

Экология, 2019, № 6, стр. 471-474
Формирование популяционной специфики озерной лягушки (Pelophylax ridibundus) в водоемах Среднего Урала
Н. Л. Иванова a, *, Д. Л. Берзин a, **
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: ivanova@ipae.uran.ru
** E-mail: smithbdl@rambler.ru
Поступила в редакцию 26.10.2018
После доработки 24.12.2018
Принята к публикации 06.02.2019
Известно, что инвазии чужеродных видов могут быть как причиной вымирания аборигенных видов, так и потери биоразнообразия [1]. Есть свидетельства, что биоразнообразие регионов растет вследствие расселения новых видов животных и растений [2]. Поэтому актуально изучение чужеродных видов, тем или иным способом появившихся в фауне региона: их влияние на аборигенную фауну, способы адаптаций чужеродных видов к новым условиям существования, так как от этого зависит, войдет ли вид в список, увеличивающий биоразнообразие региона, или исчезнет. Возникающие в новых регионах географически изолированные популяции могут отличаться в первичном ареале как по своим экологическим, морфофизиологическим, так и генетическим характеристикам [3].
Один из чужеродных видов амфибий на Среднем Урале – озерная лягушка (Pelophylax ridibundus Pallas, 1771). Этот вид далеко распространился за пределы своего естественного ареала и проник во многие регионы. Описаны популяции в термальных водоемах в окрестностях г. Якутска [4], на Алтае [5], в Красноярском крае [6], на Камчатке [7]. Озерная лягушка – высокотолерантный и экологически пластичный вид, способный существовать там, где другие виды земноводных не могут нормально расти, развиваться и воспроизводиться [8–10].
Озерная лягушка появилась в герпетофауне Среднего Урала в 70-е годы XX в. при зарыблении водохранилищ-охладителей Рефтинской и Верхнетагильской ГРЭС. Вместе с мальками рыб в водоемы попали головастики, источником которых были лягушки из водоемов Краснодарского края и Украины [11, 12].
Распространение P. ridibundus на Среднем Урале продолжается; колонизированы водоемы на значительном расстоянии от центров интродукции. Из Верхнетагильского водохранилища P. ridibundus по р. Тагил, достигнув питьевых и отстойных прудов в черте г. Нижний Тагил, проникли в верховья р. Нейвы [9], их наблюдали в водоеме-охладителе Серовской ГРЭС, р. Исеть, Белоярском водохранилище. Расселение происходит благодаря сезонным изменениям гидрологического режима водоемов и при активных миграциях в наземной среде. При этом P. ridibundus не заселяет прибрежные мелководные участки водоемов, где в период нереста скапливаются лягушки рода Rana, а предпочитает относительно крупные водоемы с крутыми берегами и обильной водной растительностью. В спектре питания P. ridibundus неизменно присутствуют корма, которых, как правило, избегают аборигенные виды земноводных, поэтому P. ridibundus не представляют угрозы для местных амфибий [13].
Цель настоящей работы – оценить важнейшие экологические характеристики инвазивного вида озерной лягушки (P. ridibundus Pallas, 1771): размерно-возрастной состав популяций, сроки размножения, плодовитость, а также продолжительность жизни в естественных водоемах на территории Среднего Урала – р. Пышма и термальных – Рефтинское водохранилище.
Рефтинское водохранилище создано на р. Рефт в 1968 г. – левобережном притоке р. Пышмы Обь-Иртышского бассейна. Площадь водоема – 2530 га, средняя глубина – 5.4 м, максимальная – 22 м, дно водохранилища в основном илистое, местами с каменистым песком, или каменистое, водоем проточный. В зоне сбросных теплых каналов и прилегающей к ней акватории температура воды в период роста и развития личинок (июль–август) составляет 24–28°С, зимой в основной акватории 4–5°С. Вследствие высоких летних температур и активного перемешивания вода обогащена кислородом, органическими и минеральными веществами, что создает благоприятную среду для роста зоо- и фитопланктона [14]. Из Рефтинского водохранилища по р. Рефт лягушки попали в р. Пышму, где в 80-е годы XX в. сформировались самовоспроизводящиеся популяции. В р. Пышма температура воды ниже, чем в Рефтинском водохранилище, и соответствует температурному режиму естественных водоемов на Среднем Урале [14].
Для работы использовали P. ridibundus, пойманных в конце мая в водоеме-охладителе Рефтинской ГРЭС, и из одной популяции р. Пышма (расстояние 40 км от водохранилища; окрестности с. Курьи). В р. Пышма постоянные группировки отмечены в начале 1990-х годов. Выборка состояла из сеголеток, субадультусов (особей, переживших одну зимовку) и взрослых животных, пришедших на размножение. Земноводные были отобраны весной в период размножения. Всего исследовано 452 особи половозрелых P. ridibundus, из них 38 – из р. Пышмы (2017–2018 гг.), 414 – из Рефтинского водохранилища (1993–2015 гг.).
Исследованы наиболее значимые характеристики для бесхвостых амфибий: размерно-возрастной состав популяции, сроки размножения, возраст полового созревания, плодовитость. Данные по возрасту и плодовитости были получены при оценке только взрослых животных.
У каждой особи измеряли длину тела от конца морды до переднего края клоакальной щели – L (Longitudo corporis) с точностью до 0.1 мм и ампутировали четвертый палец задней конечности, из третьей фаланги которого изготовляли препараты для определения индивидуального возраста [15]. Для определении скорости резорбции костной ткани по методике Э.М. Смириной [16] были исследованы сеголетки из р. Пышмы (N = 3) и Рефтинского водохранилища (N = 27). Срезы толщиной 20–25 мкм делали в самом центре диафиза фаланги пальца, окрашивали гематоксилином Эрлиха и заключали в глицерин [16, 17]. Измерения проводили с помощью окуляр-микрометра – цена деления 0.01 мм. Размеры кости в поперечном сечении сопоставляли у сеголеток и лягушек, переживших одну зимовку, с величиной костно-мозговой полости и диаметром кости, ограниченной первой видимой целиком линией склеивания и последующими линиями у неполовозрелых и половозрелых особей. Для расчета числа полных прожитых лет P. ridibundus к видимым линиям склеивания прибавляли один год [17]. Сроки размножения определяли по появлению первых и последних кладок икры, а половозрелость самцов – по развитым наружным резонаторам. Для определения плодовитости самок вскрывали, извлекали яичники со зрелыми ооцитами и подсчитывали количество икринок с помощью лупы МБС-10.
Значимость различий между выборками оценивали с помощью дисперсионного однофакторного анализа. Различия считали значимыми на уровне p < 0.05. Статистический анализ проведен с помощью пакета прикладных программ “Statistica 6.0”, (StatSoft, 2001) (лиц. № AXXR003A622407FAN8).
Исследования на р. Пышма только начаты, но полученные результаты показали, что по ряду показателей животные из этой реки отличаются от животных из Рефтинского водохранилища. Размеры тела – одна из главных морфофизиологических характеристик, которая определяет степень адаптивности вида и популяции [18]. Наблюдающаяся уже на ранних стадиях наземного роста дифференцировка в размерах тела животных в дальнейшем может привести к тому, что более крупные особи становятся раньше половозрелыми и могут участвовать в размножении на год раньше [19, 20]. Установлено, что самки имеют на 8% более высокий темп роста, чем самцы [21].
Нами были получены данные, позволяющие провести сравнительный анализ размеров тела лягушек разного возраста (рис. 1). Видимые различия между длиной тела животных разного возраста в зависимости от местообитания для всех особей отсутствуют (F (5;43) = 0.70, p = 0.62), но есть тенденция к увеличению длины тела у лягушек из Рефтинского водохранилища. У лягушек, обитающих в Рефтинском водохранилище, после трех зимовок (F = (1;12) = 81.88, p < 0.01), как и после четырех (F = (1; 87) = 28.40, p < 0.01), длина тела достоверно больше, чем в р. Пышма. Так, средняя длина тела у трехлетних животных из Рефтинского водохранилища составляет 93.0 ± 1.3 мм против 68.1 ± 1.4 мм у лягушек из р. Пышма, после четырех зимовок соответственно 101.2 ± 1.3 и 72.5 ± ± 1.0 мм. Наблюдается перекрывание размеров одновозрастных животных во всех возрастных группах. Например, лягушки из р. Пышма могут иметь длину тела 71.4 мм после трех и четырех зимовок.
Рис. 1.
Длина тела особей P. ridibundus разного возраста в Рефтинском водохранилище (1) и р. Пышма (2).
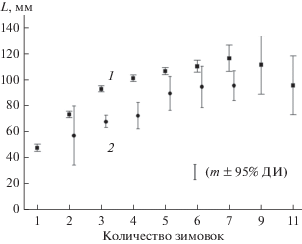
У P. ridibundus из Рефтинского водохранилища максимальное количество зимовок (11) зарегистрировано у единственной самки с длиной тела 95.8 мм. Известно, что самые старые особи не всегда самые крупные: например, самая крупная особь имела длину тела 132.2 мм после 6 зимовок [12]. В выборке из р. Пышмы максимальное количество зимовок (7) отмечено у самки, длина тела которой составила 107.0 мм. В естественном ареале, в Одесской области, обитают более крупные лягушки – со средней длиной тела 130.0–139.0 мм, максимально 170 мм [22].
С мест зимовок P. ridibundus чаще всего выходят в марте–апреле; в южных регионах, в теплые зимы, они могут быть активны круглый год. Спаривание начинается вскоре после зимовки; период икрометания продолжается более 3 мес. с возможностью повторного размножения у части особей [22–24].
На Среднем Урале P. ridibundus после зимовки обычно появляются в конце апреля–начале мая. В исследованных водоемах они приступают к размножению в середине мая при температуре воды 15–18°С, репродуктивный период занимает в среднем около трех недель. В 2018 г. в связи с холодной и затяжной весной размножение в р. Пышма проходило позже: первые кладки отмечены в начале июня. По нашим данным, личиночный период занимает от 70 до 90 сут [25]. Возможно, в связи с этим особи, завершившие метаморфоз в самом конце теплого сезона, не успевают подготовиться к зимовке.
На возраст полового созревания влияют условия существования [22, 23]. В Северной Греции P. ridibundus становятся половозрелыми уже после первой зимовки при длине тела 62.0–66.0 мм [24]. В горных водоемах Армении незначительная часть особей начинает размножаться после третьей зимовки, а основная масса – на год позже [26]. В Рефтинском водохранилище часть самок приступает к размножению после второй зимовки при длине тела от 78.0 до 83.0 мм (соответственно от 2523 до 2448 шт. икринок в яичниках), а после третьей зимовки все самки становятся половозрелыми [19].
В р. Пышма отловлено 22 самки, переживших три зимовки. Зрелые ооциты, готовые к выметыванию, были обнаружены в яичниках трех самок с длиной тела 61.1, 75.8 и 76.5 мм (соответственно 1260, 2417, 2990 икринок в яичниках). У самок с длиной тела 62.7–74.4 мм яичники имели бледно-песочный цвет, икринки были не видны невооруженным глазом – это указывает на то, что животные были неполовозрелыми. Таким образом, в р. Пышма самки приступают к размножению на год позже, чем в водохранилище: незначительная часть животных начинает размножаться после третьей зимовки, а основная масса – после четвертой.
Абсолютная плодовитость самок в Рефтинском водохранилище составляет 5778 ± 267 икринок, минимальная – 1199 икринок при длине тела 88.3 мм после двух зимовок, максимальная – 13 252 икринки у самки, которая пережила четыре зимовки и при длине тела 124.0 мм. Плодовитость лягушек в р. Пышма составляет 4456 ± 1754 икринки: максимальная (11 166 икринок) – у самки при длине тела 107.0 мм, пережившей семь зимовок, минимальная (1260 икринок) – у самки после трех зимовок с длиной тела 61.1 мм. По числу яиц самки в р. Пышма не отличаются от самок из Рефтинского водохранилища (F (1;39) = 0.01, p = = 0.96) и естественных популяций в Одесской области, где средняя абсолютная плодовитость P. ridibundus составляет 5400 (850–12 400) икринок [22].
Таким образом, P. ridibundus, обитающая в водоемах, различающихся по температурному режиму, обладает специфическим набором характеристик, прежде всего связанных с размерами и возрастом животных. В некоторых возрастных генерациях одновозрастные лягушки, обитающие в Рефтинском водохранилище, значимо крупнее, чем в р. Пышма.
В р. Пышма самки впервые приступают к размножению на год позже, чем в водохранилище: незначительная часть животных начинает размножаться после третьей зимовки, а основная масса – после четырех. По числу отложенных яиц лягушки, обитающие в исследуемых водоемах, также не отличаются.
После обнаружения P. ridibundus в водоемах-охладителях этот вид был включен в Красную книгу Среднего Урала 2006 г. [27]. Благодаря нашим исследованиям было показано, что данный вид является инвазивным, что позволило исключить P. ridibundus из числа охраняемых видов амфибий в новом издании Красной книги Свердловской области 2008 г. [28].
Полученные нами результаты подтверждают высокую экологическую пластичность вида, позволяющую ему осваивать новые водоемы и таким образом значительно расширять свой ареал.
Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН, а также частично поддержана Комплексной программой УрО РАН (проект № 18-4-4-28) и программой 211 Правительства Российской Федерации (соглашение № 02.A03.21.0006).
Список литературы
Решетников А.Н. Поедает ли ротан Perccottus glenii (Perciformes: Odontobutidae) икру рыб и амфибий // Вопр. ихтиологии. 2008. Т. 48. № 3. С. 384–392.
Пучковский С.В. Современное состояние биоразнообразия – продолжение эволюции // Успехи соврем. биол. 2016. Т. 136. № 5. С. 449–459.
Алимов А.Ф., Богуцкая Н.Г., Орлова М.И. Вид в пределах ареала и в области инвазии // Биологические инвазии в водных и наземных экосистемах. Под ред. Алимова А.Ф., Богуцкой Н.Г. М.; СПб.: КМК, 2004. С. 37–43.
Белимов Г.Т., Седалищев В.Т. Озерная лягушка Rana ridibunda Pall. в водоемах Якутска // Вестн. зоологии. 1980. № 3. С. 74–75.
Яковлев В.А. Расселение озерной лягушки на Алтае // Исчезающие и слабо изученные растения и животные Алтайского края и проблемы их охраны. Барнаул, 1987. С. 100–101.
Попов В.В. Новая встреча озерной лягушки Rana ridibunda Pallas, 1771 в окрестностях г. Зеленогорска (Красноярский край) // Байкальский зоол. журн. 2012 № 2 (10). С. 66–70.
Ляпков С.М. Озерная лягушка (Pelophylax ridibunda) в термальных водоемах Камчатки // Зоол. журн. 2014. Т. 93. № 12. С. 1427–1432.
Мисюра А.Н. Экология фонового вида амфибий центрального степного Приднестровья в условиях промышленного загрязнения водоемов: Автореф. дис. … канд. биол. наук. М., 1989. 16 с.
Фоминых А.С., Ляпков С.М. Формирование новых особенностей жизненного цикла озерной лягушки (Rana ridibunda) в условиях подогреваемого водоема // Журн. общ. биол. 2011. Т. 72. № 6. С. 403–421.
Большаков В.Н., Иванова Н.Л. Озерная лягушка (Pelophylax ridibundus Pall.) – чужеродный вид в водных экосистемах Среднего Урала // Озерные экосистемы: биологические процессы, антропогенная трансформация, качество воды: Тр.V Междунар. науч. конф. Минск, 2016. С. 12–17.
Топоркова Л.Я., Боголюбова Т.В., Хафизова Р.Т. К экологии озерной лягушки, интродуцированной в водоемы горно-таежной зоны Среднего Урала // Фауна Урала и Европейского Севера. Свердловск: Изд-во Урал. гос. ун-та, 1979. С. 108–115.
Иванова Н.Л. Особенности экологии озерной лягушки (Rana ridibunda Pall.), интродуцированной в водоемы-охладители // Экология. 1995. № 6. С. 473–476.
Вершинин В.Л., Иванова Н.Л. Специфика трофических связей вида-вселенца – Rana ridibunda Pallas, 1771 в зависимости от условий местообитаний // Поволж. экол. журн. 2006. № 3. С. 119–128.
Прохорова Н.Б., Черняев А.М., Баженова Г.А. и др. Гидротехническое регулирование водных ресурсов // Водные ресурсы Свердловской области. Екатеринбург: АМБ, 2004. С. 123–175.
Терентьев П.В., Чернов С.А. Определитель пресмыкающихся и земноводных. М.: Сов. наука, 1949. 340 с.
Смирина Э.М. Годовые слои в костях травяной лягушки (Rana temporaria) // Зоол. журн. 1972. Т. 51. Вып. 10. С. 1529–1534.
Иванова Н.Л. Характер и темпы роста озерной лягушки Pelophylax ridibundus Pall., интродуцированной в водоемы Среднего Урала // Изв. РАН. Сер. биол. 2017. № 4. С. 413–417.
Шварц С.С., Смирнов В.С., Добринский Л.Н. Метод морфофизиологических индикаторов в экологии наземных позвоночных животных. Свердловск: Урал. фил. АН СССР. 1968. 386 с.
Иванова Н.Л. Озерная лягушка (Rana ridibunda Pallas, 1771) в водоемах-охладителях на Среднем Урале // Экология. 2002. № 2. С. 137–141. [Ivanova N.L. Marsh frog (Rana redibunda Pall) in cooling ponds in the Middle Urals // Rus. J. of Ecology. 2002. V. 33. № 2. P. 125–128.]
Иванова Н.Л., Жигальский О.А. Демографические особенности популяций озерной лягушки (Rana ridibunda Pall.), интродуцированной в водоемы Среднего Урала // Экология. 2011. № 5. С. 361–369. [на англ. языке]
Иванова Н.Л., Кшнясев И.А. Особенности постметаморфического роста озерной лягушки в водоемах-охладителях Среднего Урала // Популяционная экология животных : Мат-лы II Междунар. науч. конф. Томск, 2016. С. 53.
Писанец Е.М. Земноводнi Украiни. Киiв: Изд-во Раевського, 2007. 192 с.
Жукова Т.И., Широкова В.Б. Анализ репродуктивного цикла озерной лягушки на Северном Кавказе по степени зрелости гонад // Герпетология. Краснодар: Кубан. ун-т, 1979. С. 13–18.
Kyriakopoulou-Sklavounou P., Loumbourdis N. Annual Ovarian Cycle in Frog, Rana ridibunda in Northern Greece // J. Herpetology. 1990. V. 24. № 2. P. 185–191.
Иванова Н.Л., Кашковская В.П. Особенности постметаморфического роста озерной лягушки, интродуцированной в водохранилища на Среднем Урале // Труды Ин-та зоол. нац. академии наук Азербайджана. 2017. Т. 35. № 1. С. 129–137.
Александровская Т.О., Котова Е.Л. Предварительные данные по возрастной характеристике озерной лягушки (Rana ridibunda Pall.) из трех точек Армении // Тр. Зоол. ин-та АН СССР. 1986. Т. 157. С. 177–180.
Ищенко В.Г., Юшков Р.А., Воронов Г.А. Озерная лягушка Rana ridibunda Pallas, 1771 // Красная книга Среднего Урала: Свердловская и Пермская области: редкие и находящиеся под угрозой исчезновения виды животных и растений. Екатеринбург: Изд-во Урал. ун-та, 1996. С. 50.
Красная книга Свердловской области: Животные, растения, грибы // Отв. ред. Корытин Н.С. Екатеринбург: Баско, 2008. 254 с.
Дополнительные материалы отсутствуют.