

Экология, 2019, № 6, стр. 410-418
Влияние температуры и осадков на морфологию, прорастание и стрессоустойчивость семян некоторых представителей флоры Севера
Г. В. Филиппова a, *, Д. Н. Андросова a, Э. В. Филиппов a, И. А. Прокопьев a
a Институт биологических проблем криолитозоны СО РАН
677980 Якутск, просп. Ленина, 41, Россия
* E-mail: nureeva@yandex.ru
Поступила в редакцию 04.12.2018
После доработки 28.02.2019
Принята к публикации 07.03.2019
Аннотация
Исследовано влияние температуры и осадков трех вегетационных сезонов (2014–2016 гг.) на морфологию, массу, прорастание и стрессоустойчивость семян Oxytropis candicans, Melilotus albus, Rumex aquaticus, Allium ramosum, Plantago canescens и Aster alpinus. Вегетационные сезоны оценивали по значению гидротермического коэффициента (ГТК): 2014 г. – засушливый (ГТК = 0.76), 2015 г. – очень засушливый (0.66) и 2016 г. – слабозасушливый (1.05). Показано, что в наиболее благоприятных температурно-влажностных условиях вегетационного сезона 2016 г. были сформированы более развитые семена. Всхожесть семян отличалась по годам наблюдений у пяти видов (M. аlbus, A. alpinus, R. аquaticus, A. ramosum и P. сanescens). Семена 2016 г. имели высокие показатели всхожести и/или темпы прорастания. Выявлены значительные изменения физиологической реакции у семян A. alpinus, R. аquaticus, A. ramosum и O. candicans после действия острого провокационного облучения в разные вегетационные сезоны.
Способность растений адаптироваться к условиям окружающей среды, приобретенная в результате эволюционного развития, обусловила широкое распространение растений на территориях с различными эколого-географическими особенностями. Возможность сохранения гомеостаза и воспроизведения потомства в изменяющихся условиях среды определяется способностью модификации метаболических процессов в пределах, определенных генотипом. Виды растений, имеющие широкий диапазон нормы реакции, в условиях воздействия неблагоприятных факторов обладают высокими адаптационным потенциалом и устойчивостью, что дает возможность произрастать в условиях засухи, переувлажнения, высокой или низкой температуры, засоленности почв и т.д.
Особого внимания требует изучение направленных на воспроизводство потомства приспособительных свойств у растений, произрастающих в районах Севера с суровыми резко континентальными климатическими условиями, высокой вероятностью заморозков в летний период и другими климатогеографическими особенностями. Северные природные экосистемы отличаются значительной потенциальной уязвимостью. Вместе с тем исследований, посвященных адаптации растений к условиям обитания на территории криолитозоны, немного [1, 2].
Цель настоящей работы – анализ изменчивости размеров и массы семян шести видов растений, сформировавшихся в разные вегетационные сезоны, а также оценка физиологических параметров и адаптивного потенциала семенного потомства к воздействию острого γ-облучения.
МАТЕРИАЛ И МЕТОДЫ
Исследовали 6 видов многолетних дикорастущих растений из 5 семейств. Семейство Fabaceae было представлено двумя видами. Остролодочник беловатый (Oxytropis candicans (Pall.) DC.) – бесстебельное растение с каудексом (2n = 16) [3]. Семена собраны в разнотравной степи. Вид хорошо отзывается на увлажнение почвы, увеличивая организменные и популяционные показатели, но не выдерживает конкуренции со стороны других видов. Легко вытесняется на более сухие и неровные местообитания с низким видовым составом и проективным покрытием [4]. Донник белый (Melilotus albus Medik., 2n = 16) распространяется как сорное растение в населенных пунктах, на залежах, по обочинам дорог и на полях [3]. Семена собраны на разнотравном лугу, расположенном у берега озера. Представитель семейства Polygonaceae – щавель водный (Rumex aquaticus L.) – произрастает на болотистых лугах, по болотам, берегам водоемов, на прибрежных песках, галечниках [5]. Семена собраны на заливаемом разнотравно-осоковом лугу прибрежной части озера. Представитель семейства Alliaceae – лук ветвистый (Allium ramosum L.) – встречается на остепненных лугах, в степях, на щебнистых склонах [5]. Семена собраны в разнотравно-типчаковой степи. Вид из семейства Plantaginaceae – подорожник седоватый (Plantago canescens Adams.) – предпочитает довольно богатые почвы, опушки лиственничных лесов, сухие луга, степные склоны, залежи, обочины дорог [6]. Семена собраны на типчаково-разнотравном лугу. Представитель семейства Asteraceae – астра альпийская (Aster alpinus L.) – произрастает на каменисто-щебнистых склонах, в степях, на остепненных лугах, на песчано-галечниковых речных наносах, песчаных обрывах, скалах [7]. Семена собраны на злаково-разнотравном лугу. По отношению к влажности O. candicans, A. ramosum, P. canescens и A. alpinus относятся к экологической группе ксеромезофитов, а M. albus и R. aquaticus – к мезофитам и ортогигрофитам соответственно. По отношению к трофности почвы исследуемые виды являются представителями мезоэутрофов и эутрофов, за исключением O. candicans, который может произрастать в условиях более бедных почв и относится к ортомезотрофам [8].
Сбор семян осуществляли в 2014–2016 гг. на одних и тех же участках, не подвергнутых антропогенным воздействиям и достаточным количеством особей каждого вида, в соответствующие для каждого вида сроки созревания (табл. 1). Для определения показателя “масса 1000 семян” из фракции чистых воздушно-сухих семян отбирали три пробы по 100–250 шт. Морфологическую характеристику семян осуществляли по результатам оптико-визуального обследования с использованием бинокулярного стереоскопического микроскопа МБС-10 (Биолам, Россия) путем измерения линейных размеров длины и ширины не менее чем у 100 семян каждого вида.
Таблица 1.
Сроки прохождения некоторых фаз онтогенеза исследуемых видов растений
| Вид | Отрастание | Цветение | Полное созревание семян |
|---|---|---|---|
| M. albus | Май (II) | Июнь (III) | Август (III) |
| O. candicans | Апрель (III) | Июнь (I) | Август (III) |
| A. alpinus | Апрель (III) | Июнь (III) | Август (I) |
| P. canescens | Май (I) | Июнь (II) | Июль (I–II) |
| R. aquaticus | Май (II) | Июнь (III) | Август (III) |
| A. ramosum | Май (II) | Июнь (II) | Август (III) |
Весной следующего после сбора семян года проводили проращивание в чашках Петри на увлажненных фильтрах, в четырех повторностях по 50–100 шт. в каждой в зависимости от размеров семян при стандартных условиях: t = 23 ± 1°С, фотопериодичность – 16 ч свет/8 ч темнота. Предпосевное хранение осуществляли при 4°С, семена не подвергали дополнительной обработке. Подсчет проросших семян вели ежедневно, энергию прорастания – число проклюнувшихся семян, учитывали на 7-й день от начала прорастания. Всхожесть оценивали по отношению количества проросших семян к количеству заложенных на проращивание, выраженным в %. Критерием оценки выживаемости было отношение количества живых проростков на 30-й день наблюдения к количеству всхожих семян, выраженное в %.
Изучение адаптационных возможностей растений к различным по своей природе стресс-факторам, в том числе радиации, является важным для прогнозирования устойчивости функционирования экосистем в условиях возрастающей антропогенной нагрузки. Оценку адаптивного потенциала семян проводили с использованием провокационного облучения. Для этого семена облучали на γ-установке “ГУР-120” (ВНИИСХРАЭ, г. Обнинск, Россия) в дозах 1, 10, 100, 500 Гр при мощности 60Со-источника 60 Гр/ч. В качестве образцов сравнения (контроль) использовали необлученные семена.
Сравнительный анализ погодных условий вегетационных сезонов проводили с помощью суммы эффективных температур, количества осадков и гидротермического коэффициента (ГТК) по Г.Т. Селянинову [9].
Статистическую обработку данных осуществляли с учетом общепринятых методических указаний по биологической статистике [10]. Для каждого признака определяли среднее арифметическое значение и его ошибку. Сравнение выборок осуществляли методом одно- и двухфакторного дисперсного анализа, значимость отличий определяли с использованием критерия Даннета для множественного сравнения при уровне значимости p ≤ 0.05. Факторный анализ по методу главных компонент (PCA) осуществляли в программной среде Statistica v. 10.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Характеристика погодных условий трех вегетационных сезонов. На основании значений суммы эффективных температур и количества осадков по годам наблюдений были рассчитаны ГТК четырех месяцев, соответствующих периоду вегетации от момента отрастания растений до созревания семян (табл. 2).
Таблица 2.
Погодные условия в период вегетации растений
| Номер декады | Сумма эффективных температур, °С, год | Количество осадков, мм, год | ГТК, год | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2014 | 2015 | 2016 | 2014 | 2015 | 2016 | 2014 | 2015 | 2016 | |
| Май | |||||||||
| I | 49.5 | 10.6 | – | 0.4 | 2.9 | 5.5 | 0.08 | 2.74 | – |
| II | 81.7 | 21.4 | – | – | 6.6 | 2.9 | – | 3.08 | – |
| III | 131.8 | 57.0 | 74.8 | 3.4 | 20.2 | 6.8 | 0.26 | 3.54 | 0.91 |
| Всего | 263.0 | 89.0 | 74.8 | 3.8 (20) | 29.7 (20) | 15.2 (20) | 0.14 | 3.34 | 2.03 |
| Июнь | |||||||||
| I | 115.3 | 136.9 | 168.1 | 14.7 | 8.9 | 6.1 | 1.28 | 0.65 | 0.36 |
| II | 207.5 | 175.0 | 161.5 | 1.2 | 1.2 | 21.2 | 0.06 | 0.07 | 1.31 |
| III | 180.3 | 135.5 | 151.5 | 3.8 | 40.1 | 6.2 | 0.21 | 2.96 | 0.41 |
| Всего | 503.1 | 447.4 | 481.1 | 19.7 (35) | 50.2 (35) | 33.5 (35) | 0.39 | 1.12 | 0.70 |
| Июль | |||||||||
| I | 188.8 | 204.3 | 209.0 | 55.0 | – | 13.3 | 2.91 | – | 0.64 |
| II | 227.5 | 235.5 | 138.2 | 15.8 | 2.2 | 33.4 | 0.70 | 0.09 | 2.42 |
| III | 204.1 | 192.6 | 203.8 | 3.8 | 12.8 | 26.4 | 0.19 | 0.67 | 1.30 |
| Всего | 620.4 | 632.4 | 551.0 | 74.6 (39) | 15.0 (39) | 73.1 (39) | 1.20 | 0.24 | 1.33 |
| Август | |||||||||
| I | 166.4 | 186.4 | 160.2 | 4.2 | 5.6 | 15.5 | 0.25 | 0.30 | 0.97 |
| II | 171.6 | 157.1 | 150.4 | 5.3 | 11.5 | 6.9 | 0.31 | 0.73 | 0.46 |
| III | 156.4 | 185.8 | 83.5 | 34.8 | – | 13.3 | 2.23 | – | 1.59 |
| Всего | 494.4 | 529.3 | 394.1 | 44.3 (37) | 17.1 (37) | 35.7 (37) | 0.90 | 0.32 | 0.91 |
| Итого | 1880.9 | 1698.1 | 1501.0 | 142.4 | 112.0 | 157.5 | 0.76 | 0.66 | 1.05 |
Сравнительный анализ погодных условий вегетационных сезонов показал, что значения ГТК соответствующих месяцев существенно отличались по годам наблюдений. Экстремально низкое количество осадков и, следовательно, недостаточное увлажнение при высоких значениях суммы эффективных температур обусловило очень низкий ГТК в мае (0.14) и июне (0.39) 2014 г., т.е. в месяцы, соответствующие отрастанию и цветению исследуемых видов растений (см. табл. 1 и 2). В 2015 г. подобные условия были зафиксированы в июле (0.24) и августе (0.32), тогда как в мае и июне количество осадков превышало климатическую норму. Вегетационный период 2016 г. характеризовался затяжной весной и низкими значениями суммы эффективных температур (кроме июня) по сравнению с 2014 и 2015 гг. В целом весь вегетационный период 2014 г. оценивали как засушливый (ГТК = 0.76) и наиболее соответствующий среднемноголетним климатическим наблюдениям для Центральной Якутии (ГТКср = 0.72), 2015 г. – очень засушливый (ГТК = 0.66), а 2016 г. – слабозасушливый (ГТК = 1.05).
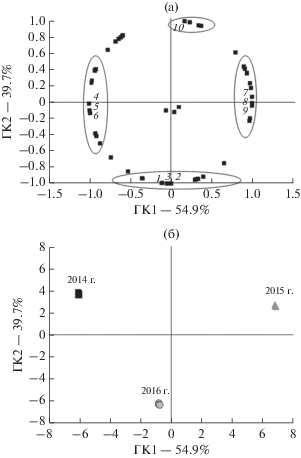
Анализ температурно-влажностных условий вегетации исследуемых видов показал, что отличительные особенности сравниваемых годов в большей степени были обусловлены совокупностью таких показателей, как количество осадков в мае, температура и осадки – в июне, температура середины июля и количество осадков – в августе (рис. 1).
Рис. 1.
Распределение в пространстве первой и второй главных компонент, полученное на основе анализа данных по температурно-влажностным условиям вегетационных сезонов: a – график нагрузок, б – график счетов; 1– осадки в первой декаде августа, 2 – ГТК первой декады августа, 3 – ГТК второй декады июня, 4 – температура июня, 5 – осадки июня, 6 – осадки августа, 7 – ГТК июня, 8 – ГТК второй декады августа, 9 – осадки мая, 10 – температура второй декады июля.
Масса и размер семян по годам наблюдений. Считается, что условия произрастания материнских растений (называемые материнской средой) определяют будущий размер семян, который является проявлением эффекта доступности ресурсов [11, 12]. Согласно исследованиям, проведенным в контролируемых условиях, при доступности воды формируются более развитые семена [13], при низкой температуре увеличивается масса семян [14], а при длительной засухе на стадии формирования семян отмечено значительное снижение их массы [15]. В этой связи в нашем исследовании следовало бы ожидать, что наиболее “крупные” и “тяжелые” семена будут сформированы в 2016 г., наиболее благоприятном с точки зрения температурно-влажностных условий, а более “мелкие” и “легкие” – в 2015 г. Вместе с тем у O. candicans, M. albus, R. aquaticus, A. ramosum и A. alpinus семена с наименьшей массой были сформированы в 2014 г. (табл. 3), занимающем промежуточное положение среди сравниваемых годов по значению ГТК. Низкие значения линейных размеров семян были отмечены у двух видов: R. аquaticus и A. ramosum (табл. 4). В слабозасушливый 2016 г. семена с одновременно большими массой и размерами семян были сформированы у трех видов (M. аlbus, O. сandicans, R. аquaticus; см. табл. 3 и 4). У A. alpinus и A. ramosum наибольшая масса семян была отмечена в очень засушливом 2015 г. Для P. сanescens, имеющего короткие сроки созревания (I–II декады июля), масса семян не отличалась по годам наблюдений, а длина семян была наибольшей в 2016 г. Такого рода стабильность вне зависимости от условий материнской среды была отмечена в работе [16] в отношении семян Sinapis arvensis.
Таблица 3.
Средняя масса 1000 семян по годам наблюдений, г
| Вид | 2014 г. | 2015 г. | 2016 г. |
|---|---|---|---|
| M. albus | 1.75 ± 0.02a | 1.94 ± 0.01b | 1.96 ± 0.01b |
| A. alpinus | 0.38 ± 0.01a | 0.61 ± 0.02b | 0.45 ± 0.01c |
| O. candicans | 1.30 ± 0.01a | 1.33 ± 0.01b | 1.45 ± 0.01c |
| P. canescens | 0.16 ± 0.01a | 0.17 ± 0.001a | 0.17 ± 0.01a |
| R. aquaticus | 1.36 ± 0.01a | 1.53 ± 0.04b | 1.59 ± 0.01c |
| A. ramosum | 2.50 ± 0.05a | 3.28 ± 0.02b | 3.06 ± 0.03c |
Таблица 4.
Морфометрические показатели семян по годам наблюдений, мм
| Вид | 2014 г. | 2015 г. | 2016 г. | |||
|---|---|---|---|---|---|---|
| длина | ширина | длина | ширина | длина | ширина | |
| M. albus | 2.05 ± 0.04a | 1.35 ± 0.02a | 1.88 ± 0.03b | 1.30 ± 0.02b | 2.08 ± 0.03a | 1.30 ± 0.03ab |
| O. candicans | 1.67 ± 0.03a | 1.52 ± 0.02a | 1.63 ± 0.03a | 1.48 ± 0.02a | 1.72 ± 0.02b | 1.60 ± 0.02b |
| A. alpinus | 2.72 ± 0.05a | 1.13 ± 0.03a | 2.12 ± 0.04b | 0.90 ± 0.03a | 2.40 ± 0.05c | 1.10 ± 0.03a |
| P. canescens | 1.35 ± 0.01a | 0.70 ± 0.01a | 1.29 ± 0.05a | 0.71 ± 0.01a | 1.38 ± 0.01b | 0.72 ± 0.01a |
| R. aquaticus | 1.98 ± 0.05a | 0.98 ± 0.02a | 2.90 ± 0.05b | 1.31 ± 0.04b | 3.10 ± 0.04c | 1.52 ± 0.02c |
| A.ramosum | 2.98 ± 0.04a | 1.90 ± 0.04a | 2.98 ± 0.04a | 2.26 ± 0.04b | 3.13 ± 0.05b | 2.15 ± 0.03c |
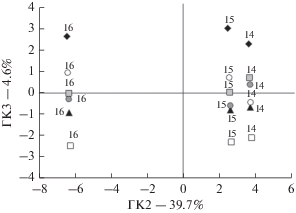
Анализ взаимосвязи температурно-влажностных условий вегетационных сезонов с морфологией и массой семян методом главных компонент позволил выделить две группы растений (рис. 2): первую группу составили семена 2016 г. сбора, вторую – 2014 и 2015 гг.
Рис. 2.
Распределение в пространстве второй и третьей главных компонент, полученное на основе анализа данных по температурно-влажностным условиям вегетационных сезонов и морфологии и массе семян: черные ромбы – A. ramosum, белые круги – R. aquaticus, серые квадраты – M. albus, серые круги – A. alpinus, черные треугольники – O. candicans, белые квадраты – P. canescens; 14 – 2014 г., 15 – 2015 г., 16 – 2016 г.
Таким образом, в наиболее благоприятных температурно-влажностных условиях вегетационного сезона 2016 г. были сформированы более развитые семена всех исследуемых видов растений. Вместе с тем размеры семян по годам наблюдений имели видоспецифический характер, очевидно, обусловленный биологическими особенностями вида. Например, в ряду факторов, оказывающих влияние на качество семенного потомства в условиях ограниченной доступности ресурсов, у O. сandicans могла быть чувствительность к межвидовой конкуренции [4]. Для A. ramosum отмечают разновозрастность цветков в пределах соцветия, которая выражается в неодновременном их распускании, а также влияние облачности на число раскрывающихся цветков. Кроме того, у большинства видов природных фитоценозов в зависимости от внешних условий число завязавшихся семян не всегда соответствует числу семяпочек, что также было отмечено для A. ramosum [1].
В ухудшающихся условиях среды, при нарастании конкурентных отношений между органами, отмечается усиление явления гетерокарпии, свойственного многим семействам, что может быть обусловлено отличиями в размерах проводящих пучков, обеспечивающих физиологически активными и питательными веществами репродуктивные органы в зависимости от пространственного положения [17]. Очевидно, что видовые анатомо-морфологические, физиолого-биохимические и как следствие поведенческие особенности являются определяющими в реализации смешанного типа стратегии выживания, сочетающего черты виолентности, патиентности и эксплерентности, отмеченного для большинства луговых и степных растений [18, 19]. Температурно-влажностные условия, сопутствующие моменту цветения и оплодотворения, по-видимому, критически важны в формировании адаптивной пластичности, поскольку обусловливают в том числе массу будущего семенного потомства.
Характер прорастания и всхожесть семян по годам наблюдений. К важным показателям всхожести дикорастущих растений относят начало, длительность и характер прорастания [20]. Для исследуемых растений семена начали прорастать на 2–4-й день и завершали этот процесс на 12–29-й день (рис. 3).
Рис. 3.
Темпы прорастания и всхожесть семян по годам наблюдений: 1 – 2014 г., 2 – 2015 г., 3 – 2016 г.

Все исследуемые виды характеризовались порционностью прорастания. Семена одного вида, собранные в разные годы, отличались по срокам начала прорастания (M. аlbus, A. alpinus, R. аquaticus, A. ramosum), энергии прорастания, а также длительности прорастания (все виды; см. рис. 3). Для семян, собранных в наиболее благоприятном 2016 г. (ГТК = 1.05), характерны наиболее интенсивные темпы прорастания (за исключением P. сanescens, O. сandicans) в первые 10 дней от момента посева по сравнению с семенами 2014 г. (ГТК = 0.76) и 2015 г. (ГТК = 0.66). Несмотря на задержку начала массового прорастания, снижение энергии прорастания и увеличение сроков прорастания у семян 2014 и/или 2015 гг. сбора, всхожесть в некоторых случаях была сопоставима с семенами 2016 г. Так, статистически достоверно не отличались показатели всхожести семян A. alpinus и P. сanescens 2014 и 2016 гг. сбора, A. ramosum – 2015 и 2016 гг. и O. сandicans – 2014–2016 гг. (см. рис. 3).
Родительские эффекты, которые могут быть результатом взаимодействия генотипа и окружающей среды, включая изменения экспрессии генов, являются определяющими в процессах прорастания семян (вероятности и скорости) и имеют высокие адаптивные последствия для выживания видов [16]. Успешность сохранения и распространения вида во времени и пространстве напрямую связана с качеством семенного потомства, включая покой, обусловливающий формирование почвенного семенного банка дикорастущих растений [21]. Показано, что у одних видов семена разных вегетационных сезонов отличаются по всхожести [22], а у других этот параметр очень стабилен [23]. Анализ влияния окружающих условий на всхожесть семян Arabidopsis thaliana [24] и Plantago lanceolata [25] показал положительное влияние высокой температуры, а для Lactuca sativa, наоборот, низкой [26]. Сухие и теплые условия формирования семян способствовали снижению интенсивности покоя у Ifloga spicata, Rumex pictus и Senecio glaucus по сравнению с влажными [27]. У Cenchrus ciliaris при низкой влажности почвы интенсивность покоя была больше, чем при высокой [28]. Нами обнаружено, что всхожесть семян отличалась по годам наблюдений у пяти из исследованных видов растений (M. аlbus, A. alpinus, R. aquaticus, A. ramosum и P. сanescens). В наиболее благоприятных условиях 2016 г. – низкие температуры и большое количество осадков – были сформированы семена, которые по показателю всхожести и/или темпам прорастания имели наибольшие значения относительно семян 2014 и 2015 гг.
Адаптивный потенциал семян по годам наблюдений. Для анализа реакции семян, собранных в разные годы, на острое провокационное γ-облучение нами были пронормированы показатели выживаемости по отношению к контролю (необлученные семена; рис. 4) соответствующего года. При этом показатели контроля были приняты за “единицу”, что позволило сравнить адаптационный потенциал семян, сформировавшихся в разных погодных условиях вегетационных сезонов 2014–2016 гг. (рис. 5). Поскольку исследуемые виды растений характеризовались порционностью прорастания, растянутой во времени, оценку эффектов проводили на 30-й день – срок, определяющий окончание прорастания и позволяющий оценить начальные эффекты выживаемости проростков. Анализ данных показал, что дозовые зависимости значительно отличались в разные годы.
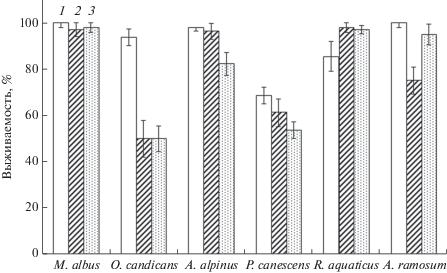
Рис. 5.
Выживаемость проростков в зависимости от дозы провокационного облучения и года созревания семян (2014–2016 гг.): 1 – 2014 г., 2 – 2015 г., 3 – 2016 г.

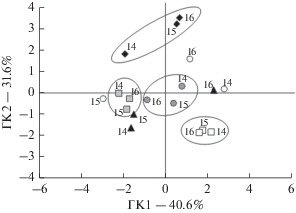
С помощью метода главных компонент были выделены две группы семян, различающиеся по ответам на действие провокационного облучения (рис. 6). Наиболее значимыми из числа используемых оказались дозы 50 и 100 Гр. Радиоустойчивыми по сравнению с другими видами были семена R. аquaticus сбора 2015 г., A. ramosum – 2014 г., A. alpinus – 2016 г., O. сandicans – 2014 и 2015 гг. (слева на рис. 6). Семена M. аlbus вне зависимости от года сбора были радиоустойчивыми, а семена P. сanescens – радиочувствительными (см. рис. 6).
Рис. 6.
Распределение в пространстве первой и второй главных компонент, полученное на основе анализа данных по выживаемости проростков после облучения в дозах 1–500 Гр: черные ромбы – A. ramosum, белые круги – R. aquaticus, серые квадраты – M. albus, серые круги – A. alpinus, черные треугольники – O. candicans, белые квадраты – P. canescens; 14 – 2014 г., 15 – 2015 г., 16 – 2016 г.
Очевидно, что наблюдаемые особенности радиационно-индуцированного адаптивного ответа семян, сформированных в разных температурно-влажностных условиях, стали следствием сочетанного проявления родительского эффекта, который обусловливал результат развертывания генетической информации и воздействия радиации на внутриклеточные мишени семян, сопряженного с изменениями скорости поступления воды в семена, концентрации ингибиторов, регуляторной роли АФК, гормональной сети сигнализации в клетках, наличием повреждений и др. [29, 30]. В целом результаты нашего исследования согласуются с немногочисленными данными о том, что погодные условия среды в период формирования семян могут обусловливать адаптивные эффекты при действии острого γ-облучения [31–33].
Таким образом, межгодовая изменчивость морфологии и массы семян Oxytropis candicans, Melilotus albus, Rumex aquaticus, Allium ramosum, Plantago canescens и Aster alpinus в зависимости от погодных условий 2014–2016 гг. имела видоспецифический характер, очевидно, обусловленный биологическими особенностями каждого из изученных видов. Вместе с тем наиболее развитые семена были сформированы в 2016 г., который характеризовался относительно низкими температурами и большим количеством осадков (ГТК = = 1.05). Всхожесть семян отличалась по годам наблюдений у пяти видов растений (M. аlbus, A. alpinus, R. аquaticus, A. ramosum и P. сanescens). Семена 2016 г. имели наибольшие значения всхожести и/или темпы прорастания относительно семян, сформированных в 2014 г. (ГТК = 0.76) и 2015 г. (ГТК = 0.66). При оценке адаптивного потенциала семян по годам наблюдений на действие провокационного облучения были выявлены значительные изменения физиологической реакции у A. alpinus, R. аquaticus, A. ramosum и O. candicans. Радиоустойчивыми были семена R. аquaticus сбора 2015 г., A. ramosum – 2014 г., A. alpinus – 2016 г., O. сandicans – 2014 и 2015 гг., что подтверждает значимость влияния погодных условий в период вегетации материнских растений на формирование адаптивного ответа у семенного потомства.
Работа выполнена в рамках государственного задания ИБПК СО РАН на 2017–2020 гг. (№ АААА-А17-117020110056-0 и АААА-А17-117020110055-3).
Список литературы
Данилова Н.С. Интродукция многолетних травянистых растений флоры Якутии. Якутск: ЯНЦ СО РАН, 1993. 164 с.
Шеин А.А., Габышев Д.В., Филиппова Г.В., Прокопьев И.А. Морфологические и физиолого-биохимические характеристики Anemone sylvestris (Ranunculaceae) в разных экологических условиях произрастания в Центральной Якутии // Растительный мир Азиатской России. 2013. № 1(11). С. 103–107.
Флора Сибири. Т. 9. Под ред. Положий А.В., Малышева Л.И. Новосибирск: Наука, 1994. 278 с.
Семенова В.В., Данилова Н.С. Онтогенез и структура ценопопуляций Oxytropis candicans (Fabaceae) в Центральной Якутии // Растительные ресурсы. 2017. Т. 53(4). С. 513–526.
Флора Сибири. Т. 4. Под ред. Малышева Л.И., Пешковой Г.А. Новосибирск: Наука, 1987. 246 с.
Флора Сибири. Т. 12. Под ред. Положий А.В., Малышева Л.И. Новосибирск: Наука, 1996. 208 с.
Флора Сибири. Т. 13. Под ред. И.M. Красноборова. Новосибирск: Наука, 1997. 471 с.
Флора Якутии: Географический и экологический аспекты / Л.В. Кузнецова, В.И. Захарова, Н.К. Сосина и др. Новосибирск: Наука, 2010. 192 с.
Лосев А.П. Практикум по агрометеорологическому обеспечению растениеводства. Санкт-Петербург: Гидрометеоиздат, 1994. 246 с.
Зайцев Г.Н. Математический анализ биологических данных. М., 1991. 183 с.
Stanton M.L. Seed variation in wild radish: effect of seed size on components of seedling and adult fitness // Ecology. 1984. V. 65. P. 1105–1112. https://doi.org/10.2307/1938318
Schmid B., Dolt C. Effects of maternal and paternal environment and genotype on offspring phenotype in Solidago altissima L. // Evolution. 1994. V. 48. P. 1525–1549. https://doi.org/10.1111/j.1558-5646.1994.tb02194.x
Wright K.J., Seavers G.P., Peters N.C.B., Marshall M.A. Influence of soil moisture on the competitive ability and seed dormancy of Sinapis arvenis in spring wheat // Weed Research. 1999. V. 39. P. 309–317.https://doi.org/10.1046/j.1365-3180.1999.00147.x
Lacey E.P., Smith S., Case A.L. Parental effects on seed mass: Seed coat but not embryo/endosperm effects // American J. of Botany. 1997. V. 84. № 11. P. 1617–1620. https://doi.org/10.2307/2446624
El Balla M.M.A., Abdelbagi A. Hamid, Abdelmageed A.H.A. Effects of time of water stress on flowering, seed yield and seed quality of common onion (Allium cepa L.) under the arid tropical conditions of Sudan // Agricultural Water Management. 2013. V. 121. P.149–157.https://doi.org/10.1016/j.agwat.2013.02.002
Luzuriaga A.L., Escudero A., PE Rez-garcia F. Environmental maternal effects on seed morphology and germination in Sinapis arvensis (Cruciferae) // Weed Research. 2006. V. 46. P. 163–174.https://doi.org/10.1111/j.1365-3180.2006.00496.x
Левина Р.Е. Способы распространения плодов и семян. М.: Изд-во МГУ, 1957. 361 с.
Grime J.P., Hodson J.G., Hunt R. Comparative Plant Ecology: a functional approach to common british species. London: Unwin Hyman, 1988. 742 p.
Frank D., Klotz S. Biologisch-ökologische daten zur flora in der DDR. Halle-Wittenberg: Martin-Luther-Universität, 1988. 110 p.
Вихирева-Василькова В.В. О прорастании семян некоторых арктических растений // Ботан. журн. 1958. Т. 43. № 7. С. 1024–1029.
Munir J., Dorn L.A., Donohue K., Schmitt J. The effect of maternal photoperiod on seasonal dormancy in Arabidopsis thaliana (Brassicaceae) // American J. of Botany. 2001. V. 88. P. 1240–1249.https://doi.org/10.2307/3558335
Zhang R., Baskin J.M., Baskin C.C. et al. Effect of population, collection year, after-ripening and incubation condition on seed germination of Stipa bungeana // Scientific Reports. 2017. V. 7. P. 1–11.https://doi.org/10.1038/s41598-017-14267-2
Harniss R.O., Mcdonough W.T. Yearly variation in germination in three subspecies of big sagebrush // J. Range Manage. 1976. V. 29. P. 167–168.
Donohue K., Heschel M.S., Chiang G.C. et al. Phytochrome mediates germination responses to multiple seasonal cues // Plant Cell Environ. 2007. V. 30. P. 202–212.https://doi.org/10.1111/j.1365-3040.2006.01619.x
Alexander H.M., Wulff R.D. Experimental ecological genetics in Plantago: X. The effects of maternal temperature on seeds and seedling characters in P. lanceolata // J. of Ecology. 1985. V. 73. P. 271–282. https://doi.org/10.2307/2259783
Koller D. Preconditioning of germination in lettuce at time of fruit ripening // American J. of Botany. 1962. V. 49. P. 841–844. https://doi.org/10.1002/j.1537-2197.1962.tb15017.x
Tielbörger K., Valleriani A. Can seeds predict their future? Germination strategies of density-regulated desert annuals // Oikos. 2005. V. 111. P. 235–244.https://doi.org/10.1111/j.0030-1299.2005.14041.x
Sharifzadeh F., Murdoch A.J. The effects of different maturation conditions on seed dormancy and germination of Cenchrus ciliaris // Seed Science Research. 2000. V. 10. № 4. P. 447–457. https://doi.org/10.1017/S0960258500000490
Bailly C. Active oxygen species and antioxidants in seed biology // Seed Science Research. 2004. V. 14. P. 93–107.
Чурюкин Р.С., Гераськин С.А. Влияние облучения (60Co) семян ячменя на развитие растений на ранних этапах онтогенеза // Радиация и риск. 2013. Т. 22. № 3. С. 80–92.
Позолотина В.Н., Антонова Е.В., Каримуллина Э.М. Оценка радиационного воздействия на ценопопуляции звездчатки злаковой в зоне Восточно-Уральского радиоактивного следа // Экология. 2010. № 6. С. 403–413. [Pozolotina V.N., Antonova E.V., Karimullina E.M. Assessment of radiation impact on Stellaria graminea cenopopulations in the zone of the eastern Ural radioactive // Rus. J. of Ecology. 2010. V. 41(6). P. 459–468.] https://doi.org/10.1134/S1067413610060019
Антонова Е.В., Позолотина В.Н., Каримуллина Э.М. Изменчивость семенного потомства костреца безостого в условиях хронического облучения зоны Восточно-Уральского радиоактивного следа // Экология. 2014. № 6. С. 459–468. [Antonova E.V., Pozolotina V.N., Karimullina E.M. Variation in the seed progeny of Smooth brome Grass, Bromus inermis Leyss., under conditions of chronic irradiation in the zone of the eastern Ural radioactive trace // Rus. J. of Ecology. 2014. V. 45(6). P. 508–516.] https://doi.org/10.1134/S1067413614060034
Прокопьев И.А., Журавская А.Н., Филиппова Г.В. Изменчивость биохимических параметров и радиоустойчивость семенного потомства дейскурании гулявниковой и клоповника безлепестного под действием различных факторов // Экология. 2011. № 4. С. 259–265. [Prokopev I.A., Zhuravskaya A.N., Filippova G.V. Variability of biochemical parameters and radiation resistance of the seed progeny of Descurania sophia and Lepidium apetalum under exposure to various factors // Rus. J. of Ecology. 2011. V. 42. № 4. P. 277–282.] https://doi.org/10.1134/S106741361104014X
Дополнительные материалы отсутствуют.