесье | 0.00007 | 0.00027 | 0.00062 | 0.0030 | 0.014 | 0.007 | 0.031 | 0.063 | |
| Сомкнутый лес | 0.00003 | 0.00046 | 0.00237 | 0.0131 | 0.023 | 0.014 | 0.053 | 0.096 | |
Оценка динамики накопления фитомассы показала, что на начальной стадии формирования древостоев в середине–второй половине ХIХ в. на всех высотных уровнях запас надземной фитомассы составлял в среднем 0.001 т/га (см. табл. 4). Средняя скорость накопления фитомассы в этот период была крайне низкой, достигая максимального значения 0.0004 т/га в год в редколесье. В древостоях на верхней границе сомкнутых лесов резкий подъем прироста фитомассы произошел в 1880–1910 гг. Годичная продукция в этот период возросла в 22 раза. В течение ХХ в. этот показатель увеличился в 280 раз, достигнув к 2000-му году 0.26 т/га в год. За первые 17 лет ХХI в. темпы прироста фитомассы возросли еще в 2 раза. Текущие запасы надземной фитомассы в сомкнутом лесу составляют 10.4– 28.5 т/га.
В редколесьях наблюдается схожая картина, но со сдвигом на 30 лет: до 1910 г. рост фитомассы древостоев происходил крайне медленно, в период 1910–1940 гг. последовало 17-кратное увеличение темпов накопления фитомассы, а к 1970 г. этот показатель вырос еще в 15 раз, достигнув 0.1 т/га в год. В последующие периоды темпы прироста фитомассы возросли еще в 9 раз. На текущий момент надземная фитомасса древостоев в редколесьях составляет 3.7–9.7 т/га.
В тундре с группами деревьев (рединах) в первой половине ХХ в. скорость накопления фитомассы оставалась крайне низкой. К 1940 г., когда фитомасса данных древостоев достигла 0.004 т/га, темпы ее прироста резко возросли: в период между 1940 и 1970 гг. этот показатель увеличился в 159 раз по сравнению с предыдущим периодом. В последующие годы здесь не наблюдается столь резких подъемов годичной продукции древостоев, хотя тенденция на увеличение темпов прироста фитомассы сохраняется. К 2017 г. запасы фитомассы древостоев в тундре с группами деревьев составили 0.5–1.7 т/га.
При анализе вклада различных видов древесных растений в общую фитомассу древостоев было выявлено, что на данный момент в ее составе доминирует береза, хотя ее вклад в течение ХХ в. на разных высотных уровнях менялся неодинаково: в сомкнутом лесу к началу ХХ в. 100% фитомассы древостоев составляла ель; в 1940-е годы произошло 4-кратное снижение вклада фитомассы ели в общую фитомассу древостоев, связанное с массовым появлением стволов березы; в последующие периоды в сомкнутом лесу доля фитомассы ели осталась практически неизменной, колеблясь в диапазоне от 23 до 25% от общих запасов фитомассы. В редколесье и тундре с группами деревьев наблюдается обратная ситуация: если в сложении фитомассы изначально преобладала береза, то ее вклад стал постепенно снижаться в 1970-е годы. Снижение вклада фитомассы березы более выражено на верхнем высотном уровне, где в различные периоды второй половины ХХ в. он составлял от 71 до 83%, в то время как в редколесье этот показатель не снижался ниже 96%.
Локальные условия мест произрастаний. Анализ данных термодатчиков показал, что средние месячные температуры воздуха в кронах берез в период с марта по октябрь на 0.4–0.8°С ниже в верхней части ЭВГДР по сравнению с нижней (табл. 5), а в декабре–феврале выше на 0.1–0.3°С или практически равны. Отмечено, что на одном и том же уровне автономными метеостанциями и термодатчиками, размещенными в густых кронах елей с защитой от прямых солнечных лучей, зафиксированы в марте–сентябре более низкие показатели (средние месячные температуры воздуха на 0.8–2.9°С), чем термодатчиками, расположенными в разреженных кронах берез без защиты, а в холодный период года они практически не отличались друг от друга. Это связано с тем, что термосенсоры метеостанций и термодатчиков в кронах елей, прикрытые от прямых солнечных лучей, показывают реальную температуру воздуха, а не прикрытые от солнца термодатчики – температуру поверхности ветвей и стволов деревьев, которая в теплый период года, как правило, в середине дня выше на 7–10°С.
Таблица 5.
Средняя температура воздуха и почвы в нижней части тундрового пояса и на верхних границах древостоев разной сомкнутости на юго-восточном склоне г. Китчепахк (Хибины) с 15 июля 2017 г. по 13 июля 2018 г.
| Год | Месяц | Температура воздуха, °С | Температура почвы, °С | |||||
|---|---|---|---|---|---|---|---|---|
| верхняя граница | горная тундра | верхняя граница | ||||||
| редины | редколесье | сомкну-тый лес | редины | редколесье | сомкну-тый лес | |||
| 2017 | Июль | 15.7 | 16.0 | 16.4 | 10.3 | 10.5 | 11.3 | 12.2 |
| 2017 | Август | 10.8 | 11.2 | 11.4 | 9.4 | 9.8 | 10.4 | 11.1 |
| 2017 | Сентябрь | 6.3 | 6.7 | 6.8 | 6.0 | 6.5 | 7.0 | 7.3 |
| 2017 | Октябрь | 0.5 | 0.9 | 1.2 | 2.6 | 3.1 | 3.6 | 3.5 |
| 2017 | Ноябрь | –3.7 | –3.5 | –3.3 | 0.6 | 1.0 | 1.4 | 1.2 |
| 2017 | Декабрь | –6.3 | –6.5 | –6.5 | 0.6 | 0.8 | 1.0 | 0.9 |
| 2018 | Январь | –7.6 | –7.5 | –7.4 | 0.5 | 0.6 | 0.8 | 0.7 |
| 2018 | Февраль | –8.9 | –9.0 | –8.8 | 0.3 | 0.4 | 0.6 | 0.6 |
| 2018 | Март | –8.7 | –8.2 | –8.1 | 0.2 | 0.2 | 0.4 | 0.2 |
| 2018 | Апрель | –0.3 | 0.4 | 0.6 | 0.1 | 0.2 | 0.3 | 0.3 |
| 2018 | Май | 7.9 | 8.5 | 8.3 | 0.4 | 1.9 | 2.0 | 3.5 |
| 2018 | Июнь | 9.6 | 10.3 | 10.4 | 6.1 | 7.1 | 7.9 | 8.8 |
| 2018 | Июль | 16.7 | 17.3 | 17.5 | 10.3 | 10.6 | 11.2 | 12.3 |
Результаты снегомерных работ в марте 2018 г. показали, что высота снежного покрова существенно варьирует при переходе от нижней к верхней части ЭВГДР на всех обследованных профилях – как в связи с явно выраженной террасированностью склонов, отмечаемой повсеместно на пологих и средних по крутизне участках Хибинских гор, так и в связи с перераспределением снега сильными горными ветрами. Так, при переходе от нижнего к среднему уровню средняя высота снега на обследованных площадках увеличивается с 128–160 до 150–191 см, а от среднего к верхнему снижается до 100–162 см (см. рис. 3). Это объясняется тем, что значительное количество снега сдувается с расположенных выше безлесных пространств и задерживается более сомкнутыми древостоями нижних уровней. Причем на среднем и нижнем уровнях снег задерживается сильнее, чем на верхнем, из-за большей густоты древостоев. Под крутыми склонами террас также накапливаются большие массы снега (до 3–5 м), которые обычно исчезают в начале июля. Например, летом 2017 г., с предшествующей ему многоснежной зимой и прохладной весной, такие снежники окончательно стаяли лишь в конце июля.
Сравнение данных термодатчиков, размещенных на глубине 10 см в почве, показало, что средние месячные температуры в мае–сентябре на 0.8–1.7°С ниже в верхней части ЭВГДР по сравнению с нижней, а в октябре–апреле различаются лишь на 0.1–0.4°С или равны (см. табл. 5). Анализ изменений температур почв показал, что в 2018 г. даты схода снега (идентифицируются по появлению резко выраженного суточного хода температур) между верхней и нижней частями ЭВГДР отличаются на 4 сут (19 и 15 мая соответственно), а между горными тундрами и нижней частью ЭВГДР – на 15 сут (1 июня и 15 мая соответственно). Также было выявлено, что в пределах ЭВГДР почва на глубине 10 см прогрелась, и ее температура стала выше 3.2°С (по мнению Дж. Паулсона и К. Кёрнера [20], корни древесных растений начинают функционировать лишь выше этой температуры) через 2–3 дня после схода снега, а в тундрах – через 8 дней. Наши расчеты показали, что к 30 июня 2018 г. сумма суточных температур почвы выше 3.2°С на верхней границе (ВГ) сомкнутых лесов была равна 364, на ВГ редколесий – 288, ВГ редин – 263, а в горной тундре – 171 град/дн.
ОБСУЖДЕНИЕ
Нами было выявлено, что в исследованном районе наибольшей высоты современная верхняя граница редколесий сомкнутостью 35–40% достигает на юго-западных склонах, где она выше, чем на северно-восточных, на 123 м. Также на склонах южных экспозиций в последние 60 лет наблюдается ее максимальный высотный сдвиг (99–107 м). Это объясняется хорошо известным фактом, что суммарная радиация (прямая + рассеянная), приходящая на открытую поверхность, на склонах различных экспозиций существенно отличается. На Хибинах (метеостанция Юкспор, 910 м над ур. м.) на южные склоны на поверхность с углом наклона 10° при реальном режиме облачности за июнь–август поступает 34.8 ккал/см2, на западные и восточные – 32.9, а северные – 30.6. С увеличением угла наклона поверхности разница между северными и южными склонами также возрастает: при 20° – до 6.5, при 30° – до 8.8, а при 50° – до 17.7 ккал/см2 [11]. Еще больше она возрастает при увеличении доли прямой радиации в общей (большом количестве часов солнечного сияния), и, как показывают наши наблюдения, прямые лучи солнца приводят к существенному нагреву крон и стволов деревьев, и их температура становится выше, чем температура окружающего воздуха, на 7–10° в середине дня и в среднем на 0.8–2.9°С за месяц.
Как отмечают Л. Куллман и Л. Оберг [9], в средней части Скандинавии с 1915 по 2007 г. на сравнительно многоснежных участках склонов верхняя граница редин (групп деревьев высотой более 2 м) с доминированием березы смещалась выше в горы по 0.74 ± 0.49 м/год, при этом в период с 1915 по 1975 г. оно было равно 0.75 ± 0.58 м/год, а с 1975 по 2007 г. – 0.65 ± 0.78 м/год. Для верховьев р. Тульйок в Хибинах было отмечено [10], что в период с 1958 по 2006 г. сдвиг верхней границы редин с доминированием березы составил 29 м высоты (0.6 м/год). В нашем районе исследований, на юго-восточном макросклоне Хибин, смещение верхней границы редколесий сомкнутостью 35–40% в период с 1956 по 2017 г. составляло в среднем 1.4 м/год и также наблюдалось различие по периодам наблюдений: 1.9 м/год – в 1956–1985 гг. и 1.0 м/год – в 1986–2017 гг. (см. табл. 1). Такие значительные различия скорости сдвига (в 2 раза) могут быть объяснены, на наш взгляд, тем, что деревьям “труднее” заселять безлесные тундры, чем “продвигаться” сомкнутыми сообществами за счет освоения пространства между уже существующими группами деревьев и островками леса, встречающимися выше верхней границы редколесий. Это объяснение подтверждает анализ возрастной структуры древостоев (см. рис. 4), большая часть которых в нижней и средней частях ЭВГДР начала формироваться еще в первой половине ХХ в., но ко времени создания топографических карт в 1956 г. были еще крайне разрежены и небольшими по высоте (по нашим расчетам 0.76–1 м) и поэтому отнесены к категории нелесных территорий. Через 30 лет (к 1986 г. – очередному году обновления информации на топографических картах) в составе древостоев появилось много новых индивидуумов, а кроны взрослых деревьев достаточно сильно разрослись, в том числе за счет появления порослевых побегов, многие из которых превратились в крупные дополнительные стволы. Это привело к резкому увеличению сомкнутости древостоев – выше 35–40%, в результате чего многие из них уже перешли в категорию лесов, и верхняя граница отмечаемых на картах лесных сообществ стала проходить значительно выше. В последующий период, до 2017 г., процессы появления новых индивидуумов и увеличения размера крон в нижней и средней частях ЭВГДР стали постепенно замедляться из-за возрастания конкуренции и, видимо, это привело к почти двукратному снижению темпов поднятия верхней границы сомкнутых лесов, хотя процессы заселения горных тундр не прекратились, но шли менее быстро. Возможно, при дальнейшем улучшении условий для роста и возобновления деревьев поколение, появившееся в 1980–2000 гг., также быстро займет пространство, и граница сомкнутых древостоев в ближайшие годы заметно продвинется выше в горы.
Сравнение данных по запасам фитомассы древостоев в пределах ЭВГДР для г. Китчепахк (Хибины) и полученных нами ранее для многоснежных районов Урала с доминированием березы показало, что они ниже в 1.5–3 раза на одних тех же типах верхних границ. Так, в Хибинах (67.6° с.ш.), в нашем районе исследований, на верхней границе (ВГ) сомкнутых лесов запасы фитомассы в среднем равны 17.8 т/га, на ВГ редколесий – 7.4 т/га, а ВГ редин – 0.8 т/га, а на южном склоне г. Конжаковский Камень (Северный Урал, 59.63° с.ш.) они составляют 59.5, 36.4 и 3.1 т/га соответственно [21]. В более северных районах Северного Урала, на склонах гор в районе хр. Молебный Камень (61.21° с.ш.), они равны 38.0, 21.6 и 2.3 т/га, а на г. Ярута (63.34° с.ш.) – 28.5, 7.3 и 0.7 т/га соответственно (не опубликованные данные). Мы предполагаем, что наблюдаемое снижение запасов фитомассы по мере продвижения с юга на север связано с общим широтным уменьшением длительности периода вегетации (на 7–10 дней), хотя основным фактором, определяющим высотную позицию ЭВГДР в каждом регионе, остается температурный режим летних месяцев [20]. Поскольку Хибины имеют наиболее северное географическое положение, запасы фитомассы здесь значительно ниже по сравнению с другими районами.
Наблюдаемая в течение последнего столетия экспансия древесной растительности в Хибинских горах на склонах разной экспозиции, крутизны, каменистости, увлажнения и снежности свидетельствует о влиянии на эти процессы общих для всей территории факторов, которыми, по нашему мнению, могут быть только климатические. Анализ данных гидрометеостанции Кандалакша, расположенной в 67 км к юго-западу и имеющей наиболее длинный непрерывный ряд наблюдений на Кольском полуострове, показал, что в период с начала XX в. по начало XXI в. средние температуры воздуха в марте–июне выросли на 0.9–1.3°С, в июле, сентябре и октябре – на 0.2, 0.3 и 0.7°С, в августе снизились на 0.3°С, в результате в мае–сентябре увеличение в среднем составило 0.5°С. В ноябре–феврале температуры воздуха отдельных месяцев снизились на 0.2–1.4°С, что привело в целом к уменьшению средней температуры этого период года на 0.9°С. По данным метеостанции, расположенной на г. Ловчорр (1091 м над ур. м.) в Хибинах рядом с г. Кировск, средние многолетние температуры в 1991–2010 гг. стали больше, чем в 1961–1990 гг., на 0.9°С в ноябре–марте, на 0.4°С – в апреле–мае, на 0.6°С – в июне–августе и на 0.7°С – в сентябре–октябре [12].
Как показано выше, в течение последних 100 лет наблюдалось потепление, особенно в начале и конце вегетационного периода (в мае–июне и сентябре). Это привело к заметному увеличению продолжительности вегетационного периода, особенно в связи с его более ранним началом, что крайне важно для развития древесных растений в период их интенсивного роста. Так, только между 1961–1990 и 1991–2010 гг. произошло увеличение периодов с температурами выше 5 и 10°С на 4 и 9 дней соответственно [12]. Что касается режима увлажнения, то небольшое снижение среднегодовой суммы осадков наблюдалось на Кольском п-ове с начала XX в. до середины 1940-х годов, а в 1950–1960-е годы она достигала наибольших значений, после чего произошел спад к началу 1990-х годов, а в последние десятилетия отмечается их небольшой рост. Как отмечает В.И. Демин [12], в целом заметен лишь очень небольшой вековой тренд на повышение сумм осадков.
Общий анализ изменения локальных условий с высотой над уровнем моря показал (см. табл. 5 и рис. 3), что при переходе от границы сомкнутых лесов к тундрам (всего 80–120 м высоты) температура воздуха снижается на 0.4–0.8°С, а почвы – на 0.8–1.7°С. В результате продолжительность вегетационного периода уменьшается на 4 суток, а прогрев почв и их теплообеспеченность (сумма суточных температур почвы выше 3.2°С) в начале сезона вегетации становятся меньше почти в 1.5–2 раза. Поэтому даже относительно незначительное увеличение средних температур воздуха (0.9–1.3°С) в последнем столетии и сдвиг сроков начала вегетации (около 7 дней) на более ранние даты могли способствовать заметному ускорению роста и возобновления древесных видов и поднятию высотных рубежей сомкнутых древостоев. Это подтверждают выявленные нами тесные корреляционные связи между количеством берез, появлявшихся в верхней, сравнительно немногоснежной (109 ± 24 см) части ЭВГДР, и средними месячными температурами мая–июня по пятилетиям в течение всего периода между 1911 и 1995 гг. (R2 = 0.46). Для средней и нижней более многоснежных (160 ± 32 и 141 ± 24 см соответственно) частей ЭВГДР в период с 1911 по 1935 г., когда среднегодовые суммы осадков были близки к средним значениям, отмечаются наиболее сильные связи между количеством появлявшихся берез и температурами июня (R2 = 0.76); в период с 1941 по 1965 г., когда среднегодовые суммы осадков достигали максимумов в XX в. [12], – со средними температурами мая–июля (R2 = 0.61), а в период с 1966 по 1995 г., когда наблюдалось снижение увлажнения, – со средними температурами июля–сентября (R2 = 0.78). В последние 20–25 лет связи были крайне слабыми с температурами любых летних месяце, что объясняется снижением уровня лесовозобновления в нижних частях ЭВГДР из-за возрастания конкуренции по мере увеличениям сомкнутости древостоев.
Исследования американских коллег на тихоокеанском побережье северо-запада США в районах, где выпадает сравнительно большое количество осадков, выявили [22, 23], что на фоне общего повышения температуры летних месяцев расселение деревьев по субальпийским лугам приурочено к периодам снижения высоты снежного покрова, в течение которых происходит более раннее стаивание снега, в результате чего увеличивается продолжительность вегетационного сезона. В горных районах США, где орографические условия определяют различную степень увлажнения склонов и локальных местообитаний, интенсивное возобновление наблюдается в разные или перекрывающиеся периоды, отличающиеся по степени увлажнения [24, 25]. Важность изучения заснеженности склонов для динамики древостоев также подтверждают наблюдения за распределением снега и датами его исчезновения на обследованных участках склонов Хибин – в местах накопления больших масс снега и поэтому более длительным его залеганием сомкнутость древостоев даже в нижней части ЭВГДР крайне низкая или древесная растительность вообще отсутствует (см. рис. 3). Это хорошо заметно при рассмотрении современных спутниковых снимков территории Хибин в программе GoogleEarth, на которых отчетливо проявляется большая неоднородность в распределении древостоев в пределах переходной полосы между сомкнутыми лесами и горными тундрами.
Таким образом, наши исследования показали, что в последние столетия на фоне повышения на Кольском полуострове ранних летних (на 0.9–1.3°С) температур и более раннего начала вегетационного сезона существенно изменились структура и продуктивность древостоев на верхнем пределе их произрастания. Верхняя граница древостоев сомкнутостью 35–40% поднялась за последние 60 лет в среднем на 88 м высоты, а надземная фитомасса древостоев на трех обследованных профилях на юго-восточном склоне г. Китчепахк (Хибины) увеличилась с начала XX в. в среднем с 0.01 до 17.8 т/га на высоте 320–340 м над ур. м. (современная верхняя граница сомкнутых лесов), до 7.4 т/га – на высоте 350–380 м над ур. м. (верхняя граница редколесий) и до 0.8 т/га – на высоте 410–450 м над ур. м. (верхняя граница групп деревьев в тундре). Полученные в ходе исследований результаты существенно уточняют общую картину депонирования углерода в региональных лесных сообществах и способствуют разработке количественной модели, имитирующей реакцию горных экосистем региона в ответ на будущие изменения климата.
Исследование выполнено за счет Российского научного фонда (проект № 17-14-01112).
Список литературы
Harsch M.A., Hulme P.E., McGlone M.S. et al. Are treelines advancing? A global meta-analysis of treeline response to climate warming // Ecol. Lett. 2009. V. 12. № 10. P. 1040–1049.
Holtmeier F.-K. Mountain timberlines: Ecology, patchiness, and dynamics. Berlin: Springer, 2009. 437 p.
Moiseev P.A., Bubnov M.O., Devi N.M. et al. Changes in the structure and phytomass of tree stands at the upper limit of their growth in the Southern Urals // Rus. J. Ecol. 2016. V. 47. № 3. P. 219–227.
Grafius D.R., Malanson G.P. Biomass distributions in dwarf tree, krummholz, and tundra vegetation in the alpine treeline ecotone // Phys. Geogr. 2015. V. 36. № 4. P. 337–352.
Xin L., Yu N., Gao-qiang K. et al. Contrasting changes in above- and below ground biomass allocation across treeline ecotones in southeast Tibet // J. Mountain Sci. 2016. V. 13. № 2. P. 2036–2045.
Kullman L. Tree limit dynamics of Betula pubescens ssp. tortuosa in relation to climate variability: evidence from central Sweden // J. Veg. Sci. 1993. V. 4. № 6. P. 765–772.
Kullman L., Engelmark O. Neoglacial climate control of subarctic Picea abies stand dynamics and range limit in Northern Sweden // Arct. Alp. Res. 1997. V. 29. № 3. P. 315–326.
Hofgaard A., Dalen L., Hytteborn H. Tree recruitment above the treeline and potential for climate-driven treeline change // J. Veg. Sci. 2009. V. 20. № 6. P. 1133–1144.
Kullman L., Öberg L. Post-Little Ice Age tree line rise and climate warming in the Swedish Scandes: A landscape ecological perspective // J. Ecol. 2009. V. 97. № 3. P. 415–429.
Mathisen I.E., Mikheeva A., Tutubalina O.V. et al. Fifty years of tree line change in the Khibiny Mountains, Russia: Advantages of combined remote sensing and dendroecological approaches // Appl. Veg. Sci. 2014. V. 17. № 1. P. 6–16.
Природные уcловия Хибинского горного массива: Учеб. пос. / Ред. Мягков С.М. М.: Изд-во МГУ, 1986. 170 c.
Демин В.И. Основные климатические тенденции на Кольском полуострове за период инструментальных метеорологических измерений // Труды Кольского научного центра РАН. 2012. T. 3. № 10. C. 98–110.
Горчаковский П.Л., Шиятов С.Г. Фитоиндикация условий среды и природных процессов в высокогорьях. М.: Наука, 1985. 208 c.
Shiyatov S.G., Terent’ev M.M., Fomin V.V. et al. Altitudinal and horizontal shifts of the upper boundaries of open and closed forests in the Polar Urals in the 20th century // Rus. J. Ecol. 2007. V. 38. № 4. P. 223–227.
Фомин В.В. Климатогенная и антропогенная пространственно-временная динамика древесной растительности во второй половине XX века. Екатеринбург: ИЭРиЖ УрО РАН, 2009. 150 c.
Bakker J.D. A new, proportional method for reconstructing historical tree diameters // Can. J. For. Res. 2005. V. 35. № 10. P. 2515–2520.
Poulter B., Bouriaud O., Szejner P. et al. A tree-ring perspective on the terrestrial carbon cycle // Oecologia. 2014. V. 176. № 2. P. 307–322.
Babst F., Bouriaud O., Papale D. et al. Above-ground woody carbon sequestration measured from tree rings is coherent with net ecosystem productivity at five eddy-covariance sites // New Phytol. 2013. V. 201. № 4. P. 1289–1303.
Babst F., Bouriaud O., Alexander R. et al. Toward consistent measurements of carbon accumulation: A multi-site assessment of biomass and basal area increment across Europe // Dendrochronologia. Elsevier GmbH., 2014. V. 32. № 2. P. 153–161.
Paulsen J., Körner C. A climate-based model to predict potential treeline position around the globe // Alp. Bot. 2014. V. 124. № 1. P. 1–12.
Нагимов З.Я., Моисеев П.А., Бартыш А.А. и др. Структура и фитомасса березовых древостоев на верхней границе леса в условиях Северного Урала // Лесной вестник. 2008. Т. 3. С. 61–67.
Franklin J.F., Moir W.H., Douglas G.W. et al. Invasion of subalpine meadows by trees in the Cascade Range, Washington and Oregon // Arct. Alp. Res. 1971. V. 3. № 3. P. 215–224.
Taylor A.H. Forest expansion and climate change in the Mountain Hemlock (Tsuga mertensiana) Zone, Lassen Volcanic National Park, California, U.S.A. // Arct. Alp. Res. 1995. V. 27. № 3. P. 207.
Rochefort R.M., Peterson D.L. Temporal and spatial distribution of trees in subalpine meadows of Mount Rainier National Park, Washington, U.S.A. // Arct. Alp. Res. 1996. V. 28. № 1. P. 52–59.
Woodward A., Schreiner E.G., Silsbee D.G. Climate, geography, and tree establishment in subalpine meadows of the Olympic Mountains, Washington, U.S.A. // Arct. Alp. Res. 1995. V. 27. № 3. P. 217–225.
Дополнительные материалы отсутствуют.