

Экология, 2019, № 6, стр. 446-455
Цикличность хода численности в разных популяциях обыкновенной бурозубки
Л. Н. Ердаков a, В. В. Панов a, Ю. Н. Литвинов a, *
a Институт систематики и экологии животных СО РАН
630091 Новосибирск, ул. Фрунзе, 11, Россия
* E-mail: litvinov@eco.nsc.ru
Поступила в редакцию 14.02.2018
После доработки 28.11.2018
Принята к публикации 24.01.2019
Аннотация
На примере разных популяций обыкновенной бурозубки (Sorex araneus) проанализированы характерные для вида циклы многолетних колебаний численности. Представлены данные долгосрочных наблюдений в Академгородке (г. Новосибирск) и с. Ровенское (Центральная Бараба). Также использованы опубликованные сведения многолетних наблюдений из европейской и восточносибирской частей ареала этой бурозубки. Целью работы была попытка выработать общее представление о цикличности многолетнего хода численности обыкновенной бурозубки, как инвариантных, так и изменчивых его особенностей в разных географических точках. Ряды данных были проанализированы с помощью спектрального анализа. Основные циклы у этого вида 10–15, 5–7, 3–4 и 2–3-летние, но амплитуда циклов различалась по регионам. Видимо, для поддержания популяционной цикличности важна синхронизация с близкими по параметрам местными ритмами: геомагнитная активность, глубина промерзания почвы, толщина снегового покрова, обводнение территории.
Динамика численности различных популяций млекопитающих и факторы, ее обусловливающие, исследуются давно. Опубликованы примеры многочисленных временных колебаний численных показателей, особенно в популяциях мелких млекопитающих [1–3]. Популяционная динамика обычно рассматривается как результат многофакторного воздействия, а синхронность протекающих процессов – как результат действия мощных синхронизирующих факторов: обилия кормов, климатической ситуации, воздействия хищников и т.п. [4]. Динамика, наблюдаемая у разных видов мелких млекопитающих, является отражением целого комплекса адаптаций к изменяющимся во времени условиям среды. Во многих случаях изменения численности носят циклический характер [5–10].
В последнее время были получены разнообразные оценки механизмов влияния разных факторов среды на популяционную динамику. Например, линейные авторегрессионные модели показали [6, 11], что различные характеристики популяционной динамики, в том числе цикличность, могут быть вызваны множеством конкретных комбинаций прямых и отсроченных зависимостей от плотности популяции. Географические градиенты моделей популяционной динамики также могут быть объяснены сочетанием различных зависимостей от плотности популяции [6, 12–14].
Хотя и существует общее понимание плотностно-зависимой природы популяционной динамики, структура такой зависимости и ее механизмы все еще достаточно сложно интерпретировать. Предложено множество моделей и объяснений динамики (циклики): влияние хищников [15–17], паразитов и инфекций [18–21], плотностно-зависимые механизмы регуляции [6, 22]. Недавние работы по колебаниям численности [23–27] свидетельствуют о том, что интерес к этому направлению не ослабевает.
Обыкновенная бурозубка – вид с обширным ареалом. На большей его части она доминирует по численности среди насекомоядных, а часто и среди всех мелких млекопитающих, что делает ее удобным модельным объектом. Вид хорошо изучен, в литературе много материалов по многолетним наблюдениям за ходом численности этой землеройки [28–32 и др.], однако цикличность хода численности специально не исследуется. Отдельные указания на выявленный тот или иной цикл подаются авторами как интересное открытие [33]. Между тем анализ многолетних материалов о популяционной динамике мелких млекопитающих позволяет получить спектры флуктуаций численных показателей у разных видов животных. Считается, что определенная временная организация биологической системы (например, популяции), характеристикой которой является ее спектр периодов [34], позволяет устанавливать соответствия между биологическими автоколебаниями и периодикой экологических факторов внешней среды. Это происходит с помощью синхронизации циклов близких по значению периодов. Ритмика внешней среды является для биологических осцилляций вынуждающей силой [34]. В.С. Мартынюк [35], исходя из результатов экспериментальных наблюдений биоритмов, делает вывод о том, что в их структуре у живых систем происходит подстройка к периодам во внешней среде.
Видимо, такому направлению исследований есть смысл уделить особое внимание и продолжить изучение биоритмов на популяционном уровне, тем более что динамика численности это традиционная проблема экологии популяций. Поэтому для нас интерес представляли работы, содержавшие достаточно длинные ряды многолетних наблюдений. Именно они дают возможность получать дополнительную информацию о биологических колебаниях разного уровня, которая авторами обычно не анализируется, а часто даже и не отмечается.
Целью работы была попытка выработать общее представление о цикличности многолетнего хода численности обыкновенной бурозубки, как инвариантных, так и изменчивых его особенностей в разных географических точках. Были поставлены следующие задачи:
– оценка статистических характеристик относительной численности в различных популяциях и выявление общих характерных для вида трендов;
– построение спектров динамики численности в различных географических популяциях для выявления гармонических составляющих хода численности;
– уточнение значений периодов в динамике каждой популяции и соотношения их мощностей;
– поиск колебаний природно-климатического фона, с которыми возможна синхронизация различных популяционных циклов.
МАТЕРИАЛ И МЕТОДЫ
При оценках многолетней цикличности численности использованы данные отловов зверьков на стационарах Института систематики и экологии животных СО РАН в Новосибирской области. Материалы сборов частично опубликованы [36, 37]. Кроме того, к анализу привлечены литературные сведения о длительных наблюдениях за ходом численности обыкновенной бурозубки:
Сибирь, стационары института, Новосибирская область (НСО):
– Академгородок (окрестности г. Новосибирска), 1980–2014 гг.;
– с. Ровенское (Каргатский р-н, Барабинская низменность, центральная часть), 1965–1988 гг.;
– с. Усть-Ургулька (Северный р-н, Барабинская низменность, северо-западная часть), 1978–1990 гг.;
– данные по характеристикам климата с метеостанции с. Северное, 40 км от точки исследований.
Литературные данные:
– Польша (Беловежская Пуща), 1986–2007 гг. [33];
– Белоруссия (Брестская область), 1981–2008 гг. [38];
– Карелия (Питкярантский район, стационар Института биологии Карельского научного центра РАН), 1966–1995 гг. [39];
– Стрелецкий лес (Курская область, Центрально-Черноземный заповедник), 1953–1983 гг. [30];
– Воронежская область (Усманский бор, Учебно-научный центр Воронежского университета), 1988–2011 гг. [40];
– Свердловская область, Висимский заповедник, 1987–2012 гг. [41];
– Саяны, Красноярский край, заповедник “Столбы”, 1981–2010 гг. [42].
Обработка данных многолетних наблюдений представляла собой разложение сложной кривой хода численности на составляющую ее простые гармоники, вычисление их величины и мощности. Для выявления скрытых колебаний в численности использовали программу спектрального анализа, находящуюся в собственности ИСиЭЖ СО РАН. Спектральную плотность мощности (СПМ) оценивали методом Уэлча (Welch) [43]. Все расчеты выполнены с привлечением свободной системы для математических вычислений GNU Octave [44, 45], которая использует язык высокого уровня и представляет интерактивный командный интерфейс для решения различных линейных и нелинейных математических задач. Для оценивания СПМ методом Уэлча использовали функцию pwelch из пакета расширений [46]. Каждый анализ многолетнего ряда данных по численности обыкновенной бурозубки генерировал распределение спектра мощности по периодам, которое затем выражали в процентах от максимального значения. Пики на полученном спектре мощности соответствуют главным периодичностям.
Кроме преобразования Фурье, для обработки эмпирических временных рядов в целях выявления в них скрытых периодических составляющих использовали модифицированный метод наименьших квадратов Прони – параметрический метод обработки временного ряда. Основной интерес к методам параметрического спектрального оценивания связан с высокой разрешающей способностью, достигаемой с их помощью при обработке последовательностей данных, содержащих малое число отсчетов. В последнее время метод Прони все чаще используется для таких материалов [47]. Этот математический аппарат опирается, в частности, на линейное прогнозирование вперед и назад. За счет этого становится возможным выделить в исследуемом процессе периодические составляющие с периодом, превышающим длину ряда. Критерием для такого выделения является наилучшая подгонка модели к исходным данным.
Для трендов проверяли гипотезу об отличии коэффициентов от нуля и рассчитывали коэффициент детерминации R2. Полученные значения отражаются в тексте только в том случае, когда нулевая гипотеза о равенстве обоих коэффициентов регрессии нулю отбрасывается. В противном случае линейная регрессия исключается из модели процесса.
Тест на характер распределения используемых выборок показал заметные отклонения от нормального распределения, поэтому был применен ранговый коэффициент корреляции для расчетов связи хода численности с избранными факторами среды.
Для статистической обработки использован пакет программ Past.
РЕЗУЛЬТАТЫ
В сообществах обыкновенная бурозубка, как правило, доминирует среди других насекомоядных. Особенно большая плотность этого вида фиксируется в Сибири. Наибольшая численность наблюдается на Саянах, причем здесь достоверный ее тренд за время наблюдений отсутствовал (табл. 1). В большинстве европейских популяций наблюдался статистически значимый положительный или отрицательный тренд.
Таблица 1.
Статистические характеристики многолетней динамики численности в географических популяциях обыкновенной бурозубки
| Регион | n | М ± m | σ | CV, % | Тренд |
|---|---|---|---|---|---|
| Польша | 22 | 6.66 ± 1.26 | 5.65 | 84.84 | 1.737 + 0.469t; R2 = 0.266; α = 0.05 |
| Белоруссия | 27 | 12.74 ± 1.34 | 6.94 | 54.47 | 20.206 – 0.575t; R2 = 0.432; α = 0.05 |
| Карелия | 34 | 3.11 ± 0.42 | 2.42 | 77.81 | 1.219 + 0.092t; R2 = 0.142; α = 0.05 |
| Стрелецкий лес | 30 | 4.31 ± 1.02 | 5.58 | 129.47 | 2.554 + 0.101t, α = 0.1 |
| Усманский бор | 23 | 3.02 ± 0.530 | 2.54 | 84.11 | 5.445 – 0.220t; R2 = 0.345; α = 0.05 |
| Висимский заповедник | 26 | 4.18 ± 0.844 | 4.30 | 102.87 | 0.899 + 0.262t; R2 = 0.217; α = 0.05 |
| Бараба | 20 | 12.46 ± 2.86 | 12.78 | 102.57 | 17.757 – 0.558t, α = 0.1 |
| Академгородок | 35 | 17.69 ± 2.49 | 14.73 | 83.25 | 18.78 – 0.064t, α = 0.1 |
| Саяны | 30 | 22.81 ± 3.99 | 21.87 | 95.88 | 23.025 – 0.015t, α = 0.1 |
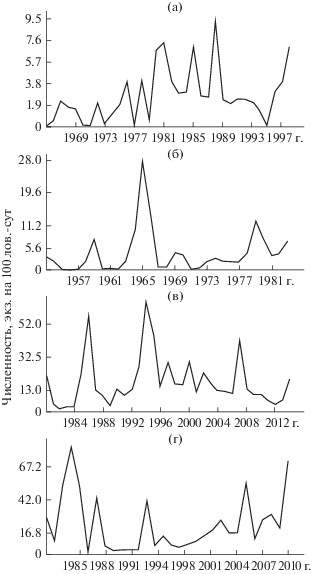
Вариабельность численности у обыкновенной бурозубки может быть высокой как в популяциях, богатых особями (с. Ровенское), так и в относительно малочисленных (Стрелецкий лес). О ходе динамики численности дает представление ее графическое изображение на шкале времени – хронограммы. Их мы приводим для наиболее длительных (30 лет и более) наблюдений (рис. 1). На всех хронограммах ход численности представлен сложной кривой неправильной формы с большим числом пиков и спадов. Расстояние между пиками можно интерпретировать как значение периода какой-либо гармоники, а высота пика дает представление о ее мощности. Полученные таким образом сведения о цикличности, часто применяемые в середине прошлого века, слишком приблизительны. Сейчас можно получить более точные сведения о цикличности, причем обо всем спектре периодических составляющих, содержащихся в данной кривой. Для этого нужно преобразовать хронограмму хода численности в спектр циклов.
Рис. 1.
Хронограммы многолетней динамики численности обыкновенной бурозубки: а – Карелия, б – Стрелецкий лес, в – Академгородок, г – Саяны.
При переносе результатов наблюдений за ходом численности с временной шкалы на частотную мы получили спектры гармонических составляющих (рис. 2). Визуально на большинстве из них доминирует по мощности примерно 7–10-летний цикл. Кроме того, на каждом еще от 2 до 4 гармоник, одна из которых находится в полосе низких частот. Эта цикличность может оказаться очень мощной на спектре (см. рис. 2б, г).
Рис. 2.
Спектры ритмов многолетней динамики численности разных популяций обыкновенной бурозубки: а – Карелия, б – Стрелецкий лес, в – Академгородок, г – Саяны.
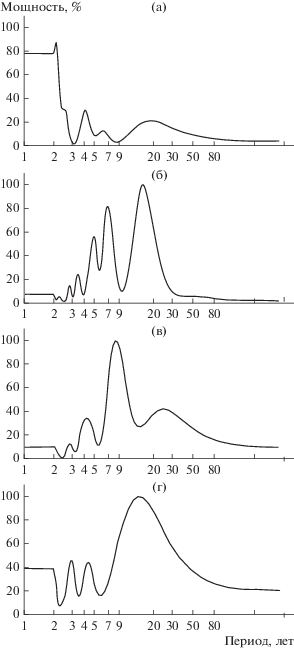
Спектры цикличности популяций бурозубок сходны между собой по наборам гармонических составляющих. Некоторые различия могут быть обусловлены несовпадением по мощности одинаковых гармоник. Так, 4–5-летний ритм есть во всех популяциях, но мощность его наибольшая в популяции из Стрелецкого леса (см. рис. 2б).
При сравнении спектров сибирских популяций обыкновенной бурозубки с европейскими у некоторых из них отмечается большее сходство циклов (см. рис. 2б, в). Более точную информацию можно получить, анализируя табличные данные параметров гармонических составляющих (табл. 2). Видна близость спектров у бурозубок из Карелии и Академгородка – практически все циклы находятся в одних и тех же полосах частот. Стрелецкий лес по набору низкочастотных составляющих отличается от них, но очень близок по соотношению средних и высоких частот со спектрами, полученными из наблюдений за ходом численности в Карелии. На спектре, полученном по данным коротких наблюдений в Новосибирской области (с. Ровенское), проявляются, но в неполном составе циклы, которые имеются и в Академгородке. Длинный ряд наблюдений позволил обнаружить более широкий диапазон цикличностей хода численности обыкновенной бурозубки.
Таблица 2.
Соотношение величины и мощности периодических составляющих многолетней динамики в популяциях обыкновенной бурозубки
| Регион | Период, лет | ||||||
|---|---|---|---|---|---|---|---|
| 11–17 | 8–10 | 5–7 | 3.1–4.9 | 3–3.9 | 2–2.9 | ||
| Польша | – | $\frac{{8.4}}{{1.48}}$ | $\frac{{5.5}}{{1.29}}$ | $\frac{{4.4}}{{1.39}}$ | – | $\frac{{2.9}}{{1.56}}$ | $\frac{{2.3}}{{1.38}}$ |
| Карелия | $\frac{{17.6}}{{0.32}}$ | – | $\frac{{6.1}}{{0.25}}$ | $\frac{{4.1}}{{0.37}}$ | – | $\frac{{2.6}}{{0.39}}$ | $\frac{{2.1}}{{0.66}}$ |
| Беларусь | $\frac{{14.2}}{{1.75}}$ | – | $\frac{{6.4}}{{1.63}}$ | $\frac{{4.3}}{{0.69}}$ | $\frac{{3.4}}{{0.79}}$ | – | $\frac{{2.3}}{{1.59}}$ |
| Стрелецкий лес | $\frac{{15.3}}{{2.24}}$ | – | $\frac{{6.9}}{{2.02}}$ | $\frac{{4.9}}{{1.68}}$ | $\frac{{3.5}}{{1.09}}$ | – | $\frac{{2.3}}{{0.44}}$ |
| Усманский бор | – | $\frac{{8.6}}{{0.83}}$ | – | – | $\frac{{3.5}}{{0.43}}$ | $\frac{{2.4}}{{0.43}}$ | $\frac{{2.1}}{{0.49}}$ |
| Висимский заповедник | $\frac{{10.0}}{{4.31}}$ | – | $\frac{{6.2}}{{0.62}}$ | $\frac{{4.6}}{{0.97}}$ | $\frac{{3.5}}{{1.17}}$ | $\frac{{2.8}}{{1.09}}$ | $\frac{{2.1}}{{1.29}}$ |
| Ровенское | $\frac{{14.2}}{{6.08}}$ | – | – | – | $\frac{{3.5}}{{1.18}}$ | $\frac{{2.8}}{{1.76}}$ | – |
| Академгородок | $\frac{{10.0}}{{4.68}}$ | $\frac{{7.2}}{{5.68}}$ | $\frac{{5.3}}{{1.34}}$ | $\frac{{4.2}}{{3.82}}$ | $\frac{{3.5}}{{2.57}}$ | $\frac{{2.7}}{{2.63}}$ | $\frac{{2.1}}{{1.89}}$ |
| Саяны | $\frac{{13.8}}{{7.66}}$ | – | – | $\frac{{4.4}}{{5.09}}$ | $\frac{{3.0}}{{5.23}}$ | – | – |
Специальное исследование воздействия погодных условий на динамику многолетней численности обыкновенной бурозубки мы провели на стационаре института в Северной Барабе (с. Усть-Ургулька). Для изучения возможностей синхронизации циклов зверьков с погодными условиями привлекли данные по характеристикам климата с ближней метеостанции. Сравнение динамики изменения численности с ходом метеохарактеристик проведено в те же сроки.
Прежде всего мы рассчитали коэффициенты корреляции с избранными факторами. Оказалось, что они у всех факторов, кроме обводнения11 территории, ниже 5% уровня значимости, т.е. недостоверны у наших выборок. Далее, исходя из значений циклов, оценили возможности популяции обыкновенных бурозубок подстраивать свою цикличность к близким колебаниям этих же факторов. Наличие близких по периоду циклов численности и параметров климата указывает на возможность их синхронизации. Для этого мы прибегли к параметрическому методу анализа временных рядов, использовав преобразование Прони [47].
При сравнении циклов численности землеройки с климатическими видно, что погодные факторы имеют в своей многолетней динамике гармонические составляющие, близкие по периоду к популяционным циклам, т.е. имеются возможности для синхронизации циклов (табл. 3). Самая мощная периодическая составляющая (это низкочастотный ритм) численности бурозубки может синхронизироваться с температурными факторами. Легче всего подстройка будет происходить к динамике промерзания и продолжительности температурных минимумов, но легко подстроится и к колебаниям как глубины промерзания почвы, так и к динамике толщины снега.
Таблица 3.
Соотношение периодических составляющих хода численности обыкновенной бурозубки с цикличностью местных погодных факторов (Западная Сибирь, Северная Бараба, с. Усть-Ургулька)
| Характеристики | Период, лет | |||||
|---|---|---|---|---|---|---|
| 11–15 | 8–10 | 5–7 | 2.9–4 | 2–2.6 | ||
| Численность | $\frac{{11.9}}{{10.55}}$ | – | $\frac{{5.02}}{{6.20}}$ | $\frac{{3.2}}{{3.95}}$ | $\frac{{2.6}}{{5.30}}$ | |
| Температура | Глубина промерзания | $\frac{{13.3}}{{12.33}}$ | – | $\frac{{4.2}}{{15.93}}$ | $\frac{{3.1}}{{12.45}}$ | $\frac{{2.3}}{{14.75}}$ |
| Минимальная температура | $\frac{{12.7}}{{8.17}}$ | – | $\frac{{6.1}}{{1.68}}$ | $\frac{{3.5}}{{8.83}}$ | $\frac{{2.2}}{{4.22}}$ | |
| Толщина снега | $\frac{{13.8}}{{1.09}}$ | – | $\frac{{4.4}}{{4.28}}$ | $\frac{{2.9}}{{1.78}}$ | $\frac{{2.3}}{{8.34}}$ | |
| Влажность | Обводнение | – | – | $\frac{{6.9}}{{29.31}}$ | $\frac{{4.1}}{{19.83}}$ | $\frac{{2.4}}{{11.31}}$ |
| Осадки, июнь | – | – | $\frac{{6.7}}{{26.62}}$ | $\frac{{4.2}}{{27.1}}$ | $\frac{{2.4}}{{15.81}}$ | |
| Осадки, среднегодовые | – | $\frac{{9.9}}{{71.65}}$ | $\frac{{6.0}}{{75.10}}$ | $\frac{{3.0}}{{16.75}}$ | – | |
В области средних частот бурозубка имеет мощный 5-летний ритм, и все колебания погодных факторов в данной местности также имеют очень близкую к нему гармоническую составляющую (см. табл. 3). У мощного цикла в высоких частотах прослеживаются сходные по периоду гармонические составляющие местных климатических факторов. Особенно важны для подстройки, видимо, колебания июньских осадков и общего обводнения территории (см. табл. 3).
В многолетних колебаниях численности бурозубки наблюдается и 3-летний цикл – он невелик по мощности, но с его помощью также можно подстроить изменения многолетней численности практически ко всем отслеженным факторам климата. Наиболее легко бурозубка с помощью этого колебания численности может подстроиться к динамике глубины промерзания почвы, среднегодовых осадков и многолетнему изменению толщины снегового покрова (см. табл. 3).
ОБСУЖДЕНИЕ
В литературе отмечено [48, 49], что вариабельность численности несколько снижается в центральных частях ареала в подзоне южной тайги, смешанных и широколиственных лесов. По нашим данным, могут быть и другие варианты ее распределения (см. табл. 1): практически в центре ареала (Стрелецкий лес, Висимский заповедник) вариабельность изменений численности оказывается наибольшей, или, например, в центре ареала (Стрелеций лес, Березинский заповедник) она значительна, а на этой же широте к западу (Брестская область) численность вдвое стабильнее. Во всех рассмотренных географических популяциях присутствуют колебания численности, их диапазон иллюстрируют спектры периодических составляющих.
В европейских популяциях обыкновенной бурозубки спектры цикличности различаются по числу периодических составляющих. Северная популяция (Карелия) имеет три такие гармоники, тогда как на спектре южной (Стрелецкий лес) пять значительных пиков (см. рис. 2а, б). Различия есть и в соотношении мощностей циклов. Кроме того, доминирующий цикл северной популяции заметно больше по периоду – он более чем 17-летний, тогда как в южных районах примерно 10–15-летний. Можно предположить, что при продвижении на север период увеличивается.
Спектры популяционных колебаний бурозубок из Сибири сходны между собой по наборам гармонических составляющих. Некоторые различия здесь обусловлены несовпадением одинаковых гармоник по мощности: саянская популяция имеет более мощные ритмы в низких частотах, а западносибирская – в средних (см. рис. 2в, г).
Расчеты спектров многолетней цикличности обыкновенной бурозубки в разных районах ареала дали возможность описать некий обобщенный спектр ее популяционных циклов, т.е. выделить в нем инвариантные периодические составляющие. Видимо, для этой бурозубки всегда характерны 10–15, 5–7, 3–4 и 2–3-летние колебания численности. Тем не менее наблюдается и большая изменчивость как периодов популяционных колебаний, так и их мощностей (см. табл. 2). Ее могут вызвать различия в условиях местообитаний, которые определяются как биотопическими особенностями, так и неоднородностью природно-климатического фона. При расчетах спектров бурозубок из Беловежской Пущи по данным [33] нами обнаружено, что условия местообитания оказали существенное влияние на цикличность их численности. Хотя большее число гармонических составляющих у них близко, но в зоне средних частот на лугу для землеройки характерна доминирующая 6-летняя цикличность, а на берегу реки этой периодической составляющей в спектре цикличности нет. Вместо нее имеются две равные по мощности доминирующие гармоники – 8- и 4-летняя.
Еще более заметные коррективы могут внести в картину спектра колебаний метеорологические факторы. Сравнение синхронизации нескольких наиболее важных с нашей точки зрения для обыкновенной бурозубки факторов климата проведено в Усть-Ургульке (Северная Бараба). У бурозубки, круглогодично активного зверька, мелкого, с высоким уровнем обмена, одним из важнейших должен быть температурный фактор. И это не только динамика атмосферной температуры, а также толщина снегового покрова, глубина промерзания почвы и продолжительность периода с отрицательными температурами. Бурозубка также зависима и от влажности климата. Из характеристик увлажнения мы взяли среднегодовые осадки, осадки в июне и обводнение территории. Последнее является комплексным воздействием, влияющим на динамику элементов ландшафта (остепнение, заболачивание). У популяции обыкновенной бурозубки в Северной Барабе имеется много возможностей синхронизации к важным для нее колебаниям климатических факторов (см. табл. 3).
Популяция, как автоколебательная система, имеет собственные колебания многих характеристик. Из них мы рассматриваем только численность. Это интегральная характеристика, изменяя которую можно сохранять соответствие с колебаниями условий среды. Важные для популяции колебания могут “подстраиваться” к ритмам среды и тем самым приобретать устойчивость. Такая подстройка может осуществляться по типу синхронизации к близким по периоду колебаниям, например климатических или других факторов.
Цикличность осадков рассчитана давно [53]. Обычные ритмы для них 10–12, 8–10, 3–4, а также 2–3-летние. Практически в каждой географической популяции обыкновенной бурозубки имеются колебания такой периодичности (см. табл. 2). Наиболее распространенными для температурных циклов являются 2–4, 5–6, 7–9, 10–12-летние [54]. Широкий диапазон автоколебаний численности у обыкновенной бурозубки дает возможность подстроиться в любом местообитании к соответствующему температурному циклу.
Обычно изменения температуры и осадков происходят совместно. Для периодичности этих климатических особенностей также известна регулярная изменчивость в 3–4 и 7–11 лет [55, 56]. Это очень важные для бурозубок колебания, ведь именно в этих частотных полосах находятся наибольшие по мощности многолетние гармонические составляющие, характерные для этого вида (см. табл. 2).
Отмечены изменения геомагнитной активности с 15–17-летней периодичностью [50]. К такому ритму могут подстраиваться популяции из Стрелецкого леса и Белоруссии, которые имеют сходные колебания численности. В Карелии тесная связь показателей численности обыкновенной бурозубки выявлена с геомагнитной активностью и суммой осадков в мае [39]. На положительное влияние суммы осадков прошедшего года и температур прошедшего июня на популяцию землероек указывают многие авторы [51, 52].
Несмотря на значительную экологическую изученность обыкновенной бурозубки, цикличность хода ее численности специально не исследовалась, и об этом имеются лишь отрывочные сведения. Поэтому новизна нашей работы определяется самой ее целью – изучением цикличности многолетнего хода численности в популяциях обыкновенной бурозубки.
Если фиксируется наличие какого-либо цикла в популяционной динамике, то авторы могут специально отметить это в тексте как “интересное новое открытие”. Так поданы сведения о наличии у обыкновенной бурозубки 3-летнего ритма численности на лугах между лесными массивами [33] или о 3- и 5-летней цикличности у красной полевки [57]. По-прежнему оценка циклов может производиться на глаз по расстоянию между пиками на хронограмме [40, 52, 58]. Чаще всего наличие циклов просто регистрируется и не является главным в исследовании [29]. Даже при отсутствии специального анализа популяционной цикличности могут быть сделаны крупные обобщения об уменьшении цикличности микромаммалий с широтой [14].
Между тем цикличность очень информативна и достойна специального изучения. Именно она лежит в основе хроноэкологического анализа. В настоящее время происходит первичное накопление информации о количестве гармонических составляющих в популяционных спектрах цикличности у различных видов, об экологических и таксономических особенностях таких спектров, чему и посвящена наша работа.
ВЫВОДЫ
1. Для популяций обыкновенной бурозубки характерны циклы многолетних колебаний численности, которые прослеживаются в близких полосах частот в разных участках ареала. Основные циклы у этого вида 10–15, 5–7, 3–4 и 2–3-летние. По мощности всегда преобладают низкочастотные ритмы численности.
2. В различных регионах популяции синхронизируют свои циклы численности к местным природным ритмам, поэтому из всего диапазона автоколебаний в локальном местообитании становятся устойчивыми только сходные по частоте с колебаниями внешних условий. Это создает различия в картинах спектров цикличностей у разных географических популяций.
3. Для поддержания популяционной цикличности важна ее подстройка к близким по периодам воздействия природно-климатическим колебаниям. Такие ритмы бурозубок могут синхронизироваться с геомагнитной активностью, глубиной промерзания почвы, толщиной снегового покрова, обводнением территории и пр. Показано, что именно к колебаниям показателей этих внешних факторов синхронизированы многие циклы динамики численности обыкновенной бурозубки.
4. Статистические характеристики относительной численности в различных популяциях имеют значительную вариабельность, обычно не зависимую от ее средней многолетней величины. Для численности всех популяций обыкновенной бурозубки характерен очень небольшой тренд. Причем если в европейских районах ареала он положительный, то в большинстве сибирских – отрицательный.
Исследование поддержано Программой фундаментальных научных исследований государственных академий наук на 2013–2020 гг., проект № VI.51.1.4 (АААА-А16-116121410119-4) и РФФИ (грант 17-04-00269). Авторы подтверждают отсутствие конфликта интересов.
Список литературы
Садыков О.Ф., Бененсон И.Е. Динамика численности мелких млекопитающих: Концепции, гипотезы, модели. М.: Наука, 1992. 191 с.
Жигальский О.А. Анализ популяционной динамики мелких млекопитающих // Зоол. журн. 2002. Т. 81. № 9. С. 1078–1106.
Cornulie Th., Yoccoz N.G., Bretagnolle V. et al. Europe-wide dampening of population cycles in keystone herbivores // Science. 2013. V. 340. № 6128, P. 63–66. https://doi.org/10.1126/science.1228992
Щипанов Н.А. Популяция как единица существования вида. Мелкие млекопитающие // Зоол. журн. 2003. Т. 82. № 4. С. 450–469.
Korpimäki E., Oksanen L., Oksanen T. et al. Vole cycles and predation in temperate and boreal zones of Europe // J. Anim. Ecol. 2005. V. 74. P. 1150–1159.
Stenseth N.C. Population cycles in voles and lemmings: density dependence and phase dependence in a stochastic world // Oikos. 1999. V. 87. № 3. P. 427–461. https://doi.org/10.2307/3546809
Krebs C.J., Reid D., Kenney A.J., Gilbert S. Fluctuations in lemming populations in north Yukon, Canada, 2007–2010 // Can. J. Zool. 2011. V. 89. № 4. P. 297–306. https://doi.org/10.1139/Z11-004
Литвинов Ю.Н., Ковалева В.Ю., Ефимов В.М., Галактионов Ю.К. Цикличность популяции водяной полевки как фактор биоразнообразия в экосистемах Западной Сибири // Экология. 2013. № 5. С. 383–388. [Litvinov Y.N., Kovaleva V.Y., Efimov V.M., Galaktionov Y.K. Cyclicity of the European water vole population as a factor of biodiversity in ecosystems of Western Siberia. Russ. J. Ecol. 2013. V. 44. № 5. P. 422–427. doi 10.1134/S1067413613050081]https://doi.org/10.7868/S0367059713050089
Brommer J.E., Pietiainen H., Ahola K. et al. The return of the vole cycle in southern Finland refutes the generality of the loss of cycles through ‘climatic forcing’ // Glob. Chang. Biol. 2010. V. 16. № 2. P. 577–586.
Ердаков Л. Биологические ритмы: особь, популяция, сообщество. Цикличность в живых системах. LAP Lambert, 2011. 152 c.
Royama T. Analytical Population Dynamics. London: Chapman & Hall, 1992. 380 p.
Bjørnstad O.N., Falck W., Stenseth N.C. A geographic gradient in small rodent density fluctuations: a statistical modelling approach // Proc. R. Soc. Biol. Sci. 1995. V. 262. № 1364. P. 127–133. https://doi.org/10.1098/rspb.1995.0186
Saitoh T., Stenseth N., Bjornstad O. Density dependence in fluctuating grey-sided vole populations // J. Anim. Ecol. 1997. V. 66. № 1. P. 14–24. https://doi.org/10.2307/5960
Tkadlec E., Stenseth N.C. A new geographical gradient in vole population dynamics // Proc. R. Soc. Biol. Sci. 2001. V. 268. № 1476. P. 1547–1552. https://doi.org/10.1098/rspb.2001.1694
Hanski I., Henttonen H. Predation on competing rodent species: a simple explanation of complex patterns // J. Anim. Ecol. 1996. V. 65. № 2. P. 220–232. https://doi.org/10.2307/5725
Lambin X., Petty S.J., Mackinnon J.L. Cyclic dynamics in field vole populations and generalist predation // J. Anim. Ecol. 2000. V. 69. № 1. P. 106–119. https://doi.org/10.1046/j.1365-2656.2000.00380.x
Sundell J., Huitu O., Henttonen H. et al. Large-scale spatial dynamics of vole populations in Finland revealed by the breeding success of vole-eating avian predators // J. Anim. Ecol. 2004. V. 73. № 1. P. 167–178. https://doi.org/10.1111/j.1365-2656.2004.00795.x
Dobson A.P., Hudson P.J. Microparasites: observed patterns in wild animal populations // Ecology of infectious diseases in natural populations / Grenfell B.T., Dobson A.P. Eds. Cambridge: Cambridge University Press, 1995. P. 52–89.
Gulland F.M.D. Impact of infectious diseases on wild animal populations: a review // Ecology of Infectious Diseases in Natural Populations / Grenfell B.T., Dobson A.P., Eds. Cambridge: Cambridge University Press, 1995. P. 20–51. https://doi.org/10.1017/CBO9780511629396.002
Hudson P.J., Dobson A.P., Newborn D. Prevention of population cycles by parasite removal // Science. 1998. V. 282. № 5397. P. 2256–2258. https://doi.org/10.1126/science.282.5397.2256
Tompkins D.M., Dobson A.P., Arneberg P. et al. Parasites and host population dynamics // The ecology of wildlife diseases / Hudson P.J., Rizzoli A., Heesterbeek H., Dobson A. Eds. Oxford: Oxford University Press, 2002. P. 45–62.
Agrell J., Erlinge S., Nelson J. et al. Delayed density-dependence in a small-rodent population // Proc. R. Soc. Biol. Sci. 1995. V. 262. № 1363. P. 65–70. https://doi.org/10.1098/rspb.1995.0177
Claussen J.C., Traulsen A. Cyclic dominance and biodiversity in well-mixed populations // Phys. Rev. Lett. 2008. V. 100. 058104. https://doi.org/10.1103/PhysRevLett.100.058104
Korpela K., Delgado M., Henttonen H. et al. Nonlinear effects of climate on boreal rodent dynamics: mild winters do not negate high-amplitude cycles // Glob. Chang. Biol. 2013. V. 19. P. 697–710. https://doi.org/10.1111/gcb.12099
Barraquand F., Pinot A., Yoccoz N., Bretagnolle V. Overcompensation and phase effects in a cyclic common vole population: between first and second-order cycles // J. Anim. Ecol. 2014. V. 83. № 6. P. 1367–1378. https://doi.org/10.1111/1365-2656.12257
Fauteux D., Cheveau M., Imbeau L., Drapeau P. Cyclic dynamics of a boreal southern red-backed vole population in northwestern Quebec // J. Mammal. 2015. V. 96. № 3. P. 573–578. https://doi.org/10.1093/jmammal/gyv062
Фрисман Е.Я., Неверова Г.П., Кулаков М.П., Жигальский О.А. Явление мультирежимности в популяционной динамике животных с коротким жизненным циклом // Докл. РАН. 2015. № 4. С. 488–493. [Frisman E.Y., Neverova G.P., Kulakov M.P., Zhigalskii O.A. Multimode phenomenon in the population dynamics of animals with short live cycles // Dokl. Biol. Sci. 2015. Т. 460. № 1. С. 42–47. doi 10.1134/S0012496615010111]https://doi.org/10.7868/S0869565215040258
Ивантер Э.В., Якимова А.Е. Численность и экологическая структура населения мелких млекопитающих // Мониторинг и сохранение биоразнообразия таежных экосистем Европейского Севера России. Петрозаводск: КарНЦ РАН, 2010. С. 170–191.
Sheftel B.I. Long-term and seasonal dynamics of shrews in Central Siberia // Ann. Zool. Fennici. 1989. V. 9. P. 357–369.
Ходашова К.С., Елисеева В.И. Землеройки в экосистемах центральной лесостепи Русской равнины. М.: Наука, 1992. 112 с.
Kaikusalo A., Hanski I. Population dynamics of the common and masked shrews (Sorex araneus and Sorex caecutuens) in Finnish Lapland // Acta Zool. Fennica. 1985. V. 173. P. 283–285.
Henttonen H., Haikusalmi V., Kaikusalo A. et al. Long- term population dynamics of the common shrew Sorex araneus in Finland // Ann. Zool. Fennici. 1989. V. 26. P. 349–355.
Zub K., Jędrzejewska B., Jędrzejewski W., Bartoń K.A. Cyclic voles and shrews and non-cyclic mice in a marginal grassland within European temperate forest // Acta Theriologica. 2012. V. 57. № 3. P. 205–216. https://doi.org/10.1007/s13364-012-0072-2
Мартынюк В.С., Владимирский Б.М., Темурьянц Н.А. Биологические ритмы и электромагнитные поля среды обитания // Геофизические процессы и биосфера. 2006. Т. 5. № 1 С. 5–23.
Мартынюк В.С. Космические и геофизические процессы и биологические ритмы // IX Международная крымская конференция “КОСМОС И БИОСФЕРА”. Алушта, 2011. www.biophys.ru/archive/ crimea2011/absr-p41.pdf
Глотов И.Н., Ердаков Л.Н., Кузякин В.А. и др. Сообщества мелких млекопитающих Барабы. Новосибирск: Наука, 1978. 231 с.
Максимов А.А., Ердаков Л.Н. Циклические процессы в сообществах животных. Новосибирск: Наука, 1985. 234 с.
Демянчик В.Т. Многолетняя динамика лесных видов микромаммалий Micromammalia на Выгонощанском лесо-болотном массиве // Науковий вicнuк Волинського нацiоналъного yniвеpcumemy imeni Лесi Украiнки. Роздiл II. Тваринний cвiт. 2009. Т. 2. С. 234–238.
Балакирев А.Е., Окулова Н.М., Ивантер Э.В. К анализу факторных воздействий на многолетнюю динамику численности обыкновенной бурозубки на севере и юге ареала // Поволжский экологич. журн. 2004. № 2. С. 111–122.
Климов А.С. Многолетняя динамика численности и современное состояние видов мелких млекопитающих низинных болот Усманского бора (Воронежская область) // Поволжский экологич. журн. 2013. № 1. С. 47–51.
Лукьянова Л.Е. Мелкие млекопитающие в экологически дестабилизированной среде (последствия локальных природных катастроф): Автореф. дис. … докт. биол. наук. Екатеринбург: Ин-т экологии растений и животных УРО РАН, 2013. 42 с.
Виноградов В.В. Пространственно-временная организация сообществ мелких млекопитающих приенисейской части Алтае-Саянской горной страны: Автореф. дис. … докт. биол. наук. Новосибирск, 2011. 43 с.
Марпл-мл. С.Л. Цифровой спектральный анализ и его приложения. М.: Мир, 1990. 584 с.
GNU Octave. http://www.gnu.org/software/octave/
GNU Octave. http://ru.wikipedia.org/wiki/GNU_Octave
Octave-Forge [http://octave.sourceforge.net/signal/ function/pwelch.html]
Кухаренко Б.Г. Исследование по методу Прони динамики систем на основе временных рядов // Труды МФТИ. 2009. Т. 1. № 2. С. 176–192.
Викторов Л.В. Результаты количественного учета землероек в Калининской области // Учен. зап. Калинин. пед. ин-та. 1964. Т. 31. С. 74–99.
Попов И.Ю. Многолетняя динамика численности и распределение землероек на постоянных площадках // Териофауна России и сопредельных территорий (VII съезд Териол. о-ва): Мат-лы междунар. совещ. М., 2003. С. 274–275.
Рубашев Б.М. Проблемы солнечной активности. М., Л.: Наука, 1964. 362 с.
Окулова Н.М., Куприянова И.Ф., Сивков А.В. Динамика численности мелких млекопитающих Пинежского заповедника. Сообщ. 1. Обыкновенная бурозубка Sorex araneus L. // Териологические исследования. СПб.: ЗИН РАН, 2003. Вып. 4. С. 38–46.
Каштальян А.П. О влиянии погодно-климатических факторов на осеннюю динамику численности обыкновенной бурозубки Sorex araneus в северной Беларуси // Териофауна России и сопредельных территорий (VII съезд Териол. о-ва): Мат-лы междунар. совещ. М., 2003. С. 155.
Дроздов О.В., Григорьева А.С. Многолетние циклические колебания атмосферных осадков на территории СССР. Л.: Гидрометеоиздат, 1971. 160 с.
Коротина Е.Ф. Многолетние колебания температурного режима Южного Урала: Автореф. дис. … канд. географ. наук. Пермь: Перм. гос. ун-т, 2002. 23 с.
Дружинин И.П. Долгосрочный прогноз и информация. Новосибирск: Наука, 1987. 251 с.
Кривенко В.Г. Водоплавающие птицы и их охрана. М.: Агропромиздат, 1991. 271 с.
Boonstra R., Krebs C.J. Population dynamics of red-backed voles (Myodes) in North America // Oecologia. 2012. V. 168. P. 601–620.
Ивантер Э.В., Коросов А.В., Якимова А.Е. Статистический анализ многолетних изменений численности мелких млекопитающих северо-восточного Приладожья // Вопросы популяционной биологии / Тр. ПетрГУ. Сер. биол. Вып. 2. Петрозаводск: Изд-во ПетрГУ, 2008. С. 5–24.
Дополнительные материалы отсутствуют.