

Экология, 2019, № 6, стр. 429-437
Эмиссия диоксида углерода почвами как критерий эффективности ремедиации техногенных пустошей вблизи медно-никелевых комбинатов в Кольской Субарктике
М. С. Кадулин a, *, Г. Н. Копцик a, **
a Московский государственный университет им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
* E-mail: tubmaxxl@mail.ru
** E-mail: koptsikg@mail.ru
Поступила в редакцию 28.12.2018
После доработки 03.04.2019
Принята к публикации 08.04.2019
Аннотация
Техногенные пустоши, сформировавшиеся под воздействием выбросов диоксида серы и тяжелых металлов вблизи предприятий цветной металлургии на Кольском полуострове, характеризуются заторможенным выделением CO2 из почв (10–30 мг С м–2 ч–1, или 12–26 г С м–2 лето–1). Ремедиация пустошей способствует росту корней растений и микроорганизмов, дыхание которых ускоряет поток СО2 из почв вдвое при хемофитостабилизации и впятеро – при нанесении плодородного слоя; при этом доля корневого дыхания возрастает от 0 до 40–60%. Интенсивность эмиссии СО2 почвами и структура его продуцирования могут использоваться в качестве критериев эффективности ремедиации техногенных пустошей.
Деградация наземных экосистем, обусловленная хозяйственной деятельностью человека, является одной из причин изменения концентрации диоксида углерода (СО2) в атмосфере, рост которой может быть ответственен за трансформацию климата на нашей планете. При этом положительный баланс СО2 в атмосфере реализуется за счет снижения продуктивности фитоценозов на фоне непрерывной микробной минерализации запасов органического вещества почв и растений [1, 2]. Такие процессы активно протекают под влиянием выбросов крупных промышленных предприятий, включая горно-металлургические комбинаты (ГМК) “Печенганикель” и “Североникель” в Кольской Субарктике. Длительное атмосферное загрязнение диоксидом серы (SO2) и тяжелыми металлами (ТМ) в сочетании с суровым климатом привело к повреждению лесных фито- и микробоценозов и деградации почв вплоть до образования вблизи источников загрязнения техногенных пустошей [3–6].
Несмотря на сокращение промышленных выбросов, сохраняющиеся экстремальные уровни загрязнения почв и их токсичность для биоты делают длительным или даже невозможным в обозримом будущем восстановление фитоценозов без вмешательства человека [3, 5, 6]. Ремедиация создает условия для роста биомассы растений и микроорганизмов в почвах, что способствует изъятию CO2 из атмосферы и началу стока углерода в экосистемы. Увеличение количества корней и микробных клеток в почвах требует дополнительного расхода энергии на поддержание их биомассы через дыхание. В свою очередь дыхание почв рассматривается как интегральный показатель их биологической активности [7] и может использоваться как критерий восстановления нарушенных экосистем [8].
Выделение диоксида углерода почвами при восстановлении нарушенных экосистем практически не исследовано. В этой связи целью нашей работы было изучение влияния ремедиации на эмиссию диоксида углерода почвами техногенных пустошей, оценка участия корней растений и микроорганизмов, а также вклада разных слоев почв в формирование потока CO2 с их поверхности в зонах влияния ГМК “Печенганикель” и “Североникель” в Кольской Субарктике.
МАТЕРИАЛ И МЕТОДЫ
Объектами исследования являлись автоморфные подзолы иллювиально-железистые и абраземы альфегумусовые техногенных пустошей вблизи (1–5 км) ГМК “Печенганикель” (п. Никель и г. Заполярный) и “Североникель” (г. Мончегорск) до и после их ремедиации. Вблизи ГМК “Печенганикель” исследовали 6 участков пустошей и 8 участков ремедиации, вблизи ГМК “Североникель” – 4 и 8 участков соответственно. Ремедиация техногенных пустошей была проведена Печенгским и Мончегорским лесхозами по рекомендациям Института проблем промышленной экологии Севера КНЦ РАН при поддержке Кольской ГМК в 2003–2008 гг. с помощью двух технологий. Первая, хемофитостабилизация, включала высадку саженцев с одновременным посевом травяно-злаковой смеси и внесением азофоски и извести без предварительной обработки почвы. Вторая технология предусматривала предварительное нанесение естественного (пахотный горизонт почв заброшенных сельхозугодий) или искусственно созданного (на основе торфа, песка, опилок, осадка сточных вод) плодородного слоя поверх исходных загрязненных почв, внесение азофоски и извести. После этого проводили высадку саженцев ивы и березы и посев злаков.
Методы исследования. Измерения эмиссии СО2in situ проводили ежегодно по одному разу в месяц в июле 2010–2013 гг. (лето), а также в мае (весна) и сентябре (осень) 2012 г. (в общей сложности 5 раз) методом закрытых камер с помощью ИК СО2-газоанализатора AZ 7752 в 8–16-кратной повторности. Разделение корневого и микробного дыхания осуществляли с помощью полевого метода субстрат-индуцированного дыхания [9]. Летние потоки СО2 из почв (за календарное лето – с 1 июня по 31 августа) рассчитывали с помощью уравнения Аррениуса в модификации [10], чувствительность дыхания почв к изменению температуры (Q10) – по уравнению Вант-Гоффа. Подробное описание методов приведено в наших предыдущих работах [11].
Для оценки вертикальной структуры продуцирования СО2 рассчитывали отношение выделения СО2 из нижележащего горизонта к выделению с поверхности почвы. Для почв пустошей и участков хемофитостабилизации нижним горизонтом был BC, а при нанесении плодородного слоя – BF.
Полевой микробный метаболический коэффициент (qпCO2, [мг С-СО2микр м–2 ч–1]/[г Смикр/м–2]) оценивали по следующей оригинальной формуле:
Содержание Смикр определяли методом фумигации-экстракции [12], концентрацию углерода и азота в водных вытяжках (1 : 10) измеряли на Shimadzu TOC(N) analyzer, водорастворимых форм биофильных элементов (Ca, Mg, K и P) и ТМ (Ni и Cu) – с помощью масс-спектроскопии с индуктивно связанной плазмой, рН – потенциометрически.
Расчет средних значений и стандартных отклонений, корреляционный анализ, построение регрессионных моделей, а также проверку значимости различий по непараметрическому U-тесту Манна-Уитни проводили с помощью программы Statistica 8.0 (Statsoft, USA).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Интенсивность эмиссии СО2 (10–30 мг С м–2 ч–1) почвами техногенных пустошей (участки П) вблизи комбинатов крайне низка (рис. 1а) – на порядок ниже по сравнению с фоновыми лесами Кольской Субарктики (230–270 мг С м–2 ч–1). Выделение СО2 почвами березового редколесья (МБ1), сохранившегося на границе с пустошами вблизи г. Мончегорска в 5 км от комбината и проявляющего признаки естественного восстановления, также заторможено. Такие низкие скорости эмиссии СО2 почвами типичны для техногенных пустошей вблизи Карабашского медеплавильного завода в сосново-березовых лесах на Южном Урале (0–54 мг С м–2 ч–1) [7, 13], а также свойственны почвам аридных пустынь [14].
Рис. 1.
Эмиссия СО2 почвами техногенных пустошей до и после ремедиации в окрестностях п. Никель и г. Мончегорск. Здесь и на рис. 2: а – летние периоды 2010–2013 гг.; б – 2012 г.; в – лето 2012 г. Показаны средние и стандартные отклонения.
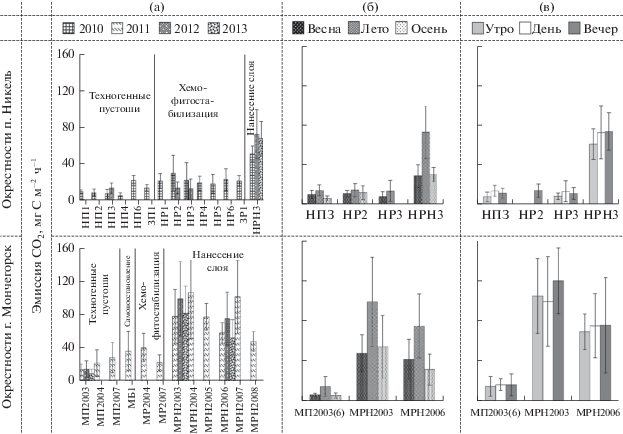
После хемофитостабилизации пустошей (участки Р) выделение СО2 возросло в среднем в 1.5–2 раза (до 20–40 мг С м–2 ч–1), однако она не всегда приводила к достоверной интенсификации выделения СО2 из почв [15, 16]. Наибольший эффект ремедиации в отношении биологической активности почв был достигнут за счет перекрытия загрязненных почв плодородным слоем (участки РН), благодаря чему выделение СО2 значимо (p < 0.007) увеличилось от – 1.5 до 8 раз (50–110 мг С м–2 ч–1). По скорости эмиссии СО2 почвы участков ремедиации с нанесением плодородного слоя приближаются к почвам лесных экосистем стадии дефолиации (85–200 мг С м–2 ч–1) [15, 16], но далеко не достигают фонового уровня. Близкая интенсивность выделения CO2 свойственна песчаным почвам после лесомелиорации карьеров по добыче фосфоритов под молодыми (20–25 лет) сосновыми лесами (20–200 мг С м–2 ч–1) [17] или почвам песчаных пустынь, покрытых древесной растительностью (саксаул зайсанский, 40–80 мг С м–2 ч–1) [14].
Сезонная динамика эмиссии СО2 почвами техногенных пустошей до (p < 0.03) и после их хемофитостабилизации (p ~ 0.05–0.2) и перекрытия плодородным слоем (p < 0.009) была типична для наземных экосистем: летом выделялось в 1.5–2.5 раза больше СО2, чем весной и осенью (рис. 1б). В этом смысле ремедиация почв не повлияла на сезонную динамику СО2. На протяжении суток летом заметного изменения скорости эмиссии СО2 не обнаружено (рис. 1в), по-видимому, из-за слабого изменения условий среды (температуры и освещенности) в течение полярного дня для протекания биохимических реакций и газообмена почвы с атмосферой.
Поток СО2 из почвы формируется за счет дыхания корней растений и почвенных микроорганизмов. Почвы техногенных пустошей практически лишены растительности, и поэтому уровень корневого дыхания в них ниже пределов чувствительности метода. После хемофитостабилизации условия развития корней растений существенно не улучшились; корневое дыхание было близко к нулю (рис. 2а). В свою очередь нанесение плодородного слоя привело к росту эмиссии СО2 за счет усиления как микробного, так и корневого дыхания, на долю которого приходилось от 30 до 80%. Столь высокий вклад корневого дыхания совпадает со значениями, характерными для почв фоновых лесов Кольской Субарктики [11].
Рис. 2.
Вклад корней в поток CO2 из почв техногенных пустошей до и после ремедиации в окрестностях п. Никель и г. Мончегорск.
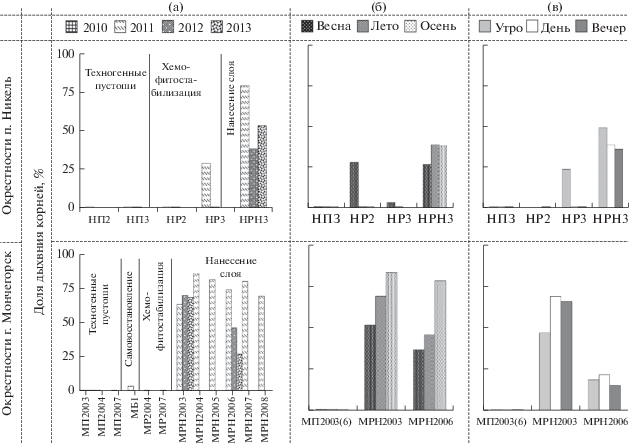
На протяжении бесснежного периода года в силу крайне низкого содержания корней в почвах сезонная и суточная динамика корневого дыхания на участках техногенных пустошей и хемофитостабилизации была не выражена. Максимальное развитие корневой системы растений произошло при нанесении плодородного слоя. При этом наименьший вклад дыхания корней в эмиссию СО2 был выявлен в начале вегетации растений – весной, тогда как максимальный (в 1.5–2 раза больше) – летом и осенью (рис. 2б). В течение суток динамика доли корневого дыхания, как и общей эмиссии СО2, была не выражена (рис. 2в).
Структура вертикального продуцирования СО2. В естественной почве основная масса живых организмов локализована ближе к поверхности, и поэтому продуцирование СО2 максимально в верхних почвенных горизонтах. При проведении ремедиации затрагивается только верхняя часть почвенной толщи. Для выявления прямого эффекта ремедиации на биологическую активность почв нами было рассчитано отношение выделения CO2 нижележащим слоем к общему потоку СО2 с поверхности почв.
Дыхание нижележащих слоев почв пустошей было сопоставимо или даже превосходило дыхание поверхностного слоя (рис. 3). Это, видимо, обусловлено неблагоприятными для живых организмов свойствами верхнего слоя почв пустошей [5], разгрузкой почвенной толщи от СО2 нижних слоев после механического нарушения при проведении измерений, ускорением окисления органического вещества при изменении окислительно-восстановительных условий [18], а также усилением дыхания тканей растений при их механическом повреждении.
Рис. 3.
Соотношение эмиссии СО2 нижними горизонтами и эмиссии с поверхности почв до и после ремедиации в окрестностях г. Мончегорска.
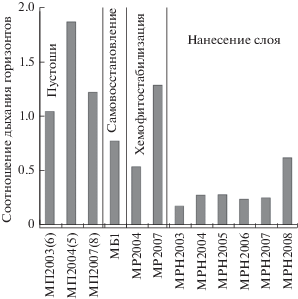
В результате хемофитостабилизации пустошей заметных изменений в структуре вертикального продуцирования СО2 не выявлено, а нанесение плодородного слоя в большинстве случаев сопровождалось снижением соотношения выделения СО2 нижележащей толщей и поверхностью почвы до значений (0.18–0.28), сопоставимых с фоновыми еловыми лесами Кольской Субарктики (0.22, [15]) и южной тайги на Валдае (0.30, [19]).
Эмиссия СО2 значительно меняется во времени и при нормальном уровне увлажнения определяется в первую очередь температурой, при которой протекают реакции дыхания внутри клеток живых организмов. Поэтому единичные измерения эмиссии СО2 дают лишь общее представление об интенсивности биохимических процессов в почвах. Корректное сравнение различных объектов возможно только при приведении результатов измерений к одинаковой температуре. Для этого нами были рассчитаны средние потоки CO2 за летние периоды 2010–2013 гг.
На техногенных пустошах до и после ремедиации была выявлена значимая зависимость интенсивности выделения CO2 из почв от температуры воздуха при использовании уравнения Вант-Гоффа и модели Аррениуса в модификации [10] (рис. 4). Небольшие величины коэффициентов детерминации (R2 = 0.22–0.40) могут свидетельствовать как о меньшем значении температуры воздуха для дыхания почв по сравнению с температурой почв и необходимости учета фактора влажности, так и об угнетении биологической активности почв под влиянием загрязнения, сглаживающем динамику выделения СО2. В то же время при исследования фоновых лесов Кольской Субарктики теснота связи эмиссии СО2 из почв и температуры была заметно выше (R2 = 0.2–0.8 [11]). Очень похожая закономерность также обнаружена при исследовании лесомелиорации карьеров по добыче фосфоритов в Московской области, где зависимость эмиссии СО2 из почв от температуры и влажности под посадками сосны (20–25 лет) была значительно слабее (R2 = 0.27) по сравнению с ненарушенными сообществами южной тайги (R2 = 0.51–0.70) [17].
Рис. 4.
Зависимость эмиссии СО2 из почв от температуры воздуха в 2010–2013 гг. Здесь и на рис. 5: НП и МП – техногенные пустоши, НР и МР – участки хемофитостабилизации, НРН и МРН – участки ремедиации с нанесением плодородного слоя в окрестностях п. Никель (а – Н) и г. Мончегорск (б – М).
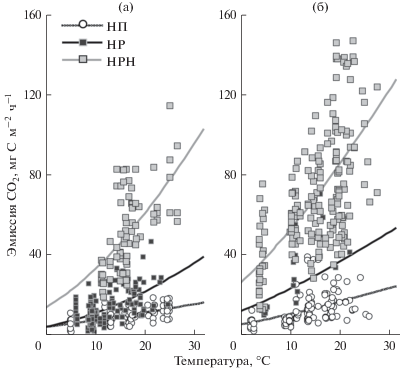
Рис. 5.
Летний поток СО2 из почв техногенных пустошей (1 – микроорганизмы, 2 – корни) до и после ремедиации в окрестностях п. Никель и г. Мончегорск; цифрами указана доля дыхания корней (%).
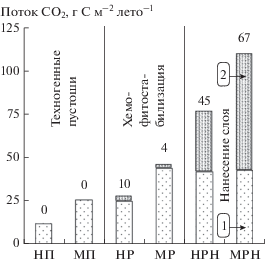
Летний поток СО2 из почв техногенных пустошей (НП, МП) был очень мал (12–26 г С м–2 лето–1) и, видимо, формировался за счет микробной деструкции старых запасов органического вещества. Поток СО2 вырос вдвое (28–46 г С м–2 лето–1) после хемофитостабилизации (НР, МР) и достиг максимума после нанесения плодородного слоя (НРН, МРН, 77–110 г С м–2 лето–1, рис. 5). Необходимо отметить, что и в первом, и во втором случаях активизация микробного дыхания была очень близкой (25–44 и 42–43 г м–2 лето–1). Однако при нанесении плодородного слоя увеличение общего потока СО2 произошло также благодаря интенсивному развитию корней растений в почвах (35–67 г С м–2 лето–1, или 45–61%), тогда как хемофитостабилизация на корневой поток почти не повлияла (1–3 г С м–2 лето–1, или 4–10%).
Динамика потока СО2 из почвы в атмосферу напрямую зависит от чувствительности почвы к изменению температуры окружающей среды, которая в свою очередь определяется энергией активации (Ea) реакций окисления органического вещества почвенными организмами и физических процессов газообмена почвы с атмосферой. В почвоведении как аналог Ea обычно рассматривают температуру активации (Ta = Ea/R, где R – универсальная газовая постоянная). Для количественной оценки температурной чувствительности дыхания почвы используют также коэффициент Вант-Гоффа (Q10), который показывает, во сколько раз возрастает скорость реакции при увеличении температуры на 10°С.
Почвы техногенных пустошей крайне неблагоприятны для развития в них жизни из-за загрязнения ТМ и истощения запасов органического вещества. При дефиците ферментов в почвах скорости биохимических реакций снижаются, вследствие чего замедляется отклик дыхания почв на изменение температуры среды [20]. Так как ТМ деактивируют ферменты, то вполне закономерно ожидать ослабление температурной чувствительности дыхания (Q10) загрязненных почв. В то же время при обеднении почв лабильным углеродом активность микроорганизмов будет зависеть в первую очередь от наличия источника энергии и только во вторую – от температуры [21]. Поэтому на техногенных пустошах Q10 дыхания почв составляет всего ~1.5 при норме в фоновых лесах в вегетационный период ~2.0 [11, 22]. Благодаря хемофитостабилизации (Q10 = 1.6–2.0) и нанесению плодородного слоя (Q10 = 1.6–1.7) условия существования организмов в почвах улучшаются, что способствует усилению влияния температуры на протекание биохимических реакций.
Кроме того, Q10 дыхания корней после хемофитостабилизации (1.7–2.7) и нанесения плодородного слоя (1.6–1.8) превышал Q10 общего дыхания почв. Существенное превышение Q10 дыхания корней по сравнению с почвой также отмечено в более ранних работах [22, 23]. Вероятно, что при одинаковой температуре среды различия Q10 дыхания корней и дыхания почвы являются следствием разных источников субстрата, используемого организмами. Энергетическое питание корней и микроорганизмов ризосферы зависит от поступления сахаров, образующихся при фотосинтезе, интенсивность которого значительно изменяется даже в течение суток, что обусловливает высокие значения Q10 корней. Дыхание же микроорганизмов-сапротрофов, населяющих основной объем почвенной массы, определяется общим пулом органического вещества, который относительно стабилен во времени, и поэтому Q10 дыхания почвы в целом невелико.
Основными факторами, мешающими развитию живых организмов в почвах пустошей вблизи комбинатов “Печенганикель” и “Североникель”, являются загрязнение ТМ $({\text{N}}{{{\text{i}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ – 7.0–10.2 мг/кг, ${\text{C}}{{{\text{u}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ – 8.6–18.5 мг/кг) и крайняя бедность такими элементами питания, как фосфор (практически отсутствует), азот (3–13 мг/кг) и калий (~4 мг/кг) (табл. 1). Низкие значения ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$ (4.1–4.2), поддерживаемые поступлением с атмосферными выпадениями серы (R = –0.54), обусловливают высокую подвижность ТМ.
Таблица 1.
Характеристика почв техногенных пустошей и участков ремедиации вблизи п. Никель и г. Мончегорск (лето 2012 г.)
| Показатели | Окрестности п. Никель | Окрестности г. Мончегорска | ||||
|---|---|---|---|---|---|---|
| НП | НР | НРН | МП | МР | МРН | |
| Корни* | 0.34 ± 0.25ae | 0.46 ± 0.37a | 1.75 ± 0.62b | 0.01 ± 0.02c | 0.05 ± 0.05d | 0.18 ± 0.17e |
| qпCO2** | 1.03 | 0.78 | 0.98 | 1.59 | 4.05 | 0.76 |
| Cмикр, мг/кг | 46 ± 28a | 109 ± 40b | 183 ± 64c | 66 ± 42a | 83 ± 34a | 355 ± 86d |
| ${{{\text{C}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$, мг/кг | 38 ± 1a | 21 ± 3b | 110 ± 16c | 17 ± 13b | 44 ± 9a | 72 ± 52a |
| ${{{\text{N}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$, мг/кг | 13 ± 2a | 5 ± 3b | 18 ± 13a | 3 ± 1c | 7 ± 1b | 19 ± 15a |
| C : ${{{\text{N}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$ | 3.0 ± 0.6a | 8.2 ± 6.4b | 8.3 ± 6.7b | 4.9 ± 1.5cd | 6.8 ± 2.2bc | 3.9 ± 1.1d |
| Cобщ, % | 3.1 ± 0.7a | 2.6 ± 0.8a | 4.2 ± 0.5b | 2.6 | 2.9 | 4.1 |
| Nобщ, % | 0.12 ± 0.08a | 0.11 ± 0.03a | 0.18 ± 0.02b | 0.11 | 0.13 | 0.20 |
| Sобщ, % | 0.08 ± 0.03a | 0.09 ± 0.03a | 0.06 ± 00a | 0.10 | 0.07 | 0.07 |
| ${{{\text{S}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$, мг/кг | 35 ± 6a | 46 ± 41a | 13 ± 3b | 42 ± 10a | 42 ± 23a | 21 ± 12c |
| ${{{\text{P}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$, мг/кг | 0 ± 0a | 0 ± 0a | 1 ± 1b | 0 ± 0a | 0 ± 0a | 4 ± 3c |
| ${{{\text{K}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$, мг/кг | 4 ± 3a | 7 ± 6ab | 13 ± 7b | 4 ± 1a | 7 ± 4ab | 26 ± 4c |
| ${\text{C}}{{{\text{a}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$, мг/кг | 18 ± 15a | 160 ± 170b | 67 ± 40c | 19 ± 17a | 92 ± 46c | 88 ± 63c |
| ${\text{N}}{{{\text{i}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$, мг/кг | 10.2 ± 4.3a | 8.5 ± 5.3ac | 1.5 ± 0.7b | 7.0 ± 3.0a | 7.1 ± 3.7a | 2.8 ± 2.1b |
| ${\text{C}}{{{\text{u}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$, мг/кг | 8.6 ± 2.6a | 1.6 ± 1.0b | 0.4 ± 0.2c | 18.5 ± 5.0d | 6.2 ± 3.4a | 1.7 ± 0.8b |
| Влажность, % | 7 ± 2a | 14 ± 9b | 38 ± 15c | 16 ± 3b | 15 ± 2b | 55 ± 43c |
Хемофитостабилизация техногенных пустошей способствовала уменьшению токсичности почв благодаря известкованию, приводящему к снижению кислотности (${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$ 4.7–5.5) и подвижности ${\text{N}}{{{\text{i}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$ (7.1–8.5 мг/кг) и ${\text{C}}{{{\text{u}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$ (1.6–6.2 мг/кг). Однако реализация этого метода не привела к заметному улучшению условий питания и развития растений. При перекрытии загрязненных и истощенных почв пустошей плодородным слоем произошло существенное снижение токсичности среды обитания организмов за счет ее пространственной изоляции от техногенно загрязненного субстрата и благоприятных химических и физических свойств вновь сформированного слоя. Этот слой характеризуется слабокислой реакций среды (${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$ 5.7–6.1), относительно высоким содержанием влаги (38–55%) и элементов питания (18–19 мг ${{{\text{N}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$/кг, 1–4 мг ${{{\text{P}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$/кг и 13–26 мг ${{{\text{K}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$/кг). Улучшение свойств почв способствовало высокой приживаемости саженцев деревьев и развитию трав. Несмотря на разную эффективность, ремедиация пустошей обеспечивает запуск круговорота углерода, усиление которого приводит к росту биологической активности почв в целом и их дыхания в частности за счет развития продуцентов CO2 в почве – корней растений и почвенных микроорганизмов. Микробное и корневое дыхание закономерно возрастают при увеличении биомассы микроорганизмов (R = 0.41–0.63) и тонких корней растений (R = 0.58–0.72) в почвах.
Микробная биомасса в почвах исходных пустошей очень низкая (45–65 мг Смикр/кг). Ремедиация содействует заметному увеличению микробной биомассы в почвах пустошей даже за счет их хемофитостабилизации (45–110 мг Смикр/кг) и значимому увеличению при нанесении плодородного слоя (185–355 мг Смикр/кг). Росту микробной биомассы (R = 0.43) способствует обогащение почв легкодоступным углеродом (${{{\text{C}}}_{{{{{\text{H}}}_{2}}{\text{O}}}}}$ – от 17–38 до 72–108 мг/кг) – источником энергии микроорганизмов, образующимся при разложении органического вещества, находящегося в составе нанесенного слоя, а также растительных остатков, начавших поступать в почву после ремедиации.
Количественным показателем состояния и устойчивости микробного сообщества почвы может служить микробный метаболический коэффициент [24]. В почвах техногенных пустошей существование микроорганизмов происходит в условиях стресса. Борьба за выживание требует дополнительных энергетических расходов на репарацию, детоксикацию и вывод токсикантов из клетки. В этой связи стоимость поддержания единицы микробной биомассы – метаболический коэффициент (qCO2), при загрязнении тяжелыми металлами может возрастать [25]. В результате ремедиации техногенное воздействие на микробные клетки ослабляется, о чем свидетельствует уменьшение qпCO2 после нанесения плодородного слоя (0.76–0.98) по сравнению с исходными пустошами (1.03–1.59). Одновременно также быстро возрастает биомасса корней (0.18–1.75 и 0.01–0.34 г/дм3 соответственно). Изменения qCO2 под действием хемофитостабилизации неоднозначны (0.78–4.05), а биомасса корней в почвах (0.05–0.46 г/см3) значимо не отличается от пустоши.
Поступление элементов питания (N, P, K) с удобрениями благоприятствует развитию корней (R = 0.37–0.67, 0.48–0.51, 0.38–0.71) и микроорганизмов (R = 0.45–0.77, 0.51–0.69, 0.35–0.67 соответственно) в почвах. Растения и микроорганизмы являются частью одних и тех же циклов биофильных элементов, поэтому их раздельное существование невозможно. Улучшение состояния растений создает условия для развития сапротрофных микроорганизмов за счет поступления органического вещества в почвы. В то же время за счет симбиотических микроорганизмов происходит улучшение минерального питания растений. Поэтому микробная и корневая биомасса закономерно тесно связаны (R = 0.69).
ВЫВОДЫ
1. Техногенные пустоши отличаются экстремально низким выделением СО2 (10–30 мг С м–2 ч–1, или 12–26 г С м–2 лето–1). Отклик биологической активности почв на ремедиацию пустошей зависит от используемой технологии. Хемофитостабилизация способствует слабой активизации эмиссии СО2 (20–40 мг С м–2 ч–1, или 28–46 г С м–2 лето–1) и практически не влияет на дыхание корней (4–10% общего дыхания). Ремедиация пустошей с нанесением плодородного слоя более эффективна и сопровождается значимым ростом не только общего потока СО2 (50–110 мг С м–2 ч–1, или 77–110 г С м–2 лето–1), но и доли корневого дыхания (45–61%).
2. Активное развитие живых организмов в верхнем слое почв после ремедиации техногенных пустошей способствует снижению вклада нижележащих минеральных горизонтов в эмиссию CO2 с поверхности почв после нанесения плодородного слоя (18–28%) по сравнению с хемофитостабилизацией и исходными пустошами (100%).
3. Ремедиация техногенных пустошей ослабляет воздействие стрессовых факторов на микробное сообщество почв, что сопровождается увеличением чувствительности дыхания почв к изменению температуры (Q10 от 1.5 до 1.6–2.0). При перекрытии загрязненных почв плодородным слоем проявляется тенденция к снижению полевого микробного метаболического коэффициента qпCO2 (от 1.03–1.59 до 0.76–0.98).
4. Согласно результатам корреляционного анализа, интенсификация выделения СО2 почвами после ремедиации прямо связана с массой корней, содержанием углерода микробной биомассы, водорастворимых соединений углерода и элементов минерального питания, рН и влажностью и обратно – с содержанием водорастворимых форм никеля и меди.
5. Дыхание почв, участие корней и минеральной толщи в формировании потока СО2 из почв могут использоваться в качестве критериев эффективности восстановления техногенно нарушенных экосистем.
Работа выполнена при финансовой поддержке РФФИ (проект № 18-04-01028).
Список литературы
Пулы и потоки углерода в наземных экосистемах России / Отв. ред. Заварзин Г.А. М.: Наука, 2007. 315 с.
Schlesinger W.H., Andrews J.A. Soil respiration and the global carbon cycle // Biogeochemistry. 2000. V. 48. № 1. P. 7–20. https://doi.org/10.1023/A:1006247623877
Евдокимова Г.А., Калабин Г.В., Мозгова Н.П. Содержание и токсичность тяжелых металлов в почвах зоны воздействия воздушных выбросов комбината “Североникель” // Почвоведение. 2011. № 2. С. 261–268. [Evdokimova G.A., Kalabin G.V., Mozgova N.P. Contents and toxicity of heavy metals in soils of the zone affected by aerial emissions from the Severonikel Enterprise // Eurasian Soil Sci. 2011. V. 44. № 2. P. 237–244. http://doi.org/10.1134/S1064229311020037]
Кашулина Г.М. Экстремальное загрязнение почв выбросами медно-никелевого предприятия на Кольском полуострове // Почвоведение. 2017. № 7. С. 860–873. [Kashulina G.M. Extreme pollution of soils by emissions of the copper-nickel industrial complex in the Kola Peninsula // Eurasian Soil Sci. 2017. V. 50. № 7. P. 837–849. http://doi.org/ 10.1134/S1064229317070031]https://doi.org/10.7868/S0032180X17070036
Копцик Г.Н., Копцик С.В., Смирнова И.Е. Альтернативные технологии ремедиации техногенных пустошей в Кольской Субарктике // Почвоведение. 2016. № 11. С. 1375–1391. [Koptsik G.N., Koptsik S.V., Smirnova I.E. Alternative technologies for remediation of technogenic barrens in the Kola Subarctic // Eurasian Soil Sci. 2016. V. 49. № 11. P. 1294–1309. http://doi.org/ 10.1134/S1064229316090088]https://doi.org/10.7868/S0032180X16090082
Лянгузова И.В. Динамические тренды содержания тяжелых металлов в растениях и почвах при разном режиме аэротехногенной нагрузки // Экология. 2017. № 4. С. 250–260. [Lyanguzova I.V. Dynamic trends of heavy metal contents in plants and soil under different industrial air pollution regimes // Rus. J. of Ecology. 2017. V. 48. № 4. P. 311–320. http://doi.org/ 10.1134/S1067413617040117]https://doi.org/10.7868/S0367059717040114
Сморкалов И.А., Воробейчик Е.Л. Почвенное дыхание лесных экосистем в градиентах загрязнения среды выбросами медеплавильных заводов // Экология. 2011. № 6. С. 429–435. [Smorkalov I.A., Vorobeichik E.L. Soil respiration of forest ecosystems in gradients of environmental pollution by emissions from copper smelters // Rus. J. of Ecology. 2011. V. 42. № 6. P. 311–320. http://doi.org/10.1134/S1067413611060166]
Bujalský L., Kaneda S., Dvorščík P., Frouz J. In situ soil respiration at reclaimed and unreclaimed post–mining sites: Responses to temperature and reclamation treatment // Ecological Engineering. 2014. V. 68. P. 53–59. http://doi.org /10.1016 /j.ecoleng.2014.03.048
Евдокимов И.В., Ларионова А.А., Шмитт М. и др. Экспериментальная оценка вклада дыхания корней растений в эмиссию углекислого газа из почвы // Почвоведение. 2010. № 12. С. 1479–1488. [Yevdokimov I.V., Larionova A.A., Schmitt M. et al. Experimental assessment of the contribution of plant root respiration to the emission of carbon dioxide from the soil // Eurasian Soil Sci. 2010. V. 43. № 12. P. 1373–1381. http://doi.org/10.1134 /S1064229310120070]
Lloyd J., Taylor J.A. On the temperature dependence of soil respiration // Functional Ecology. 1994. V. 8. P. 315–323. https://doi.org/10.2307/2389824
Кадулин М.C., Смирнова И.Е., Копцик Г.Н. Эмиссия диоксида углерода почвами лесных экосистем заповедника “Пасвик” в Кольской Субарктике // Почвоведение. 2017. № 9. С. 1098–1112. http://doi.org/ 10.7868/S0032180X17090039 [Kadulin M.S., Smir-nova I.E., Koptsik G.N. The emission of carbon dioxide from soils of the Pasvik nature reserve in the Kola Subarctic // Eurasian Soil Sci. 2017. V. 50. № 9. P. 1055–1068. http://doi.org/10.1134/S1064229317090034]
Vance E.D., Brookes P.C., Jenkinson D.S. An extraction method for measuring soil microbial biomass // Soil. Biol. Biochem. 1987. V. 19. P. 703–707. https://doi.org/10.1016/0038-0717(87)90052-6
Сморкалов И.А., Воробейчик Е.Л. Механизм стабильности эмиссии СО2 из лесной подстилки в условиях промышленного загрязнения // Лесоведение. 2016. № 1. С. 34–43.
Zhao C.Y., Zhao Z.M., Hong Z. et al. Contribution of root and rhizosphere respiration of Haloxylon ammodendron to seasonal variation of soil respiration in the Central Asian desert // Quaternary International. 2011. V. 244. P. 304–309. https://doi.org/10.1016/j.quaint.2010.11.004
Кадулин М.С., Копцик Г.Н. Эмиссия CO2 почвами в зоне влияния ГМК “Североникель” в Кольской Субарктике // Почвоведение. 2013. № 11. С. 1387–1396. [Kadulin M.S., Koptsik G.N. Emission of CO2 by soils in the impact zone of the Severonikel smelter in the Kola Subarctic region // Eurasian Soil Sci. 2013. V. 46. № 11. P. 1107–1116. http://doi.org/10.1134/ S1064229313110045].https://doi.org/10.7868/S0032180X13110063
Копцик Г.Н., Кадулин М.С., Захарова А.И. Влияние техногенного загрязнения на эмиссию диоксида углерода почвами в Кольской Субарктике // Журн. общ. биол. 2015. Т. 76. № 1. С. 48–62.
Почикалов А.В., Ларин Я.А., Арешин А.В., Карелин Д.В. Компоненты бюджета углерода в лесных посадках при рекультивации открытых горных выработок // Лесоведение. 2015. № 6. С. 447–457.
Kalbitz K., Marschner B. Controls of bioavailability and biodegradability of dissolved organic matter in soils // Geoderma. 2003. V. 113. P. 211–235. https://doi.org/10.1016/S0016-7061(02)00362-2
Карелин Д.В., Замолодчиков Д.Г., Каганов В.В. и др. Микробная и корневая составляющие дыхания дерново-подзолистых почв южной тайги // Лесоведение. 2017. № 3. С. 183–195.
Davidson E.A., Janssens I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change // Nature. 2006. V. 440. P. 165–173. https://doi.org/10.1038/nature04514
Razavi B.S., Blagodatskaya E., Kuzyakov Y. Nonlinear temperature sensitivity of enzyme kinetics explains canceling effect – a case study on loamy haplic Luvisol // Frontiers in Microbiology. 2015. V. 6 (1126). P. 1–13. https://doi.org/10.3389/fmicb.2015.01126
Курганова И.Н., Розанова Л.Н., Мякшина Т.Н. и др. Многолетний мониторинг эмиссии СО2 из дерново-подзолистой почвы: анализ влияния гидротермических условий и землепользования // Проблемы экологического мониторинга и моделирования экосистем. 2007б. Т. 21. С. 23–43.
Boone R.D., Nadelhoffer K.J., Canary J.D., Kaye J.P. Roots exert a strong influence on the temperature sensitivity of soil respiration // Nature. 1998. V. 396. P. 570–572. https://doi.org/10.1038/25119
Ананьева Н.Д. Микробиологические аспекты самоочищения и устойчивости почв. М: Наука, 2003. 223 с.
Brookes P.C. The use of microbial parameters in monitoring soil pollution by heavy metals // Biol. Fertil. Soils. 1995. V. 19. P. 269–279.
Дополнительные материалы отсутствуют.