

Экология, 2019, № 5, стр. 387-391
Основные и сопутствующие жертвы филина (Bubo bubo) в задачах исторической экологии
Н. Г. Смирнов a, *, Ю. Э. Кропачева a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: nsmirnov@ipae.uran.ru
Поступила в редакцию 04.03.2019
После доработки 10.04.2019
Принята к публикации 20.04.2019
Аннотация
Рассмотрены массовые материалы по питанию филина из трех широтных участков Урала (северная и южная тайга, степь). Все сборы проведены в однотипных местонахождениях костных остатков жертв из погадок на местах гнездования филинов в нишах и гротах на скалах по берегам рек. Жертвы филина разделены на три категории: основные, альтернативные и сопутствующие. Показана степень соответствия долей особей разных видов и их групп в питании филина и населении грызунов окрестностей гнездования. Подчеркивается важность отдельного рассмотрения основных и прочих жертв филина при реконструкции состава фауны и структуры населения грызунов по субфоссильным материалам орнитогенного происхождения.
Историческая экология, или история биогеоценозов в голоцене, интенсивно развивается в последние десятилетия с опорой на основы, заложенные в отечественной науке благодаря научным школам В.Н. Сукачева, Л.Г. Динесмана, И.М. Громова. Особенно важную роль в этом развитии играла лаборатория исторической экологии, основанная в ИЭМЭЖ АН СССР Л.Г. Динесманом, которому в 2019 г. исполнилось бы 100 лет.
В основе решения ряда задач исторической экологии находятся данные по массовым скоплениям костных остатков мелких млекопитающих, образовавшихся за счет питания хищных птиц. Еще несколько десятков лет назад в ряде районов, и в частности на Урале, филин (Bubo bubo Linnaeus, 1758) был одним из основных накопителей такого материала. Гнезда и присады этого хищника располагались в труднодоступных местах на скальных берегах рек в навесах, гротах. Там, перемешиваясь с продуктами физического и химического разрушения карстующихся пород, кости из погадок проходят начальные этапы фоссилизации и образуют костеносные горизонты [1, 2]. Повсеместное сокращение численности филина быстро приводит к тому, что эти источники субфоссильного материала становятся все более редкими, но некогда жилые гнезда еще доступны для изучения палеонтологическими методами.
Особенности питания филина в орнитологической литературе рассмотрены с разных сторон и весьма подробно [3–5]. Имея обширный ареал с громадным разнообразием условий обитания и широким набором потенциальных жертв, этот вид по разнообразию кормов относится к категории универсальных хищников. Отмечается, что в некоторых особых случаях ему нет необходимости “отвлекаться” на ловлю разной добычи, когда в изобилии присутствуют привлекательные и легкодоступные жертвы какого-то одного вида [6]. Подобная ситуация встречается нечасто. Как правило, филины облавливают территорию так, что в списке их добычи за несколько лет оказываются практически все виды грызунов доступных размеров [3–5, 7]. Кроме того, в пищевой спектр этих хищников входят птицы, но их анализ остается за рамками данной работы.
Основное внимание здесь будет уделено той части списка жертв, который обычно не привлекает внимания орнитологов. В наши задачи входит сравнительный анализ не только основной, но и сопутствующей добычи филинов разных широтных участков Урала. Она включает виды, составляющие по массе далеко не главную часть. Хищники добывают сопутствующих жертв не только на охотничьих, но и на транзитных территориях. Именно эти виды выступают в качестве индикаторов присутствия в окружающем ландшафте таких местообитаний, которые важны для палеореконструкций, но являются второстепенными для кормодобывающей деятельности хищников.
МАТЕРИАЛ И МЕТОДЫ
Данная работа основана на опубликованном материале, собранном в разных широтных районах Урала: северная тайга Печоро-Илычского заповедника [8], южная тайга на границе с лесостепью на Среднем Урале [9], степь южной оконечности Уральского хребта [10]. Костеносные слои в каждом из исследованных гнезд имеют разную мощность и накоплены за тот или иной хронологический промежуток в пределах голоцена. В некоторых из них филины периодически продолжают гнездиться и в настоящее время. Сведения о том, что некоторые гнезда какое-то время назад принадлежали филинам, получены от работников природоохранных структур соответствующих территорий. Некоторые гнезда и присады не несут следов недавнего гнездования, но никакой другой хищник, кроме филина, не мог заселять эти места. Поверхностная часть отложений в таких навесах и гротах содержит то или иное количество костных остатков мелких животных с явными признаками погадочного происхождения [11, 12]. После промывки грунта в нем обнаруживаются фрагменты скорлупы яиц филинов. Раскопки и камеральную обработку остеологических материалов проводили по стандартным для палеонтологии методикам. Количество особей жертв оценивали по одноименным костным остаткам (зубам).
Отнесение видов к основным или сопутствующим жертвам осуществляли на основе доли особей соответствующих видов в добыче. Строгой количественной границы между основными, альтернативными и сопутствующими жертвами существовать не может, так как это подвижная характеристика, которая определяется структурой доминирования жертв в рационе. Категории жертв выделяются на основе анализа питания хищника за ряд лет, включающий разные фазы динамики численности добычи [13, 14]. При анализе субфоссильного и ископаемого материала такого рода информация не может быть получена, поскольку данный материал представляет собой остатки добычи, накопленной за время, измеряемое десятками и сотнями лет.
На основании сравнительных данных по рецентным материалам [3–7] основными жертвами мы считаем виды, доминирующие в орнитогенных отложениях. Большая часть списка добычи принадлежит сопутствующим жертвам, но доля, приходящаяся на каждый из этих видов, незначительна. Наиболее сложно выделить в орнитогенном материале, накопленном за многолетний промежуток времени, альтернативные жертвы, так как эти виды достигают существенной доли в питании периодически. Количество таких жертв в усредненном рационе промежуточное между основными и сопутствующими жертвами. Массу жертв высчитывали по данным, приведенным для грызунов в справочниках и определителях [15].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
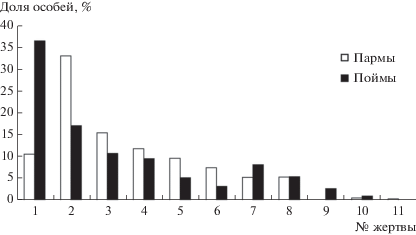
В каждом из широтных участков Урала основные жертвы разные в зависимости от биотопов, которые облавливает хищник. В таежных условиях верхнего течения р. Печоры, как правило, это пойменные луга, где основная добыча – водяная полевка. При малой площади лугов на лесных участках (пармы) филин добывает белок (рис. 1). Доля этих видов по массе составляет в пойменных местообитаниях соответственно 50 и 36%, в пармах – 14 и 65%. Вне зависимости от основной добычи в списке жертв филина присутствуют почти все виды наземных грызунов, попадающих по размерам в жертвы филина. Доля каждого из них по количеству пойманных особей варьирует от первых единиц до 10%, а по массе существенно меньше. Суммарно на остальные девять видов жертв приходится 50% особей и 18% массы (это в среднем 6 и 2% на каждый вид соответственно). Перечень видов дает ясное представление о фауне района – это обитатели лесных и околоводных биотопов. По данным отловов [16], ядро сообщества грызунов здесь составляют красная и рыжая полевки – многочисленные виды с относительно стабильной динамикой обилия. В годы подъемов обилия мелких млекопитающих отмечается высокая численность темной полевки, полевки-экономки и лесного лемминга, в годы депрессий – почти полное их отсутствие [16]. Виды, составляющие основу сообщества грызунов по данным отловов – красная и рыжая полевки, в добыче филина составляли в среднем по 5% и должны быть отнесены к категории сопутствующих жертв. Значительные колебания численности полевки-экономки и лесного лемминга и относительно высокие доли этих видов в питании филина позволяют предположить, что они могут играть роль альтернативных жертв. Вероятно, к этой же категории принадлежит и красно-серая полевка.
Рис. 1.
Доли особей жертв в питании филина в пойме р. Печоры и на таежных участках (пармы) Печоро-Илычского заповедника (Северный Урал) по [8]: 1 – Arvicola terrestris, 2 – Sciurus vulgaris, 3 – Microtus oeconomus, 4 – Clethrionomys rufocanus, 5 – Myopus schisticolor, 6 – M. agrestis, 7 – Cl. glareolus, 8 – Cl. rutilus, 9 – Pteromys volans, 10 – Eutamias sibiricus, 11 – Ondatra zibetica.
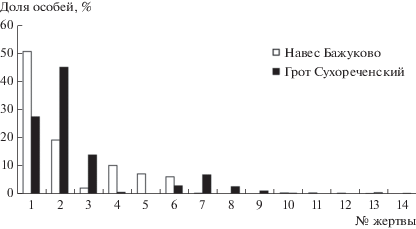
На Среднем Урале к основным жертвам филина по количеству добываемых особей можно отнести два вида – водяную и обыкновенную полевок (рис. 2): первый вид (обитатель поймы) резко преобладал в добыче обитателей навеса Бажуково, второй по доле особей в добыче была обыкновенная полевка, населяющая луга и сельхозугодия. В гроте Сухореченском соотношение долей этих видов было обратным. Это легко объясняется разницей в окружающих гнезда биотопах. В непосредственной близости от Сухореченского грота, помимо поймы р. Серги, находятся обширные пространства полей и земель разнообразного вида использования для нужд сельского хозяйства на месте островной Красноуфимской лесостепи. Здесь в населении мелких грызунов доминантом является обыкновенная полевка. На другом берегу реки находится Уфимское плато, покрытое лесами южной тайги. По массе добычи филина в Сухореченском гроте преобладает обыкновенный хомяк (55%), но и водяная полевка составляет значительную долю массы (30%). Последняя по массе абсолютно доминирует среди жертв обитателей навеса Бажуково (76%). По доле добытых особей остальные жертвы (11 таксонов) в этих местонахождениях составляют 19%, по массе – 5.5% (в среднем 2 и 0.5% на каждый вид соответственно).
Рис. 2.
Доли особей жертв в питании филина на Среднем Урале по [9]: 1 – A. terrestris, 2 – M. ex gr. arvalis, 3 – Cricetus cricetus, 4 – M. oeconomus, 5 – Clethrionomys sp., 6 – M. agrestis, 7 – Apodemus sp., 8 – Cl. glareolus, 9 – Cl. rufocanus, 10 – Sciurus vulgaris, 11 – Sicista betulina, 12 – M. schisticolor, 13 – P. volans, 14 – Rattus norvegicus.
Отловы грызунов живоловушками и конусами в разнообразных биотопах поймы р. Серги показали [17], что здесь преобладали рыжая полевка и экономка. Если вычислить средние значения долей основных видов в восьми биотопах, где грызунов ловили разными орудиями, то их последовательность по убыванию будет выглядеть так: рыжая полевка (38%), экономка (20%), малая лесная мышь (11%), лесная мышовка (9%), темная полевка (8%). Разумеется, результаты разовых отловов трудно сопоставлять с данными добычи филина за многие годы, но приведенные материалы дают удовлетворительное представление о населении пойменных участков. Эти данные можно сопоставить с данными по навесу Бажуково. Видовой состав в этих выборках совпадает, за исключением тех грызунов, которые не могли быть пойманы живоловушками и конусами, но оказались в добыче филина. Особое место в отловах принадлежит доминирующей там рыжей полевке, которая в добыче филина составляла всего несколько процентов и, безусловно, должна быть отнесена к категории сопутствующих жертв, как и 10 прочих видов с малыми долями добытых хищником особей. Единственный вид – полевка-экономка – в пище филина занимала более заметное место (около 10%). Это дает основание считать ее альтернативной жертвой. Не противоречит такому предположению и заметная доля экономки в отловах.
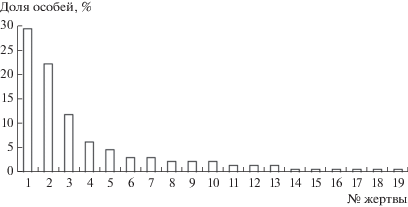
В степи южной оконечности Урала в гротах Верблюжка 1 и Верблюжка 2 основную добычу филинов по количеству особей образуют слепушонки, обыкновенный хомяк и обыкновенная полевка (рис. 3), но по массе резко преобладает хомяк (71%). На долю остальных 16 видов по количеству добытых особей приходится 36%, по массе – 22% (в среднем 2 и 1% на каждый вид соответственно). Естественное население мелких млекопитающих после освоения целинных и залежных земель в середине XX в. претерпело глубокую антропогенную трансформацию [18]. Повсюду, где преобладают агроландшафты, в сообществах мелких млекопитающих доминирует обыкновенная полевка. В поймах с древесно-кустарниковой растительностью массовым видом является рыжая полевка [19, 20].
Рис. 3.
Доли особей жертв в питании филина на Южном Урале по [10]: 1 – Ellobius talpinus, 2 – Cricetus cricetus, 3 – M. ex gr. arvalis, 4 – Ochotona pusilla, 5 – A. terrestris, 6 – Allactaga major, 7 – Clethrionomys rutilus, 8 – Allocricetulus eversmanni, 9 – M. gregalis, 10 – Mus musculus, 11 – A. ex gr. uralensis-agrarius, 12 – Cl. glareolus, 13 – Cricetulus migratorius, 14 – M. agrestis, 15 – Sylvaemus uralensis, 16 – Rattus sp., 17 – Spermophilus cf. pygmaeus, 18 – Sicista sp., 19 – Dipus sp.
Существенные изменения в структуре населения мелких млекопитающих продолжаются и в последние десятилетия. К сожалению, нам не удалось подобрать рецентный материал для корректного сравнения с усредненным рационом филина из отложений Верблюжки. Его можно использовать для сопоставления с основными и сопутствующими жертвами из других районов Урала. Такое сравнение показывает зональные особенности добычи филина в степных, пойменных и других биотопах, хорошо отражающие современный этап развития региональных сообществ грызунов [10].
ВЫВОДЫ
1. Во всех широтных участках Урала состав добычи филина хорошо отражает состав фауны грызунов.
2. Основные жертвы филина могут принадлежать к зональным комплексам или группам других категорий. В северной, южной тайге и степи в качестве основных жертв филина выявлены следующие виды грызунов соответственно: белка и водяная полевка; обыкновенная, водяная полевки и обыкновенный хомяк; слепушонка, обыкновенный хомяк. По доле добытых особей основные жертвы на Северном, Среднем и Южном Урале соответственно составляют в среднем 48, 70 и 52%. Только в северной тайге среди основных жертв филина имеется обитатель лесных биотопов – белка. Все прочие виды основных жертв обитают в открытых и околоводных биотопах, и их состав отражает избирательные предпочтения филина по выбору охотничьих угодий.
3. В список сопутствующих жертв соответствующих районов вошло следующее количество видов: 9 (северная тайга), 12 (южная тайга), 16 (степь). Доли особей отражаются в структуре сопутствующей добычи филина не пропорционально численности в природе, а избирательно в соответствии с частотой посещения хищником соответствующего биотопа. Возможны ситуации, когда вид из списка сопутствующих жертв занимает доминирующее положение в сообществе грызунов.
4. Для решения задач исторической экологии по реконструкции состава фауны и структуры населения мелких млекопитающих по субфоссильным материалам из скоплений погадок филинов необходимо рассматривать отдельно основные и сопутствующие жертвы. Если в состав основных жертв входят виды из зональных комплексов, их характеристика является определяющей. Если в числе основных жертв нет видов из зональных комплексов, реконструкции могут опираться на список сопутствующих жертв с учетом избирательности их добычи.
Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН, а также частично поддержана РФФИ (гранты № 19-04-00507 и 19-04-01008).
Список литературы
Andrews P. Owls, caves and fossils: predation, preservation and accumulation of small mammal bones in caves, with an analysis of the Pleistocene cave faunas from Westbury-sub-Mendip, Somerset, UK. Chicago: University of Chicago Press, 1990. 231 p.
Смирнов Н.Г., Вотяков С.Л., Садыкова Н.О. и др. Физико-химические характеристики ископаемых костных остатков млекопитающих и проблема оценки их относительного возраста. Ч. 1. Термический и масс-спектрометрический элементный анализ. Екатеринбург: “Гощицкий”, 2009. 118 с.
Пукинский Ю.Б. Филин // Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: Наука, 1993. С. 270–289.
Шепель А.И. Хищные птицы и совы Пермского Прикамья. Иркутск: Изд-во Иркут. ун-та, 1992. 296 с.
Mikkola H. Owls of Europe. London: T. & AD Poyser, 1983. 397 p.
Martínez J.A., Zuberogoitia I. The response of the Eagle Owl (Bubo bubo) to an outbreak of the rabbit haemorrhagic disease // J. für Ornithologie. 2001. V. 142. № 2. P. 204–211.
Terry R.C. The dead do not lie: using skeletal remains for rapid assessment of historical small-mammal community baselines // Proceedings of the Royal Society B: Biological Sciences. 2009. V. 277. № 1685. P. 1193–1201.
Смирнов Н.Г., Садыкова Н.О. Источники погрешностей при фаунистической реконструкции в четвертичной палеозоологии // Четвертичная палеозоология на Урале: Сб. науч. трудов. Екатеринбург: Изд-во Урал. ун-та, 2003. С. 98–115.
Смирнов Н.Г. Мелкие млекопитающие Среднего Урала в позднем плейстоцене и голоцене. Екатеринбург: Наука, 1993. 62 с.
Kuzmina E.A., Smirnov N.G., Ulitko A.I. New data on Late Pleistocene–Holocene small mammal communities from the Ural–Sakmara interfluve, Southern Urals // Quaternary international. 2016. V. 420. P. 56–64.
Comay O., Dayan T. Taphonomic signatures of owls: new insights into micromammal assemblages // Palaeogeography, Palaeoclimatology, Palaeoecology. 2018. V. 492. P. 81–91.
Fernández-Jalvo Y., Andrews P., Denys C. et al. Taphonomy for taxonomists: implications of predation in small mammal studies // Quaternary Science Reviews. 2016. V. 139. P. 138–157.
Korpimäki E. Diet composition, prey choice, and breeding success of Long-eared Owls: Effects of multiannual fluctuations in food abundance // Canadian J. of Zoolog. 1992. V. 70. P. 2373–2381.
Korpimäki E., Norrdahl K. Predation of Tengmalm’s owls – numerical responses, functional responses and dampening impact on population fluctuations of microtines // Oikos. 1989. V. 54. P. 154–164.
Громов И.М., Поляков И.Я. Млекопитающие. Л.: Наука, 1977. 502 с. (Фауна СССР; т. 3, вып. 8).
Бобрецов А.В., Лукьянова Л.Е. Динамика населения мелких млекопитающих предгорной тайги Северного Урала // Современное состояние и перспективы развития ООПТ Урала: Мат-лы науч.-практ. конф. Екатеринбург: ООО “УИПЦ”, 2011. С. 65–72.
Модоров М.В., Садыкова Н.О., Собенин С.В. Население мышевидных грызунов природного парка “Оленьи ручьи” по результатам отловов конусами и живоловушками // Биосфера Земли: прошлое, настоящее и будущее. Екатеринбург: “Гощицкий”, 2008. С. 139–142.
Руди В.Н. Фауна млекопитающих Южного Урала. Оренбург: Изд-во ОГПУ, 2000. 207 с.
Большаков В.Н., Васильев А.Г., Васильева И.А., Чибиряк М.В. Популяционная экология мелких млекопитающих пойменных лесов реки Сакмары (Оренбургская область) // Вестник Оренбургского гос. ун-та. 2015. № 10(185). С. 104–107.
Васильев А.Г., Большаков В.Н., Васильева И.А. и др. Фауна насекомоядных млекопитающих и грызунов Губерлинского мелкосопочника (Оренбургская область) // Фауна Урала и Сибири. 2017. № 1. С. 223–244.
Дополнительные материалы отсутствуют.